- Opsin
-
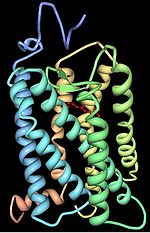 Three-dimensional structure of bovine rhodopsin. The seven transmembrane domains are shown in varying colors. The chromophore is shown in red.
Three-dimensional structure of bovine rhodopsin. The seven transmembrane domains are shown in varying colors. The chromophore is shown in red.
Opsins are a group of light-sensitive 35–55 kDa membrane-bound G protein-coupled receptors of the retinylidene protein family found in photoreceptor cells of the retina. Five classical groups of opsins are involved in vision, mediating the conversion of a photon of light into an electrochemical signal, the first step in the visual transduction cascade. Another opsin found in the mammalian retina, melanopsin, is involved in circadian rhythms and pupillary reflex but not in image-forming.
Contents
Opsin classification
There are two groups of protein termed opsins; these groups are not homologous, but have convergently evolved a similar form and function.[1][2] Type I opsins are employed by prokaryotes, whereas animals use type II opsins. No opsins have been found outside these groups (for instance in plants, fungi, or placozoans),[1] although as-yet unconfirmed reports of opsins in sponges[3] would suggest that opsins were present in the ancestral metazoan.
Based on the phylogeny of their sequences, type 2 opsins can be grouped into six families; these families are very distinct, with under 20% of their sequences shared with any other subfamily. The families consist of the vertebrate opsins/encephalopsins; Go opsins; Gs opsins; invertebrate Gq opsins; the photoisomerases and neuropsins.[4] These subfamilies can be grouped according to their expression; the first three are found in ciliary-type photoreceptor cells; Gq opsins in rhabdomeric-type photoreceptor cells; and the latter two are found elsewhere but based on their shared intron positions can be bundled together into the photoisomerases.[4]
Prokaryotic (type 1) opsins
Like eukaryotic opsins, prokaryotic opsins have a seven transmembrane domain structure similar to that found in eukaryotic G-protein coupled receptors. Despite this similarity, there is no evidence that they are evolutionarily related, suggesting that they evolved independently of one another.[2]
Several type 1 opsins, such as proteo-, halo- and bacteriorhodopsin, are used by various bacterial groups to harvest energy from light to carry out metabolic processes using a non-chlorophyll-based pathway. Additionally, sensory rhodopsins exist in Halobacteria that induce a phototactic response by interacting with transducer membrane-embedded proteins that have no relation to G proteins.[5]
Animal (type 2) opsins
Ciliary opsins
Ciliary opsins are expressed in ciliary photoreceptor cells, and include the vertebrate opsins/encephalopsins, Go and Gs opsin subfamilies.[4] They convert light signals to nerve impulses via cyclic nucleotide gated ion channels, which work by increasing the charge differential across the cell membrane (i.e. hyperpolarization.[1])
Vertebrate opsins
Vertebrate opsins can be further subdivided into rod opsins and four types of cone opsin, based on differential spatial expression, spectral sensitivity, and evolutionary history.[4] Rod opsins (rhodopsins, usually denoted Rh), are used in night vision, are thermally stable, and are found in the rod photoreceptor cells. Cone opsins, employed in color vision, are less-stable opsins located in the cone photoreceptor cells. Cone opsins are further subdivided according to their absorption maxima (λmax), the wavelength at which the highest light absorption is observed. Evolutionary relationships, deduced using the amino acid sequence of the opsins, are also frequently used to categorize cone opsins into their respective group. Both methods predict four general cone opsin groups in addition to rhodopsin.[6]
Humans have the following set of photoreceptor proteins responsible for vision:
- Rhodopsin (Rh1, OPN2, RHO) – expressed in rod cells, used in night vision
- Three cone opsins (also known as photopsins) – expressed in cone cells, used in color vision
Encephalopsins
This type of opsin is expressed throughout the mammalian body. It is also expressed in ciliary photoreceptor cells in annelids, and in the brains of some insects.[4]
Go / Gs opsins
These opsins, absent from higher vertebrates and arthropods, are found in the ciliary photoreceptor cells of molluscs and basal chordates (amphioxus); and in cnidarians, respectively.[4]
Rhabdomeric opsins
Arthropods and molluscs use Gq opsins. Arthropods appear to attain colour vision in a similar fashion to the vertebrates, by the use of three (or more) distinct groups of opsin, distinct both in terms of phylogeny and spectral sensitivity.[4] The Gq opsin melanopsin is also expressed in vertebrates, where it is responsible for the maintenance of circadian rhythms.[4]
Unlike ciliary opsins, these are associated with canonical transient receptor potential ion channels; these lead to the electric potential difference across a cell membrane being eradicated (i.e. depolarization).[1]
The identification of the crystal structure of squid rhodopsin[7] is likely to further our understanding of its function in this group.
Arthropods do use different opsins in their different eye types, but at least in Limulus the opsins expressed in lateral and in compound eyes are 99% identical and presumably diverged recently.[8]
Photoisomerases
This class of opsins are not coupled to a G-protein, and thus serve to traffic retinal around in response to light, rather than directly in signal-induction.[4]
Neuropsins
These opsins are found in nervous tissue in mammals, and despite some genetic similarities to photoisomerases, their function has not yet been identified.[4]
Name Gene Notes Melanopsin OPN4 best studied novel opsin involved in circadian rhythms and pupillary reflex Pineal Opsin (Pinopsin)[9] wide range of expression in the brain, most notably in the pineal region Vertebrate Ancient (VA) opsin[10] has three isoforms VA short (VAS), VA medium (VAM), and VA long (VAL). It is expressed in the inner retina, within the horizontal and amacrine cells, as well as the pineal organ and habenular region of the brain Parapinopsin (PP) Opsin [11] Extraretinal (or extra-ocular) Rhodopsin-Like Opsins (Exo-Rh)[12] Rhodopsin-like protein expressed in the pineal region Encephalopsin or Panopsin OPN3 originally found in human and mice tissue with a very wide range of expression (brain, testes, heart, liver, kidney, skeletal muscle, lung, pancreas and retina) Teleost Multiple Tissue (TMT) Opsin[13] Teleost fish opsin with a wide range of expression Peropsin or "Retinal pigment epithelium-derived rhodopsin homolog" RRH expressed in the retinal pigment epithelium (RPE) cells Retinal G protein coupled receptor RGR expressed in the retinal pigment epithelium (RPE) and Müller cells Neuropsin OPN5 Structure and function
Opsin proteins covalently bind to a vitamin A-based retinaldehyde chromophore through a Schiff base linkage to a lysine residue in the seventh transmembrane alpha helix. In vertebrates, the chromophore is either 11-cis-retinal (A1) or 11-cis-3,4-didehydroretinal (A2) and is found in the retinal binding pocket of the opsin. The absorption of a photon of light results in the photoisomerisation of the chromophore from the 11-cis to an all-trans conformation. The photoisomerization induces a conformational change in the opsin protein, causing the activation of the phototransduction cascade. The opsin remains insensitive to light in the trans form. It is regenerated by the replacement of the all-trans retinal by a newly synthesized 11-cis-retinal provided from the retinal epithelial cells. Opsins are functional while bound to either chromophore, with A2-bound opsin λmax being at a longer wavelength than A1-bound opsin.
Opsins contain seven transmembrane α-helical domains connected by three extra-cellular and three cytoplasmic loops. Many amino acid residues, termed functionally conserved residues, are highly conserved between all opsin groups, indicative of important functional roles. All residue positions discussed henceforth are relative to the 348 amino acid bovine rhodopsin crystallized by Palczewski et al..[14] Lys296 is conserved in all known opsins and serves as the site for the Schiff base linkage with the chromophore. Cys138 and Cys110 form a highly conserved disulfide bridge. Glu113 serves as the counterion, stabilizing the protonation of the Schiff linkage between Lys296 and the chromophore. The Glu134-Arg135-Tyr136 is another highly conserved motif, involved in the propagation of the transduction signal once a photon has been absorbed.
Certain amino acid residues, termed spectral tuning sites, have a strong effect on λmax values. Using site-directed mutagenesis, it is possible to selectively mutate these residues and investigate the resulting changes in light absorption properties of the opsin. It is important to differentiate spectral tuning sites, residues that affect the wavelength at which the opsin absorbs light, from functionally conserved sites, residues important for the proper functioning of the opsin. They are not mutually exclusive, but, for practical reasons, it is easier to investigate spectral tuning sites that do not affect opsin functionality. For a comprehensive review of spectral tuning sites see Yokoyama[15] and Deeb.[16] The impact of spectral tuning sites on λmax differs between different opsin groups and between opsin groups of different species.
External links
- Review of opsins and current research: Shichida, Y.; Matsuyama, T. (2009). "Evolution of opsins and phototransduction". Philosophical transactions of the Royal Society of London. Series B, Biological sciences 364 (1531): 2881–2895. doi:10.1098/rstb.2009.0051. PMC 2781858. PMID 19720651. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2781858.
- Illustration at Baldwin-Wallace College
- MeSH Opsin
References
- ^ a b c d Plachetzki, D.; Fong, C.; Oakley, T. (2010). "The evolution of phototransduction from an ancestral cyclic nucleotide gated pathway". Proceedings. Biological sciences / the Royal Society 277 (1690): 1963–1969. doi:10.1098/rspb.2009.1797. PMC 2880087. PMID 20219739. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2880087.
- ^ a b Fernald, R. D. (2006). "Casting a genetic light on the evolution of eyes". Science 313 (5795): 1914–1918. Bibcode 2006Sci...313.1914F. doi:10.1126/science.1127889. PMID 17008522.
- ^ "Porifera: Amphimedon queenslandica (sponge) .. 1 opsin". GenomeWiki. University of California Santa Cruz (UCSC) Genome Bioinformatics Group. http://genomewiki.ucsc.edu/index.php?title=Opsin_evolution:_key_critters_(cnidaria)&oldid=10719#Porifera:_Amphimedon_queenslandica_.28sponge.29_.._1_opsin.
- ^ a b c d e f g h i j Shichida, Y.; Matsuyama, T. (2009). "Evolution of opsins and phototransduction". Philosophical transactions of the Royal Society of London. Series B, Biological sciences 364 (1531): 2881–2895. doi:10.1098/rstb.2009.0051. PMC 2781858. PMID 19720651. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2781858.
- ^ Römpler, H.; Stäubert, C.; Thor, D.; Schulz, A.; Hofreiter, M.; Schöneberg, T. (2007). "G protein-coupled time travel: evolutionary aspects of GPCR research". Molecular interventions 7 (1): 17–25. doi:10.1124/mi.7.1.5. PMID 17339603.
- ^ Terakita, A. (2005). "The opsins". Genome biology 6 (3): 213. doi:10.1186/gb-2005-6-3-213. PMC 1088937. PMID 15774036. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1088937.
- ^ Murakami, M.; Kouyama, T. (2008). "Crystal structure of squid rhodopsin". Nature 453 (7193): 363. Bibcode 2008Natur.453..363M. doi:10.1038/nature06925. PMID 18480818.
- ^ Smith, W. C.; Price, D. A.; Greenberg, R. M.; Battelle, B. A. (1993). "Opsins from the lateral eyes and ocelli of the horseshoe crab, Limulus polyphemus". Proceedings of the National Academy of Sciences of the United States of America 90 (13): 6150–6154. Bibcode 1993PNAS...90.6150S. doi:10.1073/pnas.90.13.6150. PMC 46885. PMID 8327495. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=46885.
- ^ Okano T, Yoshizawa T, Fukada Y (1994). "Pinopsin is a chicken pineal photoreceptive molecule". Nature 372 (6501): 94–97. doi:10.1038/372094a0. PMID 7969427.
- ^ Philp AR, Garcia-Fernandez JM, Soni BG, Lucas RJ, Bellingham J, Foster RG (2000). "Vertebrate ancient (VA) opsin and extraretinal photoreception in the Atlantic salmon (Salmo salar)". J. Exp. Biol. 203 (Pt 12): 1925–36. PMID 10821749.
- ^ Blackshaw S, Snyder SH (1997). "Parapinopsin, a novel catfish opsin localized to the parapineal organ, defines a new gene family". J. Neurosci. 17 (21): 8083–92. PMID 9334384.
- ^ Mano H, Kojima D, Fukada Y (1999). "Exo-rhodopsin: a novel rhodopsin expressed in the zebrafish pineal gland". Brain Res. Mol. Brain Res. 73 (1–2): 110–118. doi:10.1016/S0169-328X(99)00242-9. PMID 10581404.
- ^ Moutsaki P, Whitmore D, Bellingham J, Sakamoto K, David-Gray ZK, Foster RG (2003). "Teleost multiple tissue (tmt) opsin: a candidate photopigment regulating the peripheral clocks of zebrafish?". Brain Res. Mol. Brain Res. 112 (1–2): 135–145. doi:10.1016/S0169-328X(03)00059-7. PMID 12670711.
- ^ Palczewski K et al. (2000). "Crystal Structure of Rhodopsin: A G Protein-Coupled Receptor". Science 289 (5480): 739–745. doi:10.1126/science.289.5480.739. PMID 10926528.
- ^ Yokoyama S (2000). "Molecular evolution of vertebrate visual pigments". Progress in Retinal and Eye Research 19 (4): 385–419. doi:10.1016/S1350-9462(00)00002-1. PMID 10785616.
- ^ Deeb SS (2005). "The molecular basis of variation in human color vision". Clinical genetics 67 (5): 369–377. doi:10.1111/j.1399-0004.2004.00343.x. PMID 15811001.
Opsin (retinylidene protein) Crystallin Other M: EYE
anat(g/a/p)/phys/devp/prot
noco/cong/tumr, epon
proc, drug(S1A/1E/1F/1L)
Categories:- G protein coupled receptors
- Vision
Wikimedia Foundation. 2010.