- Ion channel
-
- Not to be confused with: Ion Television or Ion implantation.
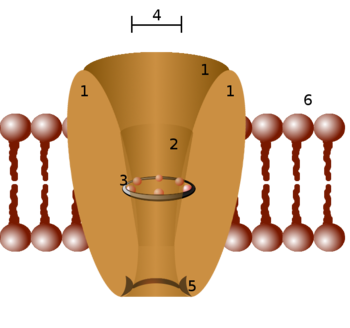 Schematic diagram of an ion channel. 1 - channel domains (typically four per channel), 2 - outer vestibule, 3 - selectivity filter, 4 - diameter of selectivity filter, 5 - phosphorylation site, 6 - cell membrane.
Schematic diagram of an ion channel. 1 - channel domains (typically four per channel), 2 - outer vestibule, 3 - selectivity filter, 4 - diameter of selectivity filter, 5 - phosphorylation site, 6 - cell membrane.
Ion channels are pore-forming proteins that help establish and control the small voltage gradient across the plasma membrane of cells (see cell potential) by allowing the flow of ions down their electrochemical gradient.[1] They are present in the membranes that surround all biological cells. The study of ion channels involves many scientific techniques such as voltage clamp electrophysiology (in particular patch clamp), immunohistochemistry, and RT-PCR.
Contents
Basic features
Ion channels regulate the flow of ions across the membrane in all cells. Ion channels are integral membrane proteins; or, more typically, an assembly of several proteins. They are present on all membranes of cell (plasma membrane) and intracellular organelles (nucleus, mitochondria, endoplasmic reticulum, golgi apparatus and so on). Such "multi-subunit" assemblies usually involve a circular arrangement of identical or homologous proteins closely packed around a water-filled pore through the plane of the membrane or lipid bilayer.[2][3] For most voltage-gated ion channels, the pore-forming subunit(s) are called the α subunit, while the auxiliary subunits are denoted β, γ, and so on. Some channels permit the passage of ions based solely on their charge of positive (cation) or negative (anion). However, the archetypal channel pore is just one or two atoms wide at its narrowest point and is selective for specific species of ion, such as sodium or potassium. These ions move through the channel pore single file nearly as quickly as the ions move through free fluid. In some ion channels, passage through the pore is governed by a "gate," which may be opened or closed by chemical or electrical signals, temperature, or mechanical force, depending on the variety of channel.
Biological role
Because channels underlie the nerve impulse and because "transmitter-activated" channels mediate conduction across the synapses, channels are especially prominent components of the nervous system. Indeed, most of the offensive and defensive toxins that organisms have evolved for shutting down the nervous systems of predators and prey (e.g., the venoms produced by spiders, scorpions, snakes, fish, bees, sea snails and others) work by modulating ion channel conductance and/or kinetics. In addition, ion channels are key components in a wide variety of biological processes that involve rapid changes in cells, such as cardiac, skeletal, and smooth muscle contraction, epithelial transport of nutrients and ions, T-cell activation and pancreatic beta-cell insulin release. In the search for new drugs, ion channels are a frequent target.[4][5][6]
Diversity
There are over 300 types of ion channels in a living cell.[7] Ion channels may be classified by the nature of their gating, the species of ions passing through those gates, the number of gates (pores) and localization of proteins.
Further heterogeneity of ion channels arises when channels with different constitutive subunits give rise to a specific kind of current.[8] Absence or mutation of one or more of the contributing types of channel subunits can result in loss of function and, potentially, underlie neurologic diseases.
By gating
Ion channels may be classified by gating, i.e. what opens and closes the channels. Voltage-gated ion channels open or close depending on the voltage gradient across the plasma membrane, while ligand-gated ion channels open or close depending on binding of ligands to the channel.[citation needed]
Voltage-gated
Main article: Voltage-gated ion channelVoltage-gated ion channels open and close in response to membrane potential.
- Voltage-gated sodium channels: This family contains at least 9 members and is largely responsible for action potential creation and propagation. The pore-forming α subunits are very large (up to 4,000 amino acids) and consist of four homologous repeat domains (I-IV) each comprising six transmembrane segments (S1-S6) for a total of 24 transmembrane segments. The members of this family also coassemble with auxiliary β subunits, each spanning the membrane once. Both α and β subunits are extensively glycosylated.
- Voltage-gated calcium channels: This family contains 10 members, though these members are known to coassemble with α2δ, β, and γ subunits. These channels play an important role in both linking muscle excitation with contraction as well as neuronal excitation with transmitter release. The α subunits have an overall structural resemblance to those of the sodium channels and are equally large.
- Cation channels of sperm: This small family of channels, normally referred to as Catsper channels, is related to the two-pore channels and distantly related to TRP channels.
- Voltage-gated potassium channels (KV): This family contains almost 40 members, which are further divided into 12 subfamilies. These channels are known mainly for their role in repolarizing the cell membrane following action potentials. The α subunits have six transmembrane segments, homologous to a single domain of the sodium channels. Correspondingly, they assemble as tetramers to produce a functioning channel.
- Some transient receptor potential channels: This group of channels, normally referred to simply as TRP channels, is named after their role in Drosophila phototransduction. This family, containing at least 28 members, is incredibly diverse in its method of activation. Some TRP channels seem to be constitutively open, while others are gated by voltage, intracellular Ca2+, pH, redox state, osmolarity, and mechanical stretch. These channels also vary according to the ion(s) they pass, some being selective for Ca2+ while others are less selective, acting as cation channels. This family is subdivided into 6 subfamilies based on homology: classical (TRPC), vanilloid receptors (TRPV), melastatin (TRPM), polycystins (TRPP), mucolipins (TRPML), and ankyrin transmembrane protein 1 (TRPA).
- Hyperpolarization-activated cyclic nucleotide-gated channels: The opening of these channels is due to hyperpolarization rather than the depolarization required for other cyclic nucleotide-gated channels. These channels are also sensitive to the cyclic nucleotides cAMP and cGMP, which alter the voltage sensitivity of the channel’s opening. These channels are permeable to the monovalent cations K+ and Na+. There are 4 members of this family, all of which form tetramers of six-transmembrane α subunits. As these channels open under hyperpolarizing conditions, they function as pacemaking channels in the heart, particularly the SA node.
- Voltage-gated proton channels: Voltage-gated proton channels open with depolarization, but in a strongly pH-sensitive manner. The result is that these channels open only when the electrochemical gradient is outward, such that their opening will only allow protons to leave cells. Their function thus appears to be acid extrusion from cells. Another important function occurs in phagocytes (e.g. eosinophils, neutrophils, macrophages) during the "respiratory burst." When bacteria or other microbes are engulfed by phagocytes, the enzyme NADPH oxidase assembles in the membrane and begins to produce reactive oxygen species (ROS) that help kill bacteria. NADPH oxidase is electrogenic, moving electrons across the membrane, and proton channels open to allow proton flux to balance the electron movement electrically.
Ligand-gated
Main article: Ligand-gated ion channelAlso known as ionotropic receptors, this group of channels open in response to specific ligand molecules binding to the extracellular domain of the receptor protein. Ligand binding causes a conformational change in the structure of the channel protein that ultimately leads to the opening of the channel gate and subsequent ion flux across the plasma membrane. Examples of such channels include the cation-permeable "nicotinic" Acetylcholine receptor, ionotropic glutamate-gated receptors and ATP-gated P2X receptors, and the anion-permeable γ-aminobutyric acid-gated GABAA receptor.
Ion channels activated by second messengers may also be categorized in this group, although ligands and second messengers are otherwise distinguished from each other.
Other gating
Other gating include activation/inactivation by e.g. second messengers from the inside of the cell membrane, rather as from outside, as in the case for ligands. Ions may count to such second messengers, and then causes direct activation, rather than indirect, as in the case were the electric potential of ions cause activation/inactivation of voltage-gated ion channels.
- Some potassium channels
- Inward-rectifier potassium channels: These channels allow potassium to flow into the cell in an inwardly rectifying manner, i.e., potassium flows effectively into, but not out of, the cell. This family is composed of 15 official and 1 unofficial members and is further subdivided into 7 subfamilies based on homology. These channels are affected by intracellular ATP, PIP2, and G-protein βγ subunits. They are involved in important physiological processes such as the pacemaker activity in the heart, insulin release, and potassium uptake in glial cells. They contain only two transmembrane segments, corresponding to the core pore-forming segments of the KV and KCa channels. Their α subunits form tetramers.
- Calcium-activated potassium channels: This family of channels is, for the most part, activated by intracellular Ca2+ and contains 8 members.
- Two-pore-domain potassium channels: This family of 15 members form what is known as leak channels, and they follow Goldman-Hodgkin-Katz (open) rectification.
- Light-gated channels like channelrhodopsin are directly opened by the action of light.
- Mechanosensitive ion channels are opening under the influence of stretch, pressure, shear, displacement.
- Cyclic nucleotide-gated channels: This superfamily of channels contains two families: the cyclic nucleotide-gated (CNG) channels and the hyperpolarization-activated, cyclic nucleotide-gated (HCN) channels. It should be noted that this grouping is functional rather than evolutionary.
- Cyclic nucleotide-gated channels: This family of channels is characterized by activation due to the binding of intracellular cAMP or cGMP, with specificity varying by member. These channels are primarily permeable to monovalent cations such as K+ and Na+. They are also permeable to Ca2+, though it acts to close them. There are 6 members of this family, which is divided into 2 subfamilies.
- Hyperpolarization-activated cyclic nucleotide-gated channels
- Temperature Gated Channels: Members of the Transient Receptor Potential ion channel superfamily, such as TRPV1 or TRPM8 are opened either by hot or cold temperatures.
`
By ions
- Chloride channels: This superfamily of poorly-understood channels consists of approximately 13 members. They include ClCs, CLICs, Bestrophins and CFTRs. These channels are non-selective for small anions; however chloride is the most abundant anion, and hence they are known as chloride channels.
- Potassium channels
- Voltage-gated potassium channels e.g., Kvs, Kirs etc.
- Calcium-activated potassium channels e.g., BKCa or MaxiK, SK, etc.
- Inward-rectifier potassium channels
- Two-pore-domain potassium channels: This family of 15 members form what is known as leak channels, and they follow Goldman-Hodgkin-Katz (open) rectification.
- Sodium channels
- voltage-gated sodium channels NaVs
- epithelial sodium channels (ENaC)
- Calcium channels CaVs
- Proton channels
- Voltage-gated proton channels
- Non-selective cation channels: These let many types of cations, mainly Na+, K+ and Ca2+ through the channel.
- Most Transient receptor potential channels
Other classifications
There are other types of ion channel classifications that are based on less normal characteristics, e.g. multiple pores and transient potentials.
Almost all ion channels have one single pore. However, there are also those with two:
- Two-pore channels: This small family of 2 members putatively forms cation-selective ion channels. They are predicted to contain two KV-style six-transmembrane domains, suggesting they form a dimer in the membrane. These channels are related to catsper channels channels and, more distantly, TRP channels.
There are channels that are classified by the duration of the response to stimuli:
- Transient receptor potential channels: This group of channels, normally referred to simply as TRP channels, is named after their role in Drosophila phototransduction. This family, containing at least 28 members, is incredibly diverse in its method of activation. Some TRP channels seem to be constitutively open, while others are gated by voltage, intracellular Ca2+, pH, redox state, osmolarity, and mechanical stretch. These channels also vary according to the ion(s) they pass, some being selective for Ca2+ while others are less selective, acting as cation channels. This family is subdivided into 6 subfamilies based on homology: canonical (TRPC), vanilloid receptors (TRPV), melastatin (TRPM), polycystins (TRPP), mucolipins (TRPML), and ankyrin transmembrane protein 1 (TRPA).
Detailed structure
Channels differ with respect to the ion they let pass (for example, Na+, K+, Cl−), the ways in which they may be regulated, the number of subunits of which they are composed and other aspects of structure. Channels belonging to the largest class, which includes the voltage-gated channels that underlie the nerve impulse, consists of four subunits with six transmembrane helices each. On activation, these helices move about and open the pore. Two of these six helices are separated by a loop that lines the pore and is the primary determinant of ion selectivity and conductance in this channel class and some others. The existence and mechanism for ion selectivity was first postulated in the 1960s by Clay Armstrong.[9] He suggested that the pore lining could efficiently replace the water molecules that normally shield potassium ions, but that sodium ions were too small to allow such shielding, and therefore could not pass through. This mechanism was finally confirmed when the structure of the channel was elucidated. The channel subunits of one such other class, for example, consist of just this "P" loop and two transmembrane helices. The determination of their molecular structure by Roderick MacKinnon using X-ray crystallography won a share of the 2003 Nobel Prize in Chemistry.
Because of their small size and the difficulty of crystallizing integral membrane proteins for X-ray analysis, it is only very recently that scientists have been able to directly examine what channels "look like." Particularly in cases where the crystallography required removing channels from their membranes with detergent, many researchers regard images that have been obtained as tentative. An example is the long-awaited crystal structure of a voltage-gated potassium channel, which was reported in May 2003.[10][11] One inevitable ambiguity about these structures relates to the strong evidence that channels change conformation as they operate (they open and close, for example), such that the structure in the crystal could represent any one of these operational states. Most of what researchers have deduced about channel operation so far they have established through electrophysiology, biochemistry, gene sequence comparison and mutagenesis.
Channels can have single (CLICs) to multiple transmembrane (K channels, P2X receptors, Na channels) domains which span plasma membrane to form pores. Pore can determine the selectivity of the channel. Gate can be formed either inside or outside the pore region.
Diseases of ion channels
There are a number of chemicals and genetic disorders which disrupt normal functioning of ion channels and have disastrous consequences for the organism. Genetic disorders of ion channels and their modifiers are known as Channelopathies. See Category:Channelopathy for a full list.
Chemicals
- Tetrodotoxin (TTX), used by puffer fish and some types of newts for defense. It blocks sodium channels.
- Saxitoxin, is produced by a dinoflagellate also known as "red tide". It blocks voltage dependent sodium channels.
- Conotoxin, is used by cone snails to hunt prey.
- Lidocaine and Novocaine belong to a class of local anesthetics which block sodium ion channels.
- Dendrotoxin is produced by mamba snakes, and blocks potassium channels.
- Iberiotoxin is produced by the Buthus tamulus and blocks potassium channels.
- Heteropodatoxin is produced by Heteropoda venatoria and blocks potassium channels.
Genetic
- Shaker gene mutations cause a defect in the voltage gated ion channels, slowing down the repolarization of the cell.
- Equine hyperkalaemic periodic paralysis as well as Human hyperkalaemic periodic paralysis (HyperPP) are caused by a defect in voltage dependent sodium channels.
- Paramyotonia congenita (PC) and potassium aggravated myotonias (PAM)
- Generalized epilepsy with febrile seizures plus (GEFS+)
- Episodic Ataxia (EA), characterized by sporadic bouts of severe discoordination with or without myokymia, and can be provoked by stress, startle, or heavy exertion such as exercise.
- Familial hemiplegic migraine (FHM)
- Spinocerebellar ataxia type 13
- Long QT syndrome is a ventricular arrhythmia syndrome caused by mutations in one or more of presently ten different genes, most of which are potassium channels and all of which affect cardiac repolarization.
- Brugada syndrome is another ventricular arrhythmia caused by voltage-gated sodium channel gene mutations.
- Cystic fibrosis is caused by mutations in the CFTR gene, which is a chloride channel.
- Mucolipidosis type IV is caused by mutations in the gene encoding the TRPML1 channel
History
The fundamental properties of currents mediated by ion channels were analyzed by the British biophysicists Alan Hodgkin and Andrew Huxley as part of their Nobel Prize-winning research on the action potential, published in 1952. They built on the work of other physiologists, such as Cole and Baker's research into voltage-gated membrane pores from 1941.[12][13] The existence of ion channels was confirmed in the 1970s by Bernard Katz and Ricardo Miledi using noise analysis. It was then shown more directly with an electrical recording technique known as the "patch clamp", which led to a Nobel Prize to Erwin Neher and Bert Sakmann, the technique's inventors. Hundreds if not thousands of researchers continue to pursue a more detailed understanding of how these proteins work. In recent years the development of automated patch clamp devices helped to increase significantly the throughput in ion channel screening.
The Nobel Prize in Chemistry for 2003 was awarded to two American scientists: Roderick MacKinnon for his studies on the physico-chemical properties of ion channel structure and function, including x-ray crystallographic structure studies, and Peter Agre for his similar work on aquaporins.[14]
The ion channel in fine art
 Birth of an Idea (2007) by Julian Voss-Andreae. The sculpture was commissioned by Roderick MacKinnon based on the molecule's atomic coordinates that were determined by MacKinnon's group in 2001.
Birth of an Idea (2007) by Julian Voss-Andreae. The sculpture was commissioned by Roderick MacKinnon based on the molecule's atomic coordinates that were determined by MacKinnon's group in 2001.Roderick MacKinnon commissioned Birth of an Idea, a 5-foot (1.5 m) tall sculpture based on the KcsA potassium channel.[15] The artwork contains a wire object representing the channel's interior with a blown glass object representing the main cavity of the channel structure.
See also
- Action potential
- Active transport
- Alpha helix
- Channelome
- Channelomics
- Channelopathy
- Electrochemical gradient
- Gating (electrophysiology)
- Ion channel family as defined in Pfam and InterPro
- Ki Database
- Lipid bilayer ion channels
- Magnesium transport
- Membrane potential
- Neurotoxin
- Passive transport
- Potassium ion channels
- Sodium ion channel
- Synthetic ion channels
- Transmembrane receptor
- MeSH entry for Ion channels
References
- ^ Hille, Bertil (2001). Ion channels of excitable membranes (third ed.). Sunderland, Mass: Sinauer Associates. ISBN 0-87893-321-2.
- ^ Purves, (2001). "Chapter 4: Channels and Transporters". In Dale Purves, George J. Augustine, David Fitzpatrick, Lawrence. C. Katz, Anthony-Samuel LaMantia, James O. McNamara, S. Mark Williams, editors. Neuroscience (2nd ed.). Sinauer Associates Inc.. ISBN 0-87893-741-2. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=neurosci.chapter.227.
- ^ Hille B, Catterall, WA (1999). "Chapter 6: Electrical Excitability and Ion Channels". In George J Siegel, Bernard W Agranoff, R. W Albers, Stephen K Fisher and Michael D Uhler. Basic neurochemistry: molecular, cellular, and medical aspects. Philadelphia: Lippincott-Raven. ISBN 0-397-51820-X. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=bnchm.chapter.421.
- ^ Camerino DC, Tricarico D, Desaphy JF (April 2007). "Ion channel pharmacology". Neurotherapeutics 4 (2): 184–98. doi:10.1016/j.nurt.2007.01.013. PMID 17395128.
- ^ Verkman AS, Galietta LJ (February 2009). "Chloride channels as drug targets". Nat Rev Drug Discov 8 (2): 153–71. doi:10.1038/nrd2780. PMID 19153558.
- ^ Camerino DC, Desaphy JF, Tricarico D, Pierno S, Liantonio A (2008). "Therapeutic approaches to ion channel diseases". Adv. Genet.. Advances in Genetics 64: 81–145. doi:10.1016/S0065-2660(08)00804-3. ISBN 9780123746214. PMID 19161833.
- ^ Gabashvili IS, Sokolowski BH, Morton CC, Giersch AB (September 2007). "Ion Channel Gene Expression in the Inner Ear". J. Assoc. Res. Otolaryngol. 8 (3): 305–28. doi:10.1007/s10162-007-0082-y. PMC 2538437. PMID 17541769. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2538437.
- ^ Vicini S (April 1999). "New perspectives in the functional role of GABA(A) channel heterogeneity". Mol. Neurobiol. 19 (2): 97–110. doi:10.1007/BF02743656. PMID 10371465.
- ^ Bezanilla F, Armstrong CM (November 1972). "Negative Conductance Caused by Entry of Sodium and Cesium Ions into the Potassium Channels of Squid Axons". J. Gen. Physiol. 60 (5): 588–608. doi:10.1085/jgp.60.5.588. PMC 2226091. PMID 4644327. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2226091.
- ^ Jiang Y, Lee A, Chen J, Ruta V, Cadene M, Chait BT, MacKinnon R (May 2003). "X-ray structure of a voltage-dependent K+ channel". Nature 423 (6935): 33–41. doi:10.1038/nature01580. PMID 12721618.
- ^ The detailed 3D structure of the magnesium channel from bacteria can be seen here [1].
- ^ Ronald Pethig and Douglas B Kell (1987). "The passive electrical properties of biological systems: their significance in physiology, biophysics and biotechnology". Phys. Med. Biol. 32 (8): 933–970. Bibcode 1987PMB....32..933P. doi:10.1088/0031-9155/32/8/001. PMID 3306721. http://dbkgroup.org/Papers/pethig_kell_pmb87.pdf. [An expansive review of bioelectrical characteristics from 1987. Lay summary]. "... the observation of an inductance (negative capacitance) by Cole and Baker (1941) during measurements of the AC electrical properties of squid axons led directly to the concept of voltage-gated membrane pores, as embodied in the celebrated Hodgkin-Huxley (1952) treatment (Cole 1972, Jack er a1 1975), as the crucial mechanism of neurotransmission."
- ^ Kenneth S. Cole and Richard F. Baker (February 1941). "LONGITUDINAL IMPEDANCE OF THE SQUID GIANT AXON". The Journal of General Physiology (The Rockefeller University Press) 24 (6): 771–88. doi:10.1085/jgp.24.6.771. PMC 2238007. PMID 19873252. http://jgp.rupress.org/cgi/reprint/24/6/771. [Describes what happens when you stick a giant squid axon with electrodes and pass through an alternating current, and then notice that sometimes the voltage rises with time, and sometimes it decreases. Lay summary]. "The inductive reactance is a property of the axon and requires that it contain an inductive structure. The variation of the impedance with interpolar distance indicates that the inductance is in the membrane"
- ^ "The Nobel Prize in Chemistry 2003" (Press release). The Royal Swedish Academy of Science. 2003-10-08. http://nobelprize.org/nobel_prizes/chemistry/laureates/2003/press.html. Retrieved 2010-01-18.
- ^ Ball, Philip (March 2008). "The crucible: Art inspired by science should be more than just a pretty picture". Chemistry World 5 (3): 42–43. http://www.rsc.org/chemistryworld/Issues/2008/March/ColumnThecrucible.asp. Retrieved 2009-01-12.
Further reading
- An Interview with Roderick MacKinnon Freeview video by the Vega Science Trust.
- Doyle DA, Morais Cabral J, Pfuetzner RA, Kuo A, Gulbis JM, Cohen SL, Chait BT, MacKinnon R (April 1998). "The structure of the potassium channel: molecular basis of K+ conduction and selectivity". Science 280 (5360): 69–77. Bibcode 1998Sci...280...69D. doi:10.1126/science.280.5360.69. PMID 9525859. http://www.cellbio.wustl.edu/faculty/huettner/69.pdf.
- Bezanilla F (April 2000). "The voltage sensor in voltage-dependent ion channels". Physiol. Rev. 80 (2): 555–92. PMID 10747201. http://physrev.physiology.org/cgi/content/abstract/80/2/555.
- Singh H (2010). "Two decades with dimorphic Chloride Intracellular Channels (CLICs)". Febs letters. 584 (10): 2112–21. doi:10.1016/j.febslet.2010.03.013. PMID 20226783.
- Catterall WA (2010). "Ion Channel Voltage Sensors: Structure, Function, and Pathophysiology". Neuron 67 (6): 915–928. doi:10.1016/j.neuron.2010.08.021. PMC 2950829. PMID 20869590. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2950829.
- Kathy Liszewski (2007-06-15). "Opening the Gates on Ion Channel Drugs". Genetic Engineering & Biotechnology News (Mary Ann Liebert, Inc.): pp. 18, 20. http://www.genengnews.com/articles/chitem.aspx?aid=2159. Retrieved 2008-07-06. "Conditions targeted include diabetes, cystic fibrosis, hypertension, pain and cancer"
- Shahidul, Islam (January 2011). Transient Receptor Potential Channels. Advances in Experimental Medicine and Biology. 704. Berlin: Springer. p. 700. ISBN 978-94-007-0264-6.
External links
- "Voltage-Gated Ion Channels". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology. http://www.iuphar-db.org/DATABASE/ReceptorFamiliesForward?type=IC.
- "TRIP Database". a manually curated database of protein-protein interactions for mammalian TRP channels. http://www.trpchannel.org.
- AurSCOPE Ion Channels Database
Ca2+: Calcium channel Ligand-gatedNa+: Sodium channel Constitutively activeProton gatedK+: Potassium channel Kvα1-6 (1.1, 1.2, 1.3, 1.4, 1.5, 1.6, 1.7, 1.8) · (2.1, 2.2) · (3.1, 3.2, 3.3, 3.4) · (4.1, 4.2, 4.3) · (5.1) · (6.1, 6.2, 6.3, 6.4)
Kvα7-12 (7.1, 7.2, 7.3, 7.4, 7.5) · (8.1, 8.2) · (9.1, 9.2, 9.3) · (10.1, 10.2) · (11.1/hERG, 11.2, 11.3) · (12.1, 12.2, 12.3)
Kvβ (1, 2, 3) · KCNIP (1, 2, 3, 4) · minK/ISK · minK/ISK-like · MiRP (1, 2, 3) · Shaker geneOther Cl-: Chloride channelHVCN1GeneralIon channel, cell surface receptor: ligand-gated ion channels Cys-loop receptors monomers: α1 · α2 · α3 · α4 · α5 · α6 · α7 · α9 · α10 · β1 · β2 · β3 · β4 · δ · ε
pentamers: (α3)2(β4)3 · (α4)2(β2)3 · (α7)5 · (α1)2(β4)3 - Ganglion type · (α1)2β1δε - Muscle typeZincZinc-activatedIonotropic glutamates Ligand-gated onlyVoltage- and ligand-gated‘Orphan’ATP-gated channels Categories:- Cell communication
- Electrophysiology
- Integral membrane proteins
- Ion channels
- Neurochemistry
- Protein families

Wikimedia Foundation. 2010.