- Potassium channel
-
In the field of cell biology, potassium channels are the most widely distributed type of ion channel and are found in virtually all living organisms.[1] They form potassium-selective pores that span cell membranes. Furthermore potassium channels are found in most cell types and control a wide variety of cell functions.[2][3]
Contents
Function
In excitable cells such as neurons, they shape action potentials and set the resting membrane potential.
By contributing to the regulation of the action potential duration in cardiac muscle, malfunction of potassium channels may cause life-threatening arrhythmias. Potassium channels may also be involved in maintaining vascular tone.
They also regulate cellular processes such as the secretion of hormones (e.g., insulin release from beta-cells in the pancreas) so their malfunction can lead to diseases (such as diabetes).
Types
There are four major classes of potassium channels:
- Calcium-activated potassium channel - open in response to the presence of calcium ions or other signalling molecules.
- Inwardly rectifying potassium channel - passes current (positive charge) more easily in the inward direction (into the cell).
- Tandem pore domain potassium channel - are constitutively open or possess high basal activation, such as the "resting potassium channels" or "leak channels" that set the negative membrane potential of neurons. When open, they allow potassium ions to cross the membrane at a rate that is nearly as fast as their diffusion through bulk water.
- Voltage-gated potassium channel - are voltage-gated ion channels that open or close in response to changes in the transmembrane voltage.
The following table contains a comparison of the major classes of potassium channels with representative examples (for a complete list of channels within each class, see the respective class pages).
Potassium channel classes, function, and pharmacology.[4] Class Subclasses Function Blockers Activators Calcium-activated
6T & 1P- inhibition following stimuli increasing intracellular calcium
- 1-EBIO
- NS309
- CyPPA
Inwardly rectifying
2T & 1P- ROMK (Kir1.1)
- recycling and secretion of potassium in nephrons
- Nonselective: Ba2+, Cs+
- none
- GPCR regulated (Kir3.x)
- mediate the inhibitory effect of many GPCRs
- GPCR antagonists
- ifenprodil[5]
- GPCR agonists
- ATP-sensitive (Kir6.x)
Tandem pore domain
4T & 2P- Contribute to resting potential
Voltage-gated
6T & 1P- action potential repolarization
- limits frequency of action potentials (disturbances cause dysrhythmia)
- tetraethylammonium
- 4-aminopyridine
- dendrotoxins (some types)
- retigabine (Kv7)[22]
Structure
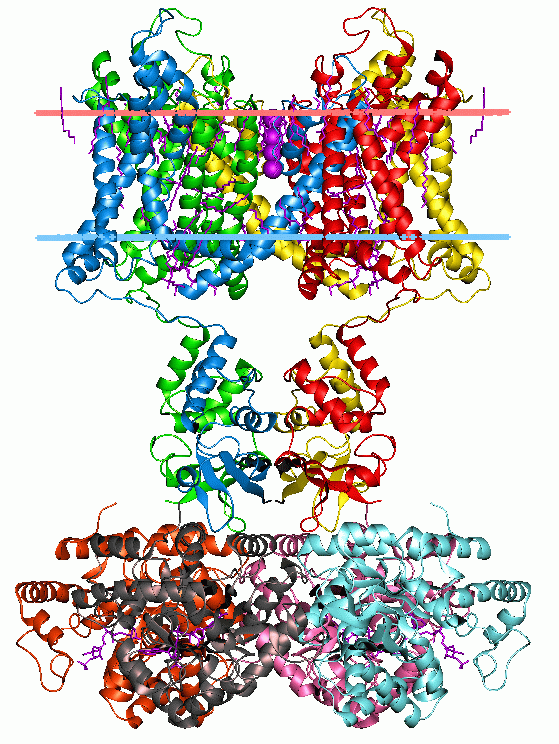
 Potassium channel KvAP, structure in a membrane-like environment. Calculated hydrocarbon boundaries of the lipid bilayer are indicated by red and blue lines.
Potassium channel KvAP, structure in a membrane-like environment. Calculated hydrocarbon boundaries of the lipid bilayer are indicated by red and blue lines.
Potassium channels have a tetrameric structure in which four identical protein subunits associate to form a fourfold symmetric (C4) complex arranged around a central ion conducting pore (i.e., a homotetramer). Alternatively four related but not identical protein subunits may associate to form heterotetrameric complexes with pseudo C4 symmetry. All potassium channel subunits have a distinctive pore-loop structure that lines the top of the pore and is responsible for potassium selective permeability.
There are over 80 mammalian genes that encode potassium channel subunits. However potassium channels found in bacteria are amongst the most studied of ion channels, in terms of their molecular structure. Using X-ray crystallography,[23][24] profound insights have been gained into how potassium ions pass through these channels and why (smaller) sodium ions do not.[25] The 2003 Nobel Prize for Chemistry was awarded to Rod MacKinnon for his pioneering work in this area.[26]
Selectivity filter
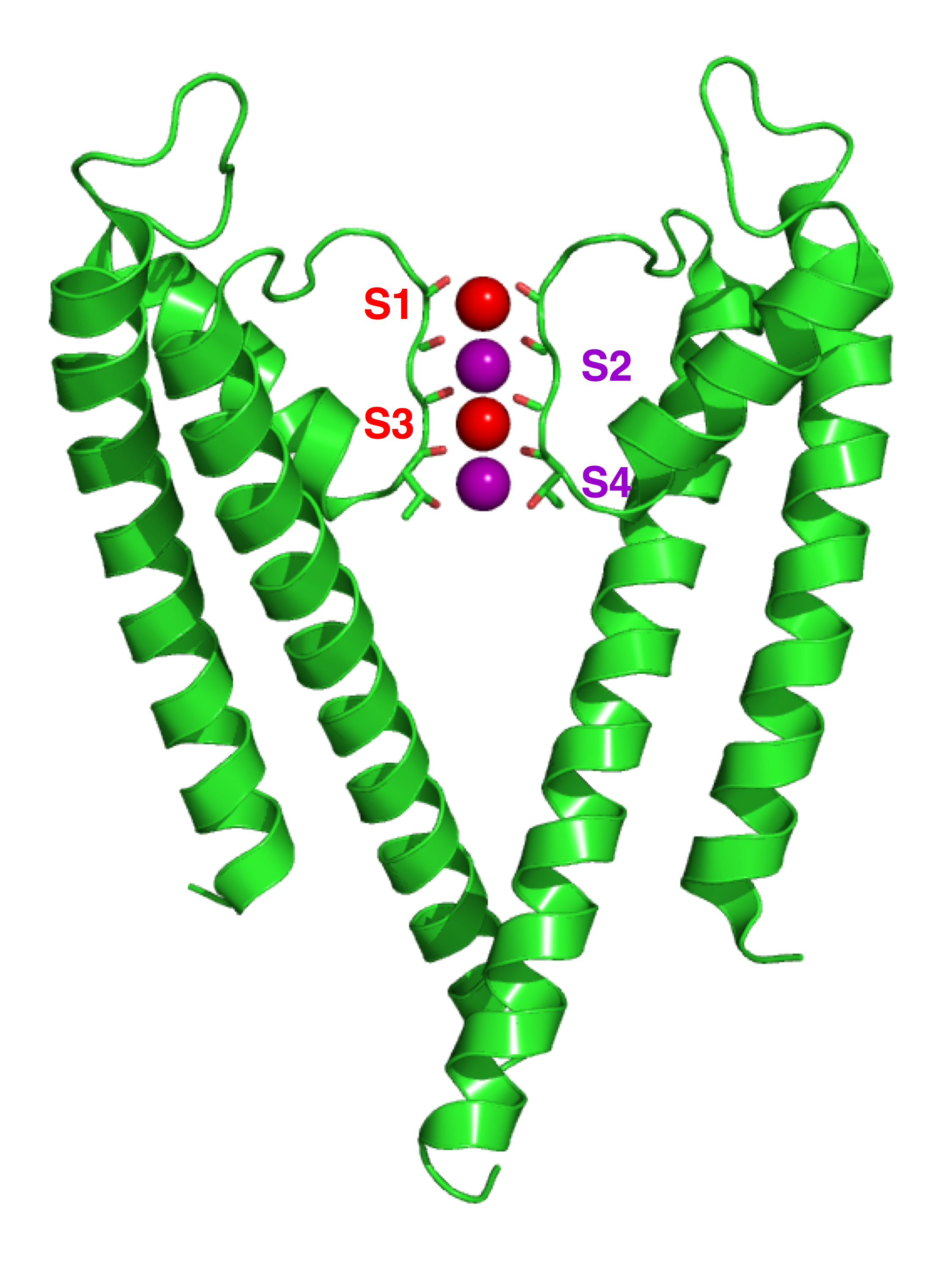
 Crystallographic structure of the bacterial KcsA potassium channel (PDB 1K4C).[27] In this figure, only two of the four subunits of the tetramer are displayed for the sake of clarity. The protein is displayed as a green cartoon diagram. In addition backbone carbonyl groups and threonine sidechain protein atoms (oxygen = red, carbon = green) are displayed. Finally potassium ions (occupying the S2 and S4 sites) and the oxygen atoms of water molecules (S1 and S3) are depicted as purple and red spheres respectively.
Crystallographic structure of the bacterial KcsA potassium channel (PDB 1K4C).[27] In this figure, only two of the four subunits of the tetramer are displayed for the sake of clarity. The protein is displayed as a green cartoon diagram. In addition backbone carbonyl groups and threonine sidechain protein atoms (oxygen = red, carbon = green) are displayed. Finally potassium ions (occupying the S2 and S4 sites) and the oxygen atoms of water molecules (S1 and S3) are depicted as purple and red spheres respectively.Potassium ion channels remove the hydration shell from the ion when it enters the selectivity filter. The selectivity filter is formed by five residues (TVGYG-in prokaryotic species) in the P loop from each subunit, which have their electro-negative carbonyl oxygen atoms aligned toward the centre of the filter pore and form an anti-prism similar to a water-solvating shell around each potassium binding site. The distance between the carbonyl oxygens and potassium ions in the binding sites of the selectivity filter is the same as between water oxygens in the first hydration shell and a potassium ion in water solution. Passage of sodium ions would be energetically unfavorable, since the strong interactions between the filter and pore helix would prevent the channel from collapsing to the smaller sodium ion size. The selectivity filter opens towards the extracellular solution, exposing four carbonyl oxygens in a glycine residue (Gly79 in KcsA). The next residue toward the extracellular side of the protein is the negatively charged Asp80 (KcsA). This residue together with the five filter residues form the pore that connects the water-filled cavity in the centre of the protein with the extracellular solution.[28]
The carbonyl oxygens are strongly electro-negative and cation-attractive. The filter can accommodate potassium ions at 4 sites usually labelled S1 to S4 starting at the extracellular side. In addition, one ion can bind in the cavity at a site called SC or one or more ions at the extracellular side at more or less well-defined sites called S0 or Sext. Several different occupancies of these sites are possible. Since the X-ray structures are averages over many molecules, it is, however, not possible to deduce the actual occupancies directly from such a structure. In general, there is some disadvantage due to electrostatic repulsion to have two neighbouring sites occupied by ions. The mechanism for ion translocation in KcsA has been studied extensively by simulation techniques. A complete map of the free energies of the 24=16 states (characterised by the occupancy of the S1, S2, S3, and S4 sites) has been calculated with molecular dynamics simulations, resulting in the prediction of an ion conduction mechanism in which the two doubly occupied states (S1, S3) and (S2, S4) play an essential role. The two extracellular states, Sext and S0, were found in a better resolved structure of KcsA at high potassium concentration. In free energy calculations, the entire ionic pathway from the cavity through the four filter sites out to S0 and Sext was covered in MD simulations. The amino acids sequence of the selectivity filter of potassium ion channels is conserved, with the exception that an isoleucine residue in eukaryotic potassium ion channels often is substituted with a valine residue in prokaryotic channels.[28]
Hydrophobic region
This region is used to neutralize the environment around the potassium ion so that it is not attracted to any charges. In turn, it speeds up the reaction.
Central cavity
A central pore, 10 Å wide, is located near the center of the transmembrane channel, where the energy barrier is highest for the transversing ion due to the hydrophobity of the channel wall. The water-filled cavity and the polar C-terminus of the pore helices ease the energetic barrier for the ion. Repulsion by preceding multiple potassium ions is thought to aid the throughput of the ions. The presence of the cavity can be understood intuitively as one of the channel's mechanisms for overcoming the dielectric barrier, or repulsion by the low-dielectric membrane, by keeping the K+ ion in a watery, high-dielectric environment.
Blockers
Potassium channel blockers, such as 4-aminopyridine and 3,4-diaminopyridine, have been investigated for the treatment of conditions such as multiple sclerosis.[29]
Muscarinic potassium channel
See also G protein-coupled inwardly-rectifying potassium channel
Some types of potassium channels are activated by muscarinic receptors and these are called muscarinic potassium channels (IKACh). These channels are a heterotetramer composed of two GIRK1 and two GIRK4 subunits.[30][31] Examples are potassium channels in the heart, which, when activated by parasympathetic signals through M2 muscarinic receptors, causes an outward current of potassium, which slows down the heart rate.[32][33]
Potassium channels in fine art
 Birth of an Idea (2007) by Julian Voss-Andreae. The sculpture was commissioned by Roderick MacKinnon based on the molecule's atomic coordinates that were determined by MacKinnon's group in 2001.
Birth of an Idea (2007) by Julian Voss-Andreae. The sculpture was commissioned by Roderick MacKinnon based on the molecule's atomic coordinates that were determined by MacKinnon's group in 2001.Roderick MacKinnon commissioned Birth of an Idea, a 5-foot (1.5 m) tall sculpture based on the KcsA potassium channel.[34] The artwork contains a wire object representing the channel's interior with a blown glass object representing the main cavity of the channel structure.
See also
- Sodium ion channel
- Calcium channel
References
- ^ Littleton JT, Ganetzky B (2000). "Ion channels and synaptic organization: analysis of the Drosophila genome". Neuron 26 (1): 35–43. doi:10.1016/S0896-6273(00)81135-6. PMID 10798390.
- ^ Hille, Bertil (2001). "Chapter 5: Potassium Channels and Chloride Channels". Ion channels of excitable membranes. Sunderland, Mass: Sinauer. pp. 131–168. ISBN 0-87893-321-2.
- ^ Jessell, Thomas M.; Kandel, Eric R.; Schwartz, James H. (2000). "Chapter 6: Ion Channels". Principles of Neural Science (4th ed.). New York: McGraw-Hill. pp. 105–124. ISBN 0-8385-7701-6.
- ^ Rang, HP (2003). Pharmacology. Edinburgh: Churchill Livingstone. p. 60. ISBN 0-443-07145-4.
- ^ Kobayashi T, Washiyama K, Ikeda K (2006). "Inhibition of G protein-activated inwardly rectifying K+ channels by ifenprodil". Neuropsychopharmacology 31 (3): 516–24. doi:10.1038/sj.npp.1300844. PMID 16123769.
- ^ a b c d e f Enyedi P, Czirják G (2010). "Molecular background of leak K+ currents: two-pore domain potassium channels". Physiological Reviews 90 (2): 559–605. doi:10.1152/physrev.00029.2009. PMID 20393194.
- ^ a b c d e f Lotshaw DP (2007). "Biophysical, pharmacological, and functional characteristics of cloned and native mammalian two-pore domain K+ channels". Cell Biochemistry and Biophysics 47 (2): 209–56. doi:10.1007/s12013-007-0007-8. PMID 17652773.
- ^ Fink M, Lesage F, Duprat F, Heurteaux C, Reyes R, Fosset M, Lazdunski M (1998). "A neuronal two P domain K+ channel stimulated by arachidonic acid and polyunsaturated fatty acids". The EMBO Journal 17 (12): 3297–308. doi:10.1093/emboj/17.12.3297. PMC 1170668. PMID 9628867. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1170668.
- ^ Goldstein SA, Bockenhauer D, O'Kelly I, Zilberberg N (2001). "Potassium leak channels and the KCNK family of two-P-domain subunits". Nature Reviews Neuroscience 2 (3): 175–84. doi:10.1038/35058574. PMID 11256078.
- ^ Sano Y, Inamura K, Miyake A, Mochizuki S, Kitada C, Yokoi H, Nozawa K, Okada H, Matsushime H, Furuichi K (2003). "A novel two-pore domain K+ channel, TRESK, is localized in the spinal cord". The Journal of Biological Chemistry 278 (30): 27406–12. doi:10.1074/jbc.M206810200. PMID 12754259.
- ^ Czirják G, Tóth ZE, Enyedi P (2004). "The two-pore domain K+ channel, TRESK, is activated by the cytoplasmic calcium signal through calcineurin". The Journal of Biological Chemistry 279 (18): 18550–8. doi:10.1074/jbc.M312229200. PMID 14981085.
- ^ Kindler CH, Yost CS, Gray AT (1999). "Local anesthetic inhibition of baseline potassium channels with two pore domains in tandem". Anesthesiology 90 (4): 1092–102. doi:10.1097/00000542-199904000-00024. PMID 10201682.
- ^ a b c Meadows HJ, Randall AD (2001). "Functional characterisation of human TASK-3, an acid-sensitive two-pore domain potassium channel". Neuropharmacology 40 (4): 551–9. doi:10.1016/S0028-3908(00)00189-1. PMID 11249964.
- ^ Kindler CH, Paul M, Zou H, Liu C, Winegar BD, Gray AT, Yost CS (2003). "Amide local anesthetics potently inhibit the human tandem pore domain background K+ channel TASK-2 (KCNK5)". Journal of Pharmacology and Experimental Therapeutics 306 (1): 84–92. doi:10.1124/jpet.103.049809. PMID 12660311.
- ^ Punke MA, Licher T, Pongs O, Friederich P (2003). "Inhibition of human TREK-1 channels by bupivacaine". Anesthesia & Analgesia 96 (6): 1665–73. doi:10.1213/01.ANE.0000062524.90936.1F. PMID 12760993.
- ^ Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G, Barhanin J (1996). "TWIK-1, a ubiquitous human weakly inward rectifying K+ channel with a novel structure". The EMBO Journal 15 (5): 1004–11. PMC 449995. PMID 8605869. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=449995.
- ^ Duprat F, Lesage F, Fink M, Reyes R, Heurteaux C, Lazdunski M (1997). "TASK, a human background K+ channel to sense external pH variations near physiological pH". The EMBO Journal 16 (17): 5464–71. doi:10.1093/emboj/16.17.5464. PMC 1170177. PMID 9312005. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1170177.
- ^ Reyes R, Duprat F, Lesage F, Fink M, Salinas M, Farman N, Lazdunski M (1998). "Cloning and expression of a novel pH-sensitive two pore domain K+ channel from human kidney". The Journal of Biological Chemistry 273 (47): 30863–9. doi:10.1074/jbc.273.47.30863. PMID 9812978.
- ^ Meadows HJ, Benham CD, Cairns W, Gloger I, Jennings C, Medhurst AD, Murdock P, Chapman CG (2000). "Cloning, localisation and functional expression of the human orthologue of the TREK-1 potassium channel". Pflügers Archiv : European Journal of Physiology 439 (6): 714–22. doi:10.1007/s004240050997. PMID 10784345.
- ^ Patel AJ, Honoré E, Lesage F, Fink M, Romey G, Lazdunski M (1999). "Inhalational anesthetics activate two-pore-domain background K+ channels". Nature Neuroscience 2 (5): 422–6. doi:10.1038/8084. PMID 10321245.
- ^ Gray AT, Zhao BB, Kindler CH, Winegar BD, Mazurek MJ, Xu J, Chavez RA, Forsayeth JR, Yost CS (2000). "Volatile anesthetics activate the human tandem pore domain baseline K+ channel KCNK5". Anesthesiology 92 (6): 1722–30. doi:10.1097/00000542-200006000-00032. PMID 10839924.
- ^ Rogawski MA, Bazil CW (July 2008). "New Molecular Targets for Antiepileptic Drugs: α2δ, SV2A, and Kv7/KCNQ/M Potassium Channels". Curr Neurol Neurosci Rep 8 (4): 345–52. doi:10.1007/s11910-008-0053-7. PMC 2587091. PMID 18590620. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2587091.
- ^ Doyle DA, Morais Cabral J, Pfuetzner RA, Kuo A, Gulbis JM, Cohen SL, Chait BT, MacKinnon R (1998). "The structure of the potassium channel: molecular basis of K+ conduction and selectivity". Science 280 (5360): 69–77. Bibcode 1998Sci...280...69D. doi:10.1126/science.280.5360.69. PMID 9525859.
- ^ MacKinnon R, Cohen SL, Kuo A, Lee A, Chait BT (1998). "Structural conservation in prokaryotic and eukaryotic potassium channels". Science 280 (5360): 106–9. Bibcode 1998Sci...280..106M. doi:10.1126/science.280.5360.106. PMID 9525854.
- ^ Armstrong C (1998). "The vision of the pore". Science 280 (5360): 56–7. doi:10.1126/science.280.5360.56. PMID 9556453.
- ^ "The Nobel Prize in Chemistry 2003". The Nobel Foundation. http://nobelprize.org/nobel_prizes/chemistry/laureates/2003/. Retrieved 2007-11-16.
- ^ Zhou Y, Morais-Cabral JH, Kaufman A, MacKinnon R (2001). "Chemistry of ion coordination and hydration revealed by a K+ channel-Fab complex at 2.0 Â resolution". Nature 414 (6859): 43–8. doi:10.1038/35102009. PMID 11689936.
- ^ a b Hellgren M, Sandberg L, Edholm O (2006). "A comparison between two prokaryotic potassium channels (KirBac1.1 and KcsA) in a molecular dynamics (MD) simulation study". Biophys. Chem. 120 (1): 1–9. doi:10.1016/j.bpc.2005.10.002. PMID 16253415.
- ^ Judge SI, Bever CT (July 2006). "Potassium channel blockers in multiple sclerosis: neuronal Kv channels and effects of symptomatic treatment". Pharmacol. Ther. 111 (1): 224–59. doi:10.1016/j.pharmthera.2005.10.006. PMID 16472864. http://linkinghub.elsevier.com/retrieve/pii/S0163-7258(05)00228-7.
- ^ Krapivinsky G, Gordon EA, Wickman K, Velimirović B, Krapivinsky L, Clapham DE (1995). "The G-protein-gated atrial K+ channel IKACh is a heteromultimer of two inwardly rectifying K+-channel proteins". Nature 374 (6518): 135–41. Bibcode 1995Natur.374..135K. doi:10.1038/374135a0. PMID 7877685.
- ^ Corey S, Krapivinsky G, Krapivinsky L, Clapham DE (1998). "Number and stoichiometry of subunits in the native atrial G-protein-gated K+ channel, IKACh". J. Biol. Chem. 273 (9): 5271–8. doi:10.1074/jbc.273.9.5271. PMID 9478984.
- ^ Kunkel MT, Peralta EG (1995). "Identification of domains conferring G protein regulation on inward rectifier potassium channels". Cell 83 (3): 443–9. doi:10.1016/0092-8674(95)90122-1. PMID 8521474.
- ^ Wickman K, Krapivinsky G, Corey S, Kennedy M, Nemec J, Medina I, Clapham DE (1999). "Structure, G protein activation, and functional relevance of the cardiac G protein-gated K+ channel, IKACh". Ann. N. Y. Acad. Sci. 868 (1): 386–98. Bibcode 1999NYASA.868..386W. doi:10.1111/j.1749-6632.1999.tb11300.x. PMID 10414308. http://www.annalsnyas.org/cgi/content/abstract/868/1/386.
- ^ Ball, Philip (March 2008). "The crucible: Art inspired by science should be more than just a pretty picture". Chemistry World 5 (3): 42–43. http://www.rsc.org/chemistryworld/Issues/2008/March/ColumnThecrucible.asp. Retrieved 2009-01-12.
External links
- Proteopedia Potassium_channel in 3D
- MeSH Potassium+Channels
- Neuromuscular Disease Center (2008-03-04). "Potassium Channels". Washington University in St. Louis. http://neuromuscular.wustl.edu/mother/chan.html#k. Retrieved 2008-03-10.
- UMich Orientation of Proteins in Membranes families/superfamily-8
Ca2+: Calcium channel Ligand-gatedNa+: Sodium channel Constitutively activeProton gatedK+: Potassium channel Kvα1-6 (1.1, 1.2, 1.3, 1.4, 1.5, 1.6, 1.7, 1.8) · (2.1, 2.2) · (3.1, 3.2, 3.3, 3.4) · (4.1, 4.2, 4.3) · (5.1) · (6.1, 6.2, 6.3, 6.4)
Kvα7-12 (7.1, 7.2, 7.3, 7.4, 7.5) · (8.1, 8.2) · (9.1, 9.2, 9.3) · (10.1, 10.2) · (11.1/hERG, 11.2, 11.3) · (12.1, 12.2, 12.3)
Kvβ (1, 2, 3) · KCNIP (1, 2, 3, 4) · minK/ISK · minK/ISK-like · MiRP (1, 2, 3) · Shaker geneOther HVCN1GeneralCategories:- Ion channels
- Electrophysiology
- Integral membrane proteins

Wikimedia Foundation. 2010.