- Haematopoiesis
-
 Diagram showing the development of different blood cells from haematopoietic stem cell to mature cells
Diagram showing the development of different blood cells from haematopoietic stem cell to mature cells
Haematopoiesis (from Ancient Greek: αἷμα, "blood"; ποιεῖν "to make") (or hematopoiesis in the United States; sometimes also haemopoiesis or hemopoiesis) is the formation of blood cellular components. All cellular blood components are derived from haematopoietic stem cells. In a healthy adult person, approximately 1011–1012 new blood cells are produced daily in order to maintain steady state levels in the peripheral circulation.[1][2]
Contents
Haematopoietic stem cells (HSCs)
Haematopoietic stem cells (HSCs) reside in the medulla of the bone (bone marrow) and have the unique ability to give rise to all of the different mature blood cell types. HSCs are self renewing: when they proliferate, at least some of their daughter cells remain as HSCs, so the pool of stem cells does not become depleted. The other daughters of HSCs (myeloid and lymphoid progenitor cells), however can each commit to any of the alternative differentiation pathways that lead to the production of one or more specific types of blood cells, but cannot self-renew. This is one of the vital processes in the body.
Lineages
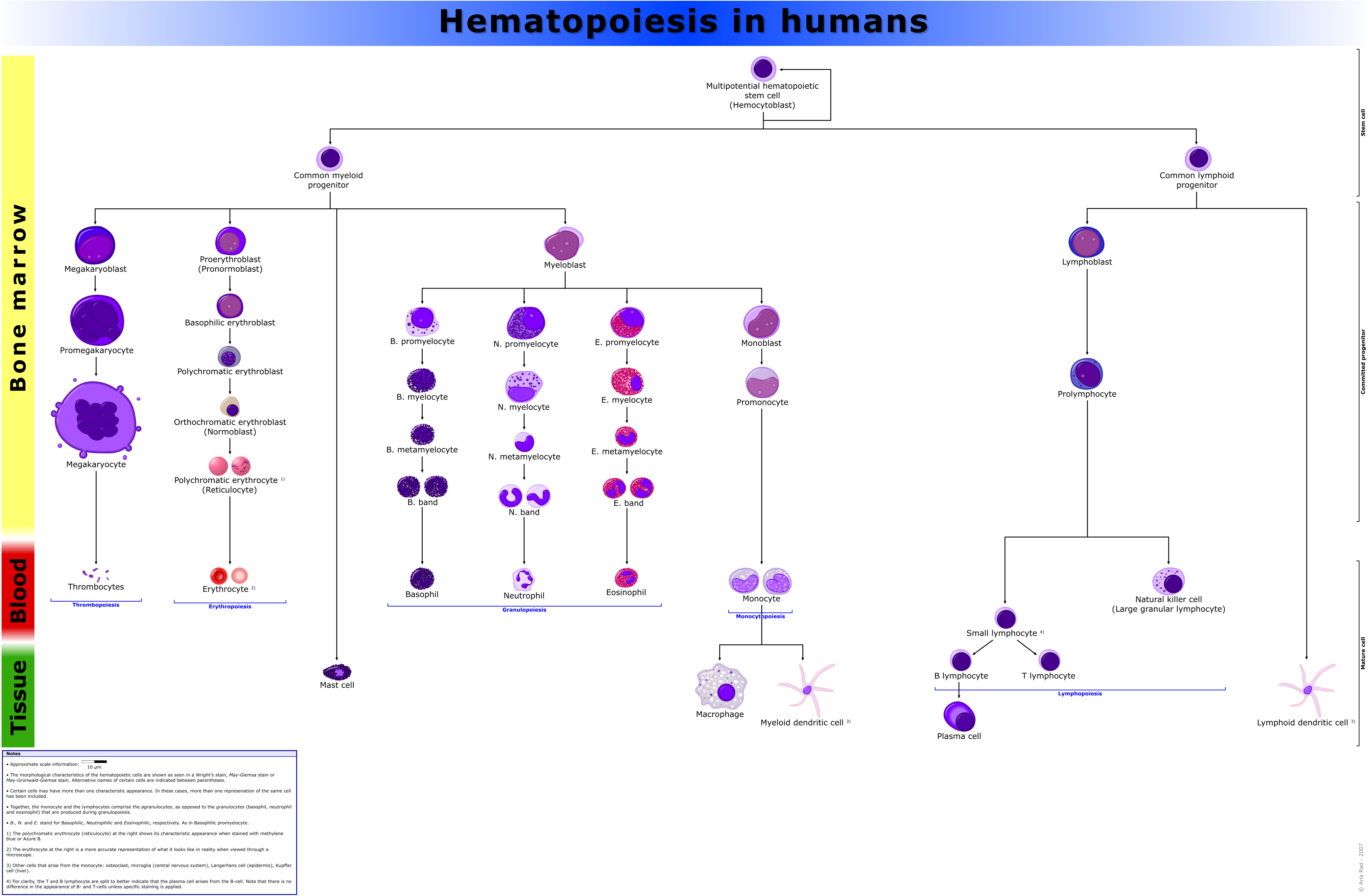
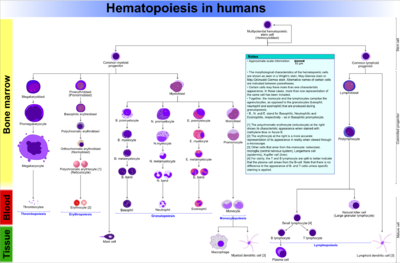 Comprehensive diagram that shows the development of different blood cells from haematopoietic stem cell to mature cells
Comprehensive diagram that shows the development of different blood cells from haematopoietic stem cell to mature cellsAll blood cells are divided into three lineages.
- Erythroid cells are the oxygen carrying red blood cells. Both reticulocytes and erythrocytes are functional and are released into the blood. In fact, a reticulocyte count estimates the rate of erythropoiesis.
- Lymphocytes are the cornerstone of the adaptive immune system. They are derived from common lymphoid progenitors. The lymphoid lineage is primarily composed of T-cells and B-cells (types of white blood cells). This is lymphopoiesis.
- Myelocytes, which include granulocytes, megakaryocytes and macrophages and are derived from common myeloid progenitors, are involved in such diverse roles as innate immunity, adaptive immunity, and blood clotting. This is myelopoiesis.
Granulopoiesis (or granulocytopoiesis) is haematopoiesis of granulocytes.
Megakaryocytopoiesis is haematopoiesis of megakaryocytes.
Locations
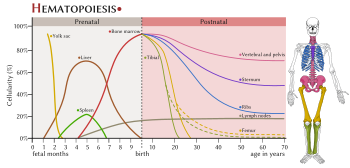 Sites of haematopoesis (human) in pre- and postnatal periods
Sites of haematopoesis (human) in pre- and postnatal periodsIn developing embryos, blood formation occurs in aggregates of blood cells in the yolk sac, called blood islands. As development progresses, blood formation occurs in the spleen, liver and lymph nodes. When bone marrow develops, it eventually assumes the task of forming most of the blood cells for the entire organism. However, maturation, activation, and some proliferation of lymphoid cells occurs in secondary lymphoid organs (spleen, thymus, and lymph nodes). In children, haematopoiesis occurs in the marrow of the long bones such as the femur and tibia. In adults, it occurs mainly in the pelvis, cranium, vertebrae, and sternum.
Extramedullary
In some cases, the liver, thymus, and spleen may resume their haematopoietic function, if necessary. This is called extramedullary haematopoiesis. It may cause these organs to increase in size substantially. During fetal development, since bones and thus the bone marrow, develop later, the liver functions as the main haematopoetic organ. Therefore, the liver is enlarged during development.
Other vertebrates
In some vertebrates, haematopoiesis can occur wherever there is a loose stroma of connective tissue and slow blood supply, such as the gut, spleen, kidney or ovaries.
Maturation
As a stem cell matures it undergoes changes in gene expression that limit the cell types that it can become and moves it closer to a specific cell type. These changes can often be tracked by monitoring the presence of proteins on the surface of the cell. Each successive change moves the cell closer to the final cell type and further limits its potential to become a different cell type.
Determination
Cell determination appears to be dictated by the location of differentiation.[citation needed] For instance, the thymus provides an ideal environment for thymocytes to differentiate into a variety of different functional T cells. For the stem cells and other undifferentiated blood cells in the bone marrow, the determination is generally explained by the determinism theory of haematopoiesis, saying that colony stimulating factors and other factors of the haematopoietic microenvironment determine the cells to follow a certain path of cell differentiation. This is the classical way of describing haematopoiesis. In fact, however, it is not really true. The ability of the bone marrow to regulate the quantity of different cell types to be produced is more accurately explained by a stochastic theory: Undifferentiated blood cells are determined to specific cell types by randomness. The haematopoietic microenvironment prevails upon some of the cells to survive and some, on the other hand, to perform apoptosis and die. By regulating this balance between different cell types, the bone marrow can alter the quantity of different cells to ultimately be produced.
Haematopoietic growth factors
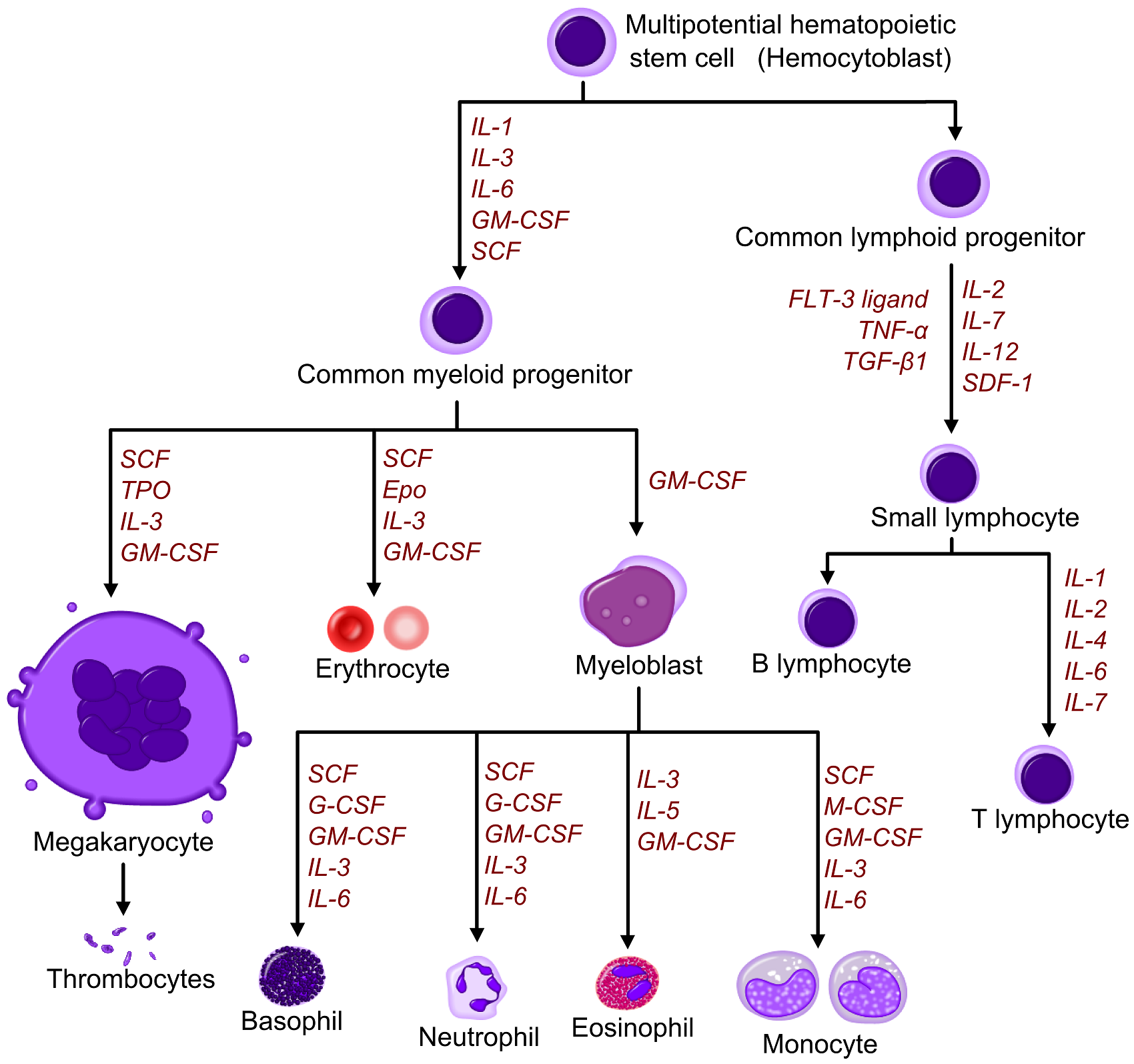
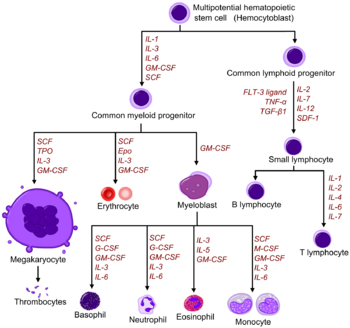 Diagram including some of the important cytokines that determine which type of blood cell will be created.[3] SCF= Stem Cell Factor Tpo= Thrombopoietin IL= Interleukin GM-CSF= Granulocyte Macrophage-colony stimulating factor Epo= Erythropoietin M-CSF= Macrophage-colony stimulating factor G-CSF= Granulocyte-colony stimulating factor SDF-1= Stromal cell-derived factor-1 FLT-3 ligand= FMS-like tyrosine kinase 3 ligand TNF-a = Tumour necrosis factor-alpha TGFβ = Transforming growth factor beta [4]
Diagram including some of the important cytokines that determine which type of blood cell will be created.[3] SCF= Stem Cell Factor Tpo= Thrombopoietin IL= Interleukin GM-CSF= Granulocyte Macrophage-colony stimulating factor Epo= Erythropoietin M-CSF= Macrophage-colony stimulating factor G-CSF= Granulocyte-colony stimulating factor SDF-1= Stromal cell-derived factor-1 FLT-3 ligand= FMS-like tyrosine kinase 3 ligand TNF-a = Tumour necrosis factor-alpha TGFβ = Transforming growth factor beta [4]Red and white blood cell production is regulated with great precision in healthy humans, and the production of granulocytes is rapidly increased during infection. The proliferation and self-renewal of these cells depend on stem cell factor (SCF). Glycoprotein growth factors regulate the proliferation and maturation of the cells that enter the blood from the marrow, and cause cells in one or more committed cell lines to proliferate and mature. Three more factors that stimulate the production of committed stem cells are called colony-stimulating factors (CSFs) and include granulocyte-macrophage CSF (GM-CSF), granulocyte CSF (G-CSF) and macrophage CSF (M-CSF). These stimulate much granulocyte formation and are active on either progenitor cells or end product cells.
Erythropoietin is required for a myeloid progenitor cell to become an erythrocyte.[3] On the other hand, thrombopoietin makes myeloid progenitor cells differentiate to megakaryocytes (thrombocyte-forming cells).[3] Examples of cytokines and the blood cells they give rise to, is shown in the picture to the right.
Transcription factors
Growth factors initiate signal transduction pathways, altering transcription factors, that, in turn activate genes that determine the differentiation of blood cells.
The early committed progenitors express low levels of transcription factors that may commit them to discrete cell lineages. Which cell lineage is selected for differentiation may depend both on chance and on the external signals received by progenitor cells. Several transcription factors have been isolated that regulate differentiation along the major cell lineages. For instance, PU.1 commits cells to the myeloid lineage whereas GATA-1 has an essential role in erythropoietic and megakaryocytic differentiation. The Ikaros, Aiolos and Helios transcription factors play a major role in lymphoid development.[5]
The myeloid-based model
For a decade now, the evidence is growing that HSC maturation follows a myeloid-based model instead of the 'classical' schoolbook dichotomy model. In the latter model, the HSC first generates a common myeloid-erythroid progenitor (CMEP) and a common lymphoid progenitor (CLP). The CLP produces only T or B cells. The myeloid-based model postulates that HSCs first diverge into the CMEP and a common myelo-lymphoid progenitor (CMLP), which generates T and B cell progenitors through a bipotential myeloid-T progenitor and a myeloid-B progenitor stage. The main difference is that in this new model, all erythroid, T and B lineage branches retain the potential to generate myeloid cells (even after the segregation of T and B cell lineages). The model proposes the idea of erythroid, T and B cells as specialized types of a prototypic myeloid HSC. Read more in Kawamoto et al. 2010.[6]
See also
- Haematon
- Haematopoietic stimulants:
- Erythropoiesis-stimulating agents
References
- ^ Semester 4 medical lectures at Uppsala University 2008 by Leif Jansson
- ^ Parslow,T G.;Stites, DP.; Terr, AI.; and Imboden JB.. Medical Immunology (1 ed.). ISBN 0838562787.
- ^ a b c Molecular cell biology. Lodish, Harvey F. 5. ed. : - New York : W. H. Freeman and Co., 2003, 973 s. b ill. ISBN 0-7167-4366-3
- ^
- For the growth factors also mentioned in previous version File:Hematopoiesis (human) cytokines.jpg: Molecular cell biology. Lodish, Harvey F. 5. ed. : - New York : W. H. Freeman and Co., 2003, 973 s. b ill. ISBN 0-7167-4366-3
- The rest: Rod Flower; Humphrey P. Rang; Maureen M. Dale; Ritter, James M. (2007). Rang & Dale's pharmacology. Edinburgh: Churchill Livingstone. ISBN 0-443-06911-5.
- ^ Rebollo, A.; C. Schmitt (2003). "Ikaros, Aiolos and Helios: Transcription regulators and lymphoid malignancies". Immunology and Cell Biology 81 (3): 171–175. doi:10.1046/j.1440-1711.2003.01159.x. PMID 12752680.
- ^ Kawamoto, Wada, Katsura. A revised scheme for developmental pathways of haematopoietic cells: the myeloid-based model. International Immunology 2010.
Further reading
- Godin, Isabelle & Cumano, Ana, ed (2006). Hematopoietic stem cell development. Springer. ISBN 9780306478727. http://books.google.com/books?id=tUsSmZwW_9MC.
External links
Immunology: Lymphocytic adaptive immune system and complement Lymphoid AntigensAntibodiesImmunity vs.
toleranceaction: Immunity · Autoimmunity · Alloimmunity · Allergy · Hypersensitivity · Inflammation · Cross-reactivity
inaction: Tolerance (Central, Peripheral, Clonal anergy, Clonal deletion, Tolerance in pregnancy) · ImmunodeficiencyLymphocytes Substances Complement Lymphoid system (TA A13.1–2, TH H3.10, GA 8 and 9) Primary lymphoid organs Secondary lymphoid organs structural: Hilum · Trabeculae · Diaphragmatic surface of spleen · Visceral surface of spleen
Red pulp (Cords of Billroth, Marginal zone)
White pulp (Periarteriolar lymphoid sheaths, Germinal center)
blood flow: Trabecular arteries · Trabecular veinslymph flow: Afferent lymph vessels · Cortical sinuses · Medullary sinuses · Efferent lymph vessels
T cells: High endothelial venules
B cells: Primary follicle/Germinal center · Mantle zone · Marginal zone
layers: Capsule/Trabeculae · Subcapsular sinus · Cortex · Paracortex · Medulla (Medullary cord) · HilumMALT
(process mucosa)M: LMO
anat(h, u, t, a, l)/phys/depv
noco/cong/tumr
proc
Myeloid physiology Hematopoiesis CFU-GMGranulopoiesis (Myeloblast, Promyelocyte, Myelocyte, Metamyelocyte, Band cell)
Monocytopoiesis (Monoblast, Promonocyte)Thrombopoiesis (Megakaryoblast, Promegakaryocyte)
Erythropoiesis (Proerythroblast, Normoblast, Reticulocyte)GeneralExtramedullary hematopoiesisHemostasis Other Erythrocyte aggregationCell signaling: cytokines By family CCLCXCLCX3CLXCLIL6 like/gp130IL-12 family/IL12RB1OtherIL-10 familyIL-17 familyOtherBy function/
cellCategories:- Hematopoiesis
- Histology


Wikimedia Foundation. 2010.