- Glucose
-
This article is about the naturally-occurring D-form of glucose. For the L-form, see L-Glucose.
D-glucose 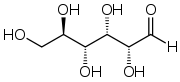
 D-glucoseSystematic name(2R,3S,4R,5R)-2,3,4,5,6-PentahydroxyhexanalOther namesBlood sugar
D-glucoseSystematic name(2R,3S,4R,5R)-2,3,4,5,6-PentahydroxyhexanalOther namesBlood sugar
Dextrose
Corn sugar
D-Glucose
Grape sugarIdentifiers Abbreviations Glc CAS number 50-99-7 
PubChem 5793 ChemSpider 5589 UNII 5SL0G7R0OK EC number 200-075-1 KEGG C00031 
MeSH Glucose ChEBI CHEBI:4167 ChEMBL CHEMBL1222250 RTECS number LZ6600000 ATC code B05,V04CA02, V06DC01 Beilstein Reference 1281604 Gmelin Reference 83256 3DMet B04623 Jmol-3D images Image 1
Image 2- OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O
C([C@@H]1[C@H]([C@@H]([C@H]([C@H](O1)O)O)O)O)O
- InChI=1S/C12H24O12/c7-1-2-3(8)4(9)5(10)6(11)12-2/h2-11H,1H2/t2-,3-,4+,5-,6?/m1/s1
Key: WQZGKKKJIJFFOK-GASJEMHNSA-N
InChI=1/C12H24O12/c7-1-2-3(8)4(9)5(10)6(11)12-2/h2-11H,1H2/t2-,3-,4+,5-,6+/m1/s1
Key: WQZGKKKJIJFFOK-DVKNGEFBBQ
Properties Molecular formula C6H12O6 Molar mass 180.16 g/mol Exact mass 180.063388 Density 1.54 g/cm3 Melting point α-D-glucose: 146 °C
β-D-glucose: 150 °CSolubility in water 91 g/100 mL Thermochemistry Std enthalpy of
formation ΔfHo298−1271 kJ/mol Std enthalpy of
combustion ΔcHo298−2805 kJ/mol Standard molar
entropy So298209.2 J K−1 mol−1 Hazards MSDS ICSC 0865 EU Index not listed  (verify) (what is:
(verify) (what is:  /
/ ?)
?)
Except where noted otherwise, data are given for materials in their standard state (at 25 °C, 100 kPa)Infobox references Glucose (pronounced /ˈɡluːkoʊs, -koʊz/; C6H12O6, also known as D-glucose, dextrose, or grape sugar) is a simple sugar (monosaccharide) and an important carbohydrate in biology. Cells use it as the primary source of energy[1] and a metabolic intermediate. Glucose is one of the main products of photosynthesis and starts cellular respiration.
Glucose exists in several different structures, but all of these structures can be divided into two families of mirror-images (stereoisomers). Only one set of these isomers exists in nature, those derived from the "right-handed form" of glucose, denoted D-glucose. D-glucose is often referred to as dextrose. The term dextrose is derived from dextrorotatory glucose.[2] Solutions of dextrose rotate polarized light to the right. Starch and cellulose are polymers derived from the dehydration of D-glucose. The other stereoisomer, called L-glucose, is hardly found in nature.
The name "glucose" comes from the Greek word glukus (γλυκύς), meaning "sweet". The suffix "-ose" denotes a sugar. The name "dextrose" and the 'D-' prefix come from Latin dexter ("right"), referring to the handedness of the molecules.
Contents
Function
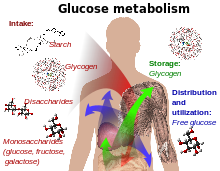 Glucose metabolism and various forms of it in the process.
Glucose metabolism and various forms of it in the process.
-Glucose-containing compounds and isomeric forms are digested and taken up by the body in the intestines, including starch, glycogen, disaccharides and monosaccharides.
-Glucose is stored in mainly the liver and muscles as glycogen.
-It is distributed and utilized in tissues as free glucose.Scientists can speculate on the reasons that glucose, and not another monosaccharide such as fructose, is so widely used in organisms. One reason might be that glucose has a lower tendency, relative to other hexose sugars, to react non-specifically with the amino groups of proteins. This reaction (glycation) reduces or destroys the function of many enzymes. The low rate of glycation is due to glucose's preference for the less reactive cyclic isomer. Nevertheless, many of the long-term complications of diabetes (e.g., blindness, renal failure, and peripheral neuropathy) are probably due to the glycation of proteins or lipids.[3] In contrast, enzyme-regulated addition of glucose to proteins by glycosylation is often essential to their function.[citation needed]
Analyte in medical blood test
Main article: Glucose testGlucose is a common medical analyte measured in blood samples. Eating or fasting prior to taking a blood sample has an effect on the result. Higher than usual glucose levels may be a sign of prediabetes or diabetes mellitus.
As an energy source
Glucose is a ubiquitous fuel in biology. It is used as an energy source in most organisms, from bacteria to humans. Use of glucose may be by either aerobic respiration, anaerobic respiration, or fermentation. Glucose is the human body's key source of energy, through aerobic respiration, providing approximately 3.75 kilocalories (16 kilojoules) of food energy per gram.[4] Breakdown of carbohydrates (e.g. starch) yields mono- and disaccharides, most of which is glucose. Through glycolysis and later in the reactions of the citric acid cycle (TCAC), glucose is oxidized to eventually form CO2 and water, yielding energy sources, mostly in the form of ATP. The insulin reaction, and other mechanisms, regulate the concentration of glucose in the blood. A high fasting blood sugar level is an indication of prediabetic and diabetic conditions.
Glucose is a primary source of energy for the brain, and hence its availability influences psychological processes. When glucose is low, psychological processes requiring mental effort (e.g., self-control, effortful decision-making) are impaired.[5][6][7][8]
Glucose in glycolysis
α-D-Glucose Hexokinase α-D-Glucose-6-phosphate 

ATP ADP 
Compound C00031 at KEGG Pathway Database. Enzyme 2.7.1.1 at KEGG Pathway Database. Compound C00668 at KEGG Pathway Database. Reaction R01786 at KEGG Pathway Database. Use of glucose as an energy source in cells is via aerobic or anaerobic respiration. Both of these start with the early steps of the glycolysis metabolic pathway. The first step of this is the phosphorylation of glucose by hexokinase to prepare it for later breakdown to provide energy. The major reason for the immediate phosphorylation of glucose by a hexokinase is to prevent diffusion out of the cell. The phosphorylation adds a charged phosphate group so the glucose 6-phosphate cannot easily cross the cell membrane. Irreversible first steps of a metabolic pathway are common for regulatory purposes.
In anaerobic respiration one glucose molecule produces a net gain of 2 ATP molecules (4 ATP molecules are produced during glycolysis but 2 are required by enzymes used during the process).[9] In aerobic respiration a molecule of glucose is much more profitable in that a net worth of 32 ATP molecules are generated (34 gross with 2 being required in the process).[10]
As a precursor
Glucose is critical in the production of proteins and in lipid metabolism. In plants and most animals, it is also a precursor for vitamin C (ascorbic acid) production. It is modified for use in these processes by the glycolysis pathway.
Glucose is used as a precursor for the synthesis of several important substances. Starch, cellulose, and glycogen ("animal starch") are common glucose polymers (polysaccharides). Lactose, the predominant sugar in milk, is a glucose-galactose disaccharide. In sucrose, another disaccharide, glucose is joined to fructose. These synthesis processes also rely on the phosphorylation of glucose through the first step of glycolysis.
Industrial use
In industry, glucose is used as a precursor to make vitamin C in the Reichstein process, to make citric acid, gluconic acid, bio-ethanol, polylactic acid, sorbitol.
Structure and nomenclature
Glucose is a monosaccharide with formula C6H12O6 or H-(C=O)-(CHOH)5-H, whose five hydroxyl (OH) groups are arranged in a specific way along its six-carbon backbone.
Open-chain form
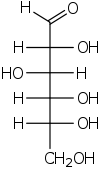 D-glucose in Fischer projection
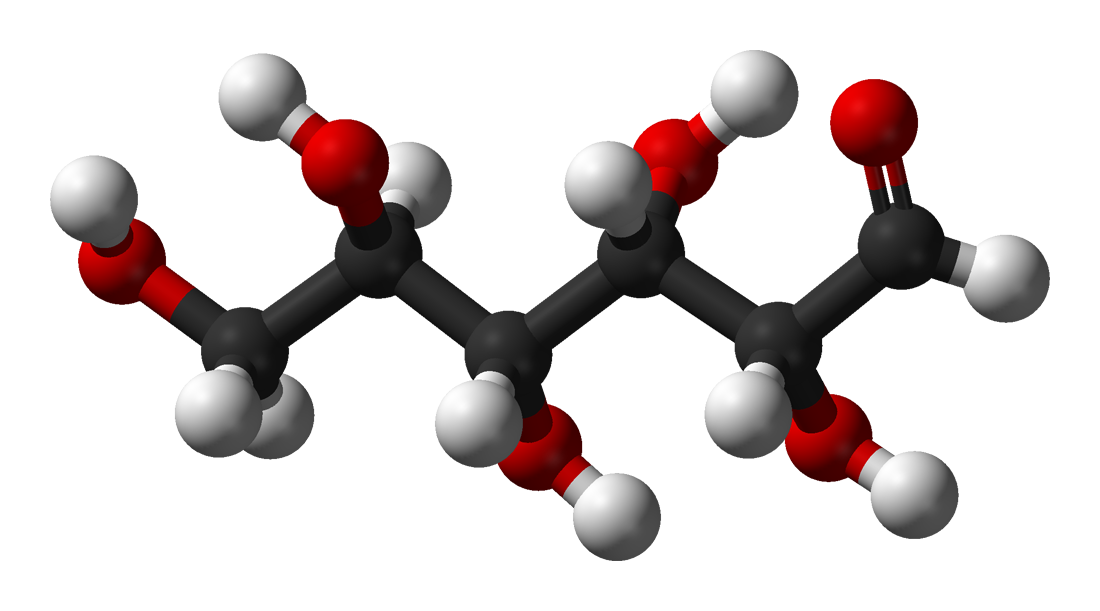
D-glucose in Fischer projectionIn its fleeting open-chain form, the glucose molecule has an open (as opposed to cyclic) and unbranched backbone of six carbon atoms, C-1 through C-6; where C-1 is part of an aldehyde group H(C=O)-, and each of the other five carbons bears one hydroxyl group -OH. The remaining bonds of the backbone carbons are satisfied by hydrogen atoms -H. Therefore glucose is an hexose and an aldose, or an aldohexose.
Each of the four carbons C-2 through C-5 is chiral, meaning that its four bonds connect to four different substituents. (Carbon C-2, for example, connects to -(C=O)H, -OH, -H, and -(CHOH)4H.) In D-glucose, these four parts must be in a specific three-dimensional arrangement. Namely, when the molecule is drawn in the Fischer projection, the hydroxyls on C-2, C-4, and C-5 must be on the right side, while that on C-3 must be on the left side.
The positions of those four hydroxyls are exactly reversed in the Fischer diagram of L-Glucose. D- and L-glucose are two of the 16 possible aldohexoses; the other 14 are allose, altrose, mannose, gulose, idose, galactose, and talose, each with two isomers, 'D-' and 'L-'.
Cyclic forms
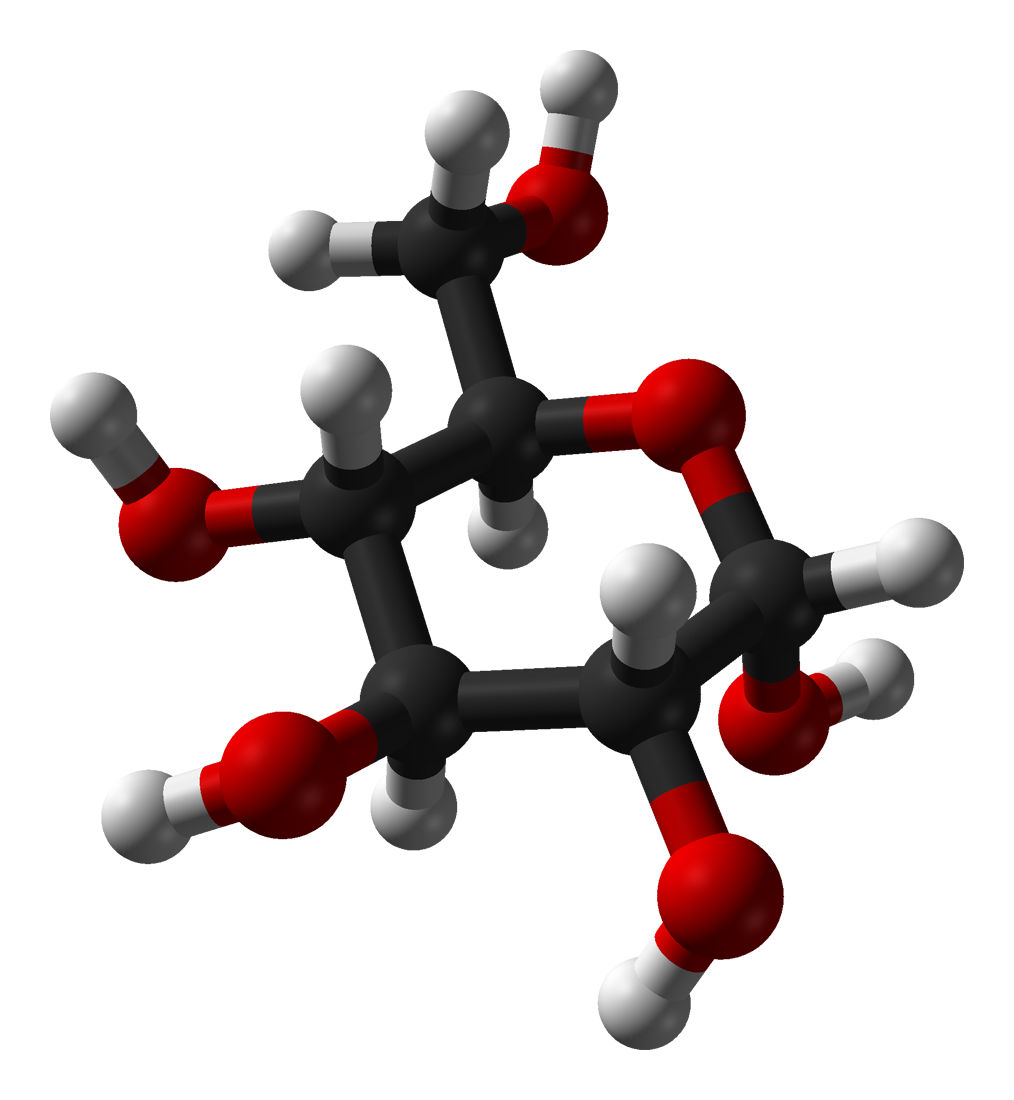
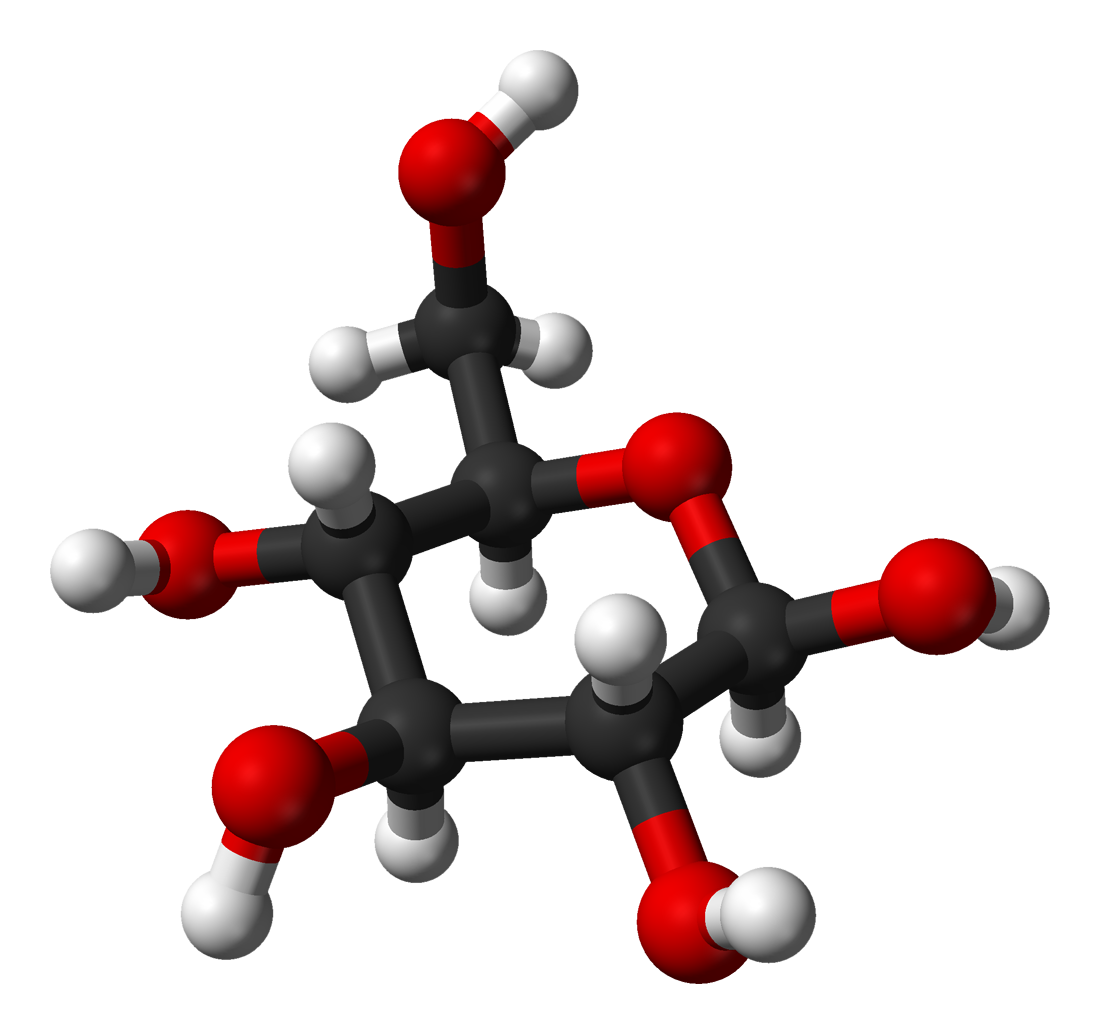
In solutions, the open-chain form of glucose (either 'D-' or 'L-') exists in equilibrium with several cyclic isomers, each containing a ring of carbons closed by one oxygen atom. In aqueous solution, however, glucose exists as pyranose for more than 99%. The open-chain form is limited to about 0.25% and furanose exists in negligible amounts. The terms "glucose" and "D-glucose" are generally used for these cyclic forms as well. The ring arises from the open-chain form by a nucleophilic addition reaction between the aldehyde group -(C=O)H at C-1 and the hydroxyl group -OH at C-4 or C-5, yielding a hemiacetal group -C(OH)H-O-.
The reaction between C-1 and C-5 creates a molecule with a six-membered ring, called pyranose, after the cyclic ether pyran, the simplest molecule with the same carbon-oxygen ring. The (much rarer) reaction between C-1 and C-4 creates a molecule with a five-membered ring, called furanose, after the cyclic ether furan. In either case, each carbon in the ring has one hydrogen and one hydroxyl attached, except for the last carbon (C-4 or C-5) where the hydroxyl is replaced by the remainder of the open molecule (which is -(CHOH)2-H or -(CHOH)-H, respectively).
The ring-closing reaction makes carbon C-1 chiral, too, since its four bonds lead to -H, to -OH, to carbon C-2, and to the ring oxygen. These four parts of the molecule may be arranged around C-1 (the anomeric carbon) in two distinct ways, designated by the prefixes 'α-' and 'β-'. When a glucopyranose molecule is drawn in the Haworth projection, the designation 'α-' means that the hydroxyl group attached to C-1 and the -CH2OH group at C-5 lies on opposite sides of the ring's plane (a trans arrangement), while 'β-' means that they are on the same side of the plane (a cis arrangement).
Therefore, the open isomer D-glucose gives rise to four distinct cyclic isomers: α-D-glucopyranose, β-D-glucopyranose, α-D-glucofuranose, and β-D-glucofuranose; which are all chiral.
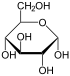
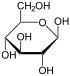
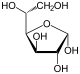
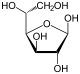
α-D-
Glucopyranoseβ-D-
Glucopyranoseα-D-
Glucofuranoseβ-D-
Glucofuranose-
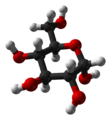
α-D-
Glucopyranose -
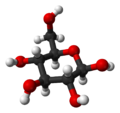
β-D-
Glucopyranose
The other open-chain isomer L-glucose similarly gives rise to four distinct cyclic forms of L-glucose, each the mirror image of the corresponding D-glucose.
The rings are not planar but twisted in three dimensions. The glucopyranose ring (α or β) can assume several non-planar shapes, analogous to the 'chair' and 'boat' conformations of cyclohexane. Similarly, the glucofuranose ring may assume several shapes, analogous to the 'envelope' conformations of cyclopentane.
The glucopyranose forms of glucose predominate in solution, and are the only forms observed in the solid state. They are crystalline colorless solids, highly soluble in water and acetic acid, poorly soluble in methanol and ethanol. They melt at 146 °C (295 °F) (α) and 150 °C (302 °F) (β), and decompose at higher temperatures into carbon and water.
Rotational isomers
Each glucose isomer is subject to rotational isomerism. Within the cyclic form of glucose, rotation may occur around the O6-C6-C5-O5 torsion angle, termed the ω-angle, to form three staggered rotamer conformations called gauche-gauche (gg), gauche-trans (gt) and trans-gauche (tg). For methyl α-D-glucopyranose at equilibrium the ratio of molecules in each rotamer conformation is reported as 57:38:5 gg:gt:tg.[11] This tendency for the ω-angle to prefer to adopt a gauche conformation is attributed to the gauche effect.
Physical properties
Solutions
All forms of glucose are colorless and easily soluble in water, acetic acid, and several other solvents. They are only sparingly soluble in methanol and ethanol.
The open-chain form is thermodynamically unstable, and it spontaneously tautomerizes to the cyclic forms. (Although the ring closure reaction could in theory create four- or three-atom rings, these would be highly strained and are not observed.) In solutions at room temperature, the four cyclic isomers interconvert over a timescale of hours, in a process called mutarotation.[12] Starting from any proportions, the mixture converges stable ratio of α:β 36:64. The ratio would be α:β 11:89 if it were not for the influence of the anomeric effect.[13] Mutarotation is considerably slower at temperatures close to 0°C.
Mutarotation consists of a temporary reversal of the ring-forming reaction, resulting in the open-chain form, followed by a re-forming of the ring. The ring closure step may use a different -OH group than the one recreated by the opening step (thus switching between pyranose and furanose forms), and/or the new hemiacetal group created on C-1 may have the same or opposite handedness as the original one (thus switching between the α and β forms). Thus, even though the open-chain form is barely detectable in solution, it is an essential component of the equilibrium.
Solid state
Depending on conditions, three major solid forms of glucose can be crystallised from water solutions: α-glucopyranose, β-glucopyranose, and β-glucopyranose hydrate.[14]
Optical activity
Whether in water or in the solid form, D-glucose is dextrorotatory, meaning that it will rotate the direction of polarized light clockwise. The effect is due to the chirality of the molecules, and indeed the mirror-image isomer, L-glucose, is levorotatory (rotates polarized light counterclockwise) by the same amount. The strength of the effect is different for each of the five tautomers.
Note that the D- prefix does not refer directly to the optical properties of the compound. It indicates that the C-2 chiral center has the same handedness as that of D-glutaraldehyde (which was so labeled because it is dextrorotatory). The fact that D-glucose is dextrorotatory is a combined effect of its four chiral centers, not just of C-2; and indeed some of the other D-aldohexoses are levorotatory.
Production
 Glucose tablets
Glucose tabletsBiosynthesis
In plants and some prokaryotes, glucose is a product of photosynthesis. In animals and fungi, glucose results from the breakdown of glycogen, a process known as glycogenolysis. In plants the breakdown substrate is starch.
In animals, glucose is synthesized in the liver and kidneys from non-carbohydrate intermediates, such as pyruvate and glycerol, by a process known as gluconeogenesis.
In some deep-sea bacteria glucose is produced by chemosynthesis.
Commercial
Glucose is produced commercially via the enzymatic hydrolysis of starch. Many crops can be used as the source of starch. Maize, rice, wheat, cassava, corn husk and sago are all used in various parts of the world. In the United States, cornstarch (from maize) is used almost exclusively. Most commercial glucose occurs as a component of invert sugar, an approximately 1:1 mixture of glucose and fructose. In principle, cellulose could be hydrolysed to glucose, but this process is not yet commercially practical.[14]
Sources and absorption
Most dietary carbohydrates contain glucose, either as their only building block, as in starch and glycogen, or together with another monosaccharide, as in sucrose and lactose.
In the lumen of the duodenum and small intestine, the glucose oligo- and polysaccharides are broken down to monosaccharides by the pancreatic and intestinal glycosidases. Other polysaccharides cannot be processed by the human intestine and require assistance by intestinal flora if they are to be broken down; the most notable exceptions are sucrose (fructose-glucose) and lactose (galactose-glucose). Glucose is then transported across the apical membrane of the enterocytes by SLC5A1, and later across their basal membrane by SLC2A2.[15] Some of the glucose is directly utilized as an energy source by brain cells, intestinal cells and red blood cells, while the rest reaches the liver, adipose tissue and muscle cells, where it is absorbed and stored as glycogen (under the influence of insulin). Liver cell glycogen can be converted to glucose and returned to the blood when insulin is low or absent; muscle cell glycogen is not returned to the blood because of a lack of enzymes. In fat cells, glucose is used to power reactions that synthesize some fat types and have other purposes. Glycogen is the body's 'glucose energy storage' mechanism because it is much more 'space efficient' and less reactive than glucose itself.
History
Because glucose is a basic necessity of many organisms, a correct understanding of its chemical makeup and structure contributed greatly to a general advancement in organic chemistry. This understanding occurred largely as a result of the investigations of Emil Fischer, a German chemist who received the 1902 Nobel Prize in Chemistry as a result of his findings.[16] The synthesis of glucose established the structure of organic material and consequently formed the first definitive validation of Jacobus Henricus van't Hoff's theories of chemical kinetics and the arrangements of chemical bonds in carbon-bearing molecules.[17] Between 1891 and 1894, Fischer established the stereochemical configuration of all the known sugars and correctly predicted the possible isomers, applying van't Hoff's theory of asymmetrical carbon atoms.
See also
- HbA1c
- DMF (potential glucose-based biofuel)
- Beriberi - vitamin deficiency affecting ability to convert carbohydrates into glucose
- Fructose
- Galactose
- Lactose
- Maltose
- Sucrose
- Sugar
- Sugars in wine
- Trinder glucose activity test
- Glucose transporter (GLUT): GLUT1, GLUT2
- Caramelization
- Peritoneal dialysis
- Fludeoxyglucose_(18F)
References
- ^ Clark, D.; Sokoloff, L. (1999), Basic Neurochemistry: Molecular, Cellular and Medical Aspects, Lippincott, pp. 637–670
- ^ "dextrose", Merriam-Webster Online Dictionary, http://www.m-w.com/dictionary/dextrose, retrieved 2009-09-02.
- ^ High Blood Glucose and Diabetes Complications: The buildup of molecules known as AGEs may be the key link, American Diabetes Association, 2010, ISSN 0095-8301, http://forecast.diabetes.org/magazine/features/high-blood-glucose-and-diabetes-complications
- ^ "Chapter 3: Calculation of the Energy Content of Foods – Energy Conversion Factors", Food energy - methods of analysis and conversion factors, FAO Food and Nutrition Paper 77, Rome: Food and Agriculture Organization, 2003, ISBN 92-5-105014-7, http://www.fao.org/docrep/006/Y5022E/y5022e04.htm.
- ^ Fairclough, Stephen H.; Houston, Kim (2004), "A metabolic measure of mental effort", Biol. Psychol. 66 (2): 177–90, doi:10.1016/j.biopsycho.2003.10.001, PMID 15041139.
- ^ Gailliot, Matthew T.; Baumeister, Roy F.; DeWall, C. Nathan; Plant, E. Ashby; Brewer, Lauren E.; Schmeichel, Brandon J.; Tice, Dianne M.; Maner, Jon K. (2007), "Self-Control Relies on Glucose as a Limited Energy Source: Willpower is More than a Metaphor", J. Personal. Soc. Psychol. 92 (2): 325–36, doi:10.1037/0022-3514.92.2.325, PMID 17279852.
- ^ Gailliot, Matthew T.; Baumeister, Roy F. (2007), "The Physiology of Willpower: Linking Blood Glucose to Self-Control", Personal. Soc. Psychol. Rev. 11 (4): 303–27, doi:10.1177/1088868307303030, PMID 18453466.
- ^ Masicampo, E. J.; Baumeister, Roy F. (2008), "Toward a Physiology of Dual-Process Reasoning and Judgment: Lemonade, Willpower, and Expensive Rule-Based Analysis", Psychol. Sci. 19 (3): 255–60, doi:10.1111/j.1467-9280.2008.02077.x, PMID 18315798.
- ^ Medical Biochemistry at a Glance @Google books, Blackwell Publishing, 2006, ISBN 978-1-4051-1322-9, http://books.google.co.uk/books?id=9BtxCWxrWRoC&pg=PA52
- ^ Medical Biochemistry at a Glance @Google books, Blackwell Publishing, 2006, ISBN 978-1-4051-1322-9, http://books.google.co.uk/books?id=9BtxCWxrWRoC&pg=PA50
- ^ Kirschner, Karl N.; Woods, Robert J. (2001), "Solvent interactions determine carbohydrate conformation", Proc. Natl. Acad. Sci. USA 98 (19): 10541–45, doi:10.1073/pnas.191362798, PMC 58501, PMID 11526221, http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=58501
- ^ McMurry, John E. (1988), Organic Chemistry (2nd ed.), Brooks/Cole, p. 866, ISBN 0534079687.
- ^ Juaristi, Eusebio; Cuevas, Gabriel (1995), The Anomeric Effect, CRC Press, pp. 9–10, ISBN 0849389410.
- ^ a b Fred W. Schenck “Glucose and Glucose-Containing Syrups” in Ullmann's Encyclopedia of Industrial Chemistry 2006, Wiley-VCH, Weinheim. doi: 10.1002/14356007.a12_457.pub2
- ^ Ferraris, Ronaldo P. (2001), "Dietary and developmental regulation of intestinal sugar transport", Biochem. J. 360 (Pt 2): 265–76, doi:10.1042/0264-6021:3600265, PMC 1222226, PMID 11716754, http://www.biochemj.org/bj/360/0265/bj3600265.htm.
- ^ Emil Fischer, Nobel Foundation, http://nobelprize.org/nobel_prizes/chemistry/laureates/1902/fischer-bio.html, retrieved 2009-09-02.
- ^ Fraser-Reid, Bert, "van't Hoff's Glucose", Chem. Eng. News 77 (39): 8.
External links
Glycolysis Metabolic Pathway Glucose Hexokinase Glucose 6-phosphate Glucose-6-phosphate isomerase Fructose 6-phosphate phosphofructokinase-1 Fructose 1,6-bisphosphate Fructose bisphosphate aldolase 
ATP ADP 
ATP ADP 

Dihydroxyacetone phosphate Glyceraldehyde 3-phosphate Triosephosphate isomerase Glyceraldehyde 3-phosphate Glyceraldehyde-3-phosphate dehydrogenase 1,3-Bisphosphoglycerate 
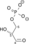
NAD+ + Pi NADH + H+ 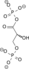
+ 2 
2 Phosphoglycerate kinase 3-Phosphoglycerate Phosphoglycerate mutase 2-Phosphoglycerate Phosphopyruvate hydratase(Enolase) Phosphoenolpyruvate Pyruvate kinase Pyruvate ADP ATP 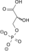
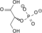
H2O 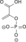
ADP ATP 
2 2 
2 2 Types of carbohydrates General: Geometry Monosaccharides Aldodiose (Glycolaldehyde)Ketopentose (Ribulose, Xylulose)
Aldopentose (Ribose, Arabinose, Xylose, Lyxose)
Deoxy sugar (Deoxyribose)Ketoheptose (Sedoheptulose, Mannoheptulose)>7Multiple Other oligosaccharidesGlucose/Glucan: Glycogen · Starch (Amylose, Amylopectin) · Cellulose · Dextrin/Dextran · Beta-glucan (Zymosan, Lentinan, Sizofiran) · Maltodextrin
Fructose/Fructan: Inulin · Levan beta 2→6
N-Acetylglucosamine: Chitinbiochemical families: prot · nucl · carb (glpr, alco, glys) · lipd (fata/i, phld, strd, gllp, eico) · amac/i · ncbs/i · ttpy/i Categories:- Glycolysis
- Chemical pathology
- Glucose
- Nutrition
- Sweeteners
- Starch
- World Health Organization essential medicines
- OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O








Wikimedia Foundation. 2010.