- Cellular respiration
-
 Cellular respiration in a typical eukaryotic cell.
Cellular respiration in a typical eukaryotic cell.
Cellular respiration is the set of the metabolic reactions and processes that take place in the cells of organisms to convert biochemical energy from nutrients into adenosine triphosphate (ATP), and then release waste products. The reactions involved in respiration are catabolic reactions that involve the redox reaction (oxidation of one molecule and the reduction of another). Respiration is one of the key ways a cell gains useful energy to fuel cellular reformations. (glucose+oxygen= carbon dioxide+water+energy
Nutrients commonly used by animal and plant cells in respiration include sugar, amino acids and fatty acids, and a common oxidizing agent (electron acceptor) is molecular oxygen (O2). Bacteria and archaea can also be lithotrophs and these organisms may respire using a broad range of inorganic molecules as electron donors and acceptors, such as sulfur, metal ions, methane or hydrogen. Organisms that use oxygen as a final electron acceptor in respiration are described as aerobic, while those that do not are referred to as anaerobic.[1]
The energy released in respiration is used to synthesize ATP to store this energy. The energy stored in ATP can then be used to drive processes requiring energy, including biosynthesis, locomotion or transportation of molecules across cell membranes.
Contents
Aerobic respiration
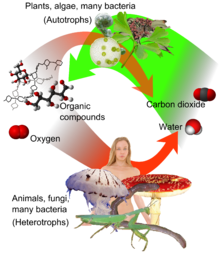 Aerobic respiration (red arrows) is the main means by which both plants and animals utilize energy in the form of organic compounds that were previously created through photosynthesis (green arrow).
Aerobic respiration (red arrows) is the main means by which both plants and animals utilize energy in the form of organic compounds that were previously created through photosynthesis (green arrow).Aerobic respiration requires oxygen in order to generate energy (ATP). Although carbohydrates, fats, and proteins can all be processed and consumed as reactant, it is the preferred method of pyruvate breakdown in glycolysis and requires that pyruvate enter the mitochondrion in order to be fully oxidized by the Krebs cycle. The product of this process is energy in the form of ATP (Adenosine triphosphate), by substrate-level phosphorylation, NADH and FADH2
Simplified reaction: C6H12O6 (aq) + 6 O2 (g) → 6 CO2 (g) + 6 H2O (l) ΔG = -2880 kJ per mole of C6H12O6 The negative ΔG indicates that the reaction can happen spontaneously
The reducing potential of NADH and FADH2 is converted to more ATP through an electron transport chain with oxygen as the "terminal electron acceptor". Most of the ATP produced by aerobic cellular respiration is made by oxidative phosphorylation. This works by the energy released in the consumption of pyruvate being used to create a chemiosmotic potential by pumping protons across a membrane. This potential is then used to drive ATP synthase and produce ATP from ADP and a phosphate group. Biology textbooks often state that 38 ATP molecules can be made per oxidised glucose molecule during cellular respiration (2 from glycolysis, 2 from the Krebs cycle, and about 34 from the electron transport system).[2] However, this maximum yield is never quite reached due to losses (leaky membranes) as well as the cost of moving pyruvate and ADP into the mitochondrial matrix and current estimates range around 29 to 30 ATP per glucose.[2]
Aerobic metabolism is 19 times more efficient than anaerobic metabolism (which yields 2 mol ATP per 1 mol glucose). They share the initial pathway of glycolysis but aerobic metabolism continues with the Krebs cycle and oxidative phosphorylation. The post glycolytic reactions take place in the mitochondria in eukaryotic cells, and in the cytoplasm in prokaryotic cells.
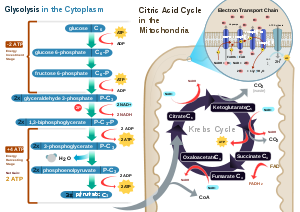
Glycolysis
Main article: GlycolysisGlycolysis is a metabolic pathway that is found in the cytosol of cells in all living organisms. This pathway does not require oxygen, and can therefore function under anaerobic circumstances. The process converts one molecule of glucose into two molecules of pyruvate (pyruvic acid), making energy in the form of two net molecules of ATP. Four molecules of ATP per glucose are actually produced; however, two are consumed for the preparatory phase. The initial phosphorylation of glucose is required to destabilize the molecule for cleavage into two pyruvate. During the pay-off phase of glycolysis, four phosphate groups are transferred to ADP by substrate-level phosphorylation to make four ATP, and two NADH are produced when the pyruvate are oxidized. The overall reaction can be expressed this way:
- Glucose + 2 NAD+ + 2 Pi + 2 ADP → 2 pyruvate + 2 NADH + 2 ATP + 2 H+ + 2 H2O
There can be at the most 38 ATP molecules formed
Oxidative decarboxylation of pyruvate
Main article: Pyruvate decarboxylationPyruvate is oxidized to acetyl-CoA and CO2 by the Pyruvate dehydrogenase complex (PDC). The PDC contains multiple copies of three enzymes and is located in the mitochondria of eukaryotic cells and in the cytosol of prokaryotes. In the conversion of pyruvate to acetyl-CoA one molecule of NADH and one molecule of CO2 is formed. This step is also known as the link reaction or transition step, as it links glycolysis and the Krebs cycle.
Citric acid cycle
Main article: Citric acid cycleThis is also called the Krebs cycle or the tricarboxylic acid cycle. When oxygen is present, acetyl-CoA is produced from the pyruvate molecules created from glycolysis. Once acetyl-CoA is formed, two processes can occur, aerobic or anaerobic respiration. When oxygen is present, the mitochondria will undergo aerobic respiration which leads to the Krebs cycle. However, if oxygen is not present, fermentation of the pyruvate molecule will occur. In the presence of oxygen, when acetyl-CoA is produced, the molecule then enters the citric acid cycle (Krebs cycle) inside the mitochondrial matrix, and gets oxidized to CO2 while at the same time reducing NAD to NADH. NADH can be used by the electron transport chain to create further ATP as part of oxidative phosphorylation. To fully oxidize the equivalent of one glucose molecule, two acetyl-CoA must be metabolized by the Krebs cycle. Two waste products, H2O and CO2, are created during this cycle.
The citric acid cycle is an 8-step process involving different enzymes and co-enzymes. Throughout the entire cycle, acetyl-CoA(2 carbons) + Oxaloacetate(4 carbons). Citrate(6 carbons) is rearranged to a more reactive form called Isocitrate(6 carbons). Isocitrate(6 carbons) modifies to become α-Ketoglutarate(5 carbons), Succinyl-CoA, Succinate, Fumarate, Malate, and finally, Oxaloacetate. The net energy gain from one cycle is 3 NADH, 1 FADH2, and 1 GTP; the GTP may subsequently be used to produce ATP. Thus, the total energy yield from one whole glucose molecule (2 pyruvate molecules) is 6 NADH, 2 FADH2, and 2 ATP.
Oxidative phosphorylation
Main articles: Oxidative phosphorylation, Electron transport chain, Electrochemical gradient, and ATP synthaseIn eukaryotes, oxidative phosphorylation occurs in the mitochondrial cristae. It comprises the electron transport chain that establishes a proton gradient (chemiosmotic potential) across the inner membrane by oxidizing the NADH produced from the Krebs cycle. ATP is synthesised by the ATP synthase enzyme when the chemiosmotic gradient is used to drive the phosphorylation of ADP. The electrons are finally transferred to exogenous oxygen and, with the addition of two protons, water is formed.
The table below describes the reactions involved when one glucose molecule is fully oxidized into carbon dioxide. It is assumed that all the reduced coenzymes are oxidized by the electron transport chain and used for oxidative phosphorylation.
Step coenzyme yield ATP yield Source of ATP Glycolysis preparatory phase -2 Phosphorylation of glucose and fructose 6-phosphate uses two ATP from the cytoplasm. Glycolysis pay-off phase 4 Substrate-level phosphorylation 2 NADH 4-6 Oxidative phosphorylation - Each NADH produces net 2 ATP due to NADH transport over the mitochondrial membrane Oxidative decarboxylation of pyruvate 2 NADH 6 Oxidative phosphorylation Krebs cycle 2 Substrate-level phosphorylation 6 NADH 18 Oxidative phosphorylation 2 FADH2 4 Oxidative phosphorylation Total yield 36-38 ATP From the complete oxidation of one glucose molecule to carbon dioxide and oxidation of all the reduced coenzymes. Although there is a theoretical yield of 38 ATP molecules per glucose during cellular respiration, such conditions are generally not realized due to losses such as the cost of moving pyruvate (from glycolysis), phosphate, and ADP (substrates for ATP synthesis) into the mitochondria. All are actively transported using carriers that utilise the stored energy in the proton electrochemical gradient.
- Pyruvate is taken up by a specific, low km transporter to bring it into the mitochondrial matrix for oxidation by the pyruvate dehydrogenase complex.
- The phosphate translocase is a symporter and the driving force for moving phosphate ions into the mitochondria is the proton motive force.
- The adenine nucleotide carrier is an antiporter and exchanges ADP and ATP across the inner membrane. The driving force is due to the ATP (-4) having a more negative charge than the ADP (-3) and thus it dissipates some of the electrical component of the proton electrochemical gradient.
The outcome of these transport processes using the proton electrochemical gradient is that more than 3 H+ are needed to make 1 ATP. Obviously this reduces the theoretical efficiency of the whole process and the likely maximum is closer to 28-30 ATP molecules.[2] In practice the efficiency may be even lower due to the inner membrane of the mitochondria being slightly leaky to protons.[3] Other factors may also dissipate the proton gradient creating an apparently leaky mitochondria. An uncoupling protein known as thermogenin is expressed in some cell types and is a channel that can transport protons. When this protein is active in the inner membrane it short circuits the coupling between the electron transport chain and ATP synthesis. The potential energy from the proton gradient is not used to make ATP but generates heat. This is particularly important in brown fat thermogenesis of newborn and hibernating mammals.
Fermentation
Main article: Fermentation (biochemistry)Without oxygen, pyruvate (pyruvic acid) is not metabolized by cellular respiration but undergoes a process of fermentation. The pyruvate is not transported into the mitochondrion, but remains in the cytoplasm, where it is converted to waste products that may be removed from the cell. This serves the purpose of oxidizing the electron carriers so that they can perform glycolysis again and removing the excess pyruvate. Fermentation oxidizes NADH to NAD+ so it can be re-used in glycolysis. In the absence of oxygen, fermentation prevents the build up of NADH in the cytoplasm and provides NAD+ for glycolysis. This waste product varies depending on the organism. In skeletal muscles, the waste product is lactic acid. This type of fermentation is called lactic acid fermentation. In yeast, the waste products are ethanol and carbon dioxide. This type of fermentation is known as alcoholic or ethanol fermentation. The ATP generated in this process is made by substrate-level phosphorylation, which does not require oxygen.
Fermentation is less efficient at using the energy from glucose since 2 ATP are produced per glucose, compared to the 38 ATP per glucose produced by aerobic respiration. This is because the waste products of fermentation still contain plenty of energy. Ethanol, for example, can be used in gasoline (petrol) solutions. Glycolytic ATP, however, is created more quickly. For prokaryotes to continue a rapid growth rate when they are shifted from an aerobic environment to an anaerobic environment, they must increase the rate of the glycolytic reactions. For multicellular organisms, during short bursts of strenuous activity, muscle cells use fermentation to supplement the ATP production from the slower aerobic respiration, so fermentation may be used by a cell even before the oxygen levels are depleted, as is the case in sports that do not require athletes to pace themselves, such as sprinting.
Anaerobic respiration
Main article: Anaerobic RespirationAnaerobic respiration is used by some microorganisms in which neither oxygen (aerobic respiration) nor pyruvate or pyruvate derivative (fermentation) is the final electron acceptor. Rather, an inorganic acceptor (for example, Sulfur) is used.
See also
- Tetrazolium chloride: cellular respiration indicator
- Maintenance respiration: maintenance as a functional component of cellular respiration
- Pasteur point
- Respirometry: research tool to explore cellular respiration
Pyruvate is decarboxylated to form acetylaldehyde and then to ethanol. There can be at the most 2 ATP molecules formed
References
- ^ Campbell, Neil A.; Brad Williamson; Robin J. Heyden (2006). Biology: Exploring Life. Boston, Massachusetts: Pearson Prentice Hall. ISBN 0-13-250882-6. http://www.phschool.com/el_marketing.html.
- ^ a b c Rich, P. R. (2003). "The molecular machinery of Keilin's respiratory chain". Biochemical Society Transactions 31 (Pt 6): 1095–1105. doi:10.1042/BST0311095. PMID 14641005.
- ^ Porter, R.; Brand, M. (1 September 1995). "Mitochondrial proton conductance and H+/O ratio are independent of electron transport rate in isolated hepatocytes" (Free full text). The Biochemical journal 310 ( Pt 2) (Pt 2): 379–382. ISSN 0264-6021. PMC 1135905. PMID 7654171. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1135905.</noinclude>h m
External links
- Chart of Important Metabolic Products
- A detailed description of respiration vs. fermentation
- Kimball's online resource to cellular respiration
- Cellular Respiration and Fermentation at Clermont College
Metabolism (Catabolism, Anabolism) General Cellular respiration Aerobic RespirationSpecific paths HumanNonhumanOtherNucleotide metabolismOtherbiochemical families: prot · nucl · carb (glpr, alco, glys) · lipd (fata/i, phld, strd, gllp, eico) · amac/i · ncbs/i · ttpy/iCategories:

Wikimedia Foundation. 2010.