- Lactic acid
-
Lactic acid 
L-Lactic acid 2-Hydroxypropanoic acidOther namesMilk acid
2-Hydroxypropanoic acidOther namesMilk acidIdentifiers CAS number 50-21-5  , 79-33-4 (L), 10326-41-7 (D)
, 79-33-4 (L), 10326-41-7 (D)ChemSpider 96860 ChEBI CHEBI:422 ChEMBL CHEMBL330546 ATC code G01,QP53AG02 Jmol-3D images Image 1 - CC(O)C(=O)O
- InChI=1S/C3H6O3/c1-2(4)3(5)6/h2,4H,1H3,(H,5,6)/t2-/m0/s1
Key: JVTAAEKCZFNVCJ-REOHCLBHSA-N
Properties Molecular formula C3H6O3 Molar mass 90.08 g mol−1 Melting point L: 53 °C
D: 53 °C
D/L: 16.8 °CBoiling point 122 °C @ 12 mmHg
Acidity (pKa) 3.86[1] Thermochemistry Std enthalpy of
combustion ΔcHo2981361.9 kJ/mol, 325.5 kcal/mol, 15.1 kJ/g, 3.61 kcal/g Related compounds Other anions lactate Related carboxylic acids acetic acid
glycolic acid
propionic acid
3-hydroxypropanoic acid
malonic acid
butyric acid
hydroxybutyric acidRelated compounds 1-propanol
2-propanol
propionaldehyde
acrolein
sodium lactateHazards GHS pictograms  [2]
[2]GHS hazard statements H315, H318[2] GHS precautionary statements P280, P305+351+338[2]  acid (verify) (what is:
acid (verify) (what is:  /
/ ?)
?)
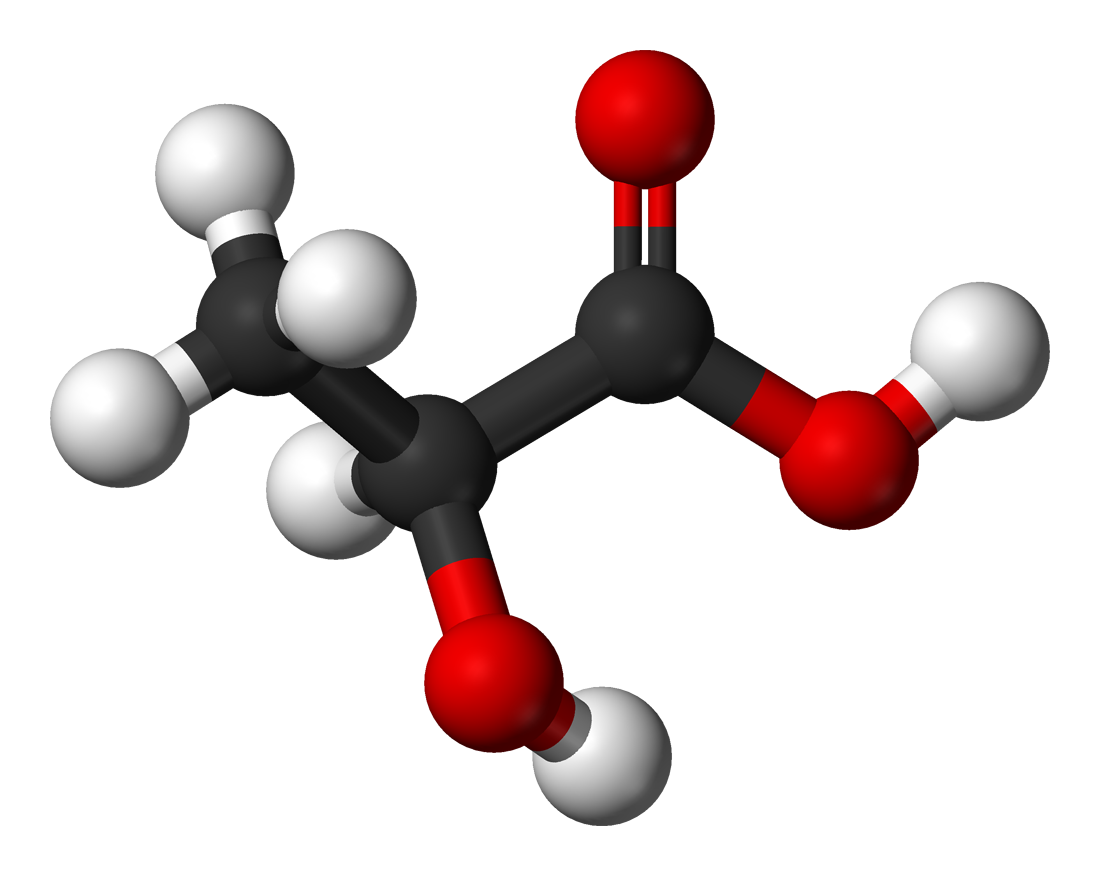
Except where noted otherwise, data are given for materials in their standard state (at 25 °C, 100 kPa)Infobox references Lactic acid, also known as milk acid, is a chemical compound that plays a role in various biochemical processes and was first isolated in 1780 by the Swedish chemist Carl Wilhelm Scheele. Lactic acid is a carboxylic acid with the chemical formula C3H6O3. It has a hydroxyl group adjacent to the carboxyl group, making it an alpha hydroxy acid (AHA).
In solution, it can lose a proton from the acidic group, producing the lactate ion (to be specific, an anion due to being negatively charged with an extra electron) CH3CH(OH)COO−. Compared to acetic acid, its pKa is 1 unit smaller, meaning lactic acid loses its proton ten times as easily as acetic acid does. This higher acidity is the consequence of the intramolecular hydrogen bridge between the α-hydroxyl and the carboxylate group, making the latter less capable of keeping its proton tight.
Lactic acid is miscible with water or ethanol, and is hygroscopic.
Lactic acid is chiral and has two optical isomers. One is known as L-(+)-lactic acid or (S)-lactic acid and the other, its mirror image, is D-(−)-lactic acid or (R)-lactic acid.
In animals, L-lactate is constantly produced from pyruvate via the enzyme lactate dehydrogenase (LDH) in a process of fermentation during normal metabolism and exercise. It does not increase in concentration until the rate of lactate production exceeds the rate of lactate removal, which is governed by a number of factors, including monocarboxylate transporters, concentration and isoform of LDH, and oxidative capacity of tissues. The concentration of blood lactate is usually 1–2 mmol/L at rest, but can rise to over 20 mmol/L during intense exertion.
In industry, lactic acid fermentation is performed by lactic acid bacteria. These bacteria can also grow in the mouth; the acid they produce is responsible for the tooth decay known as caries.[3][4][5][6]
In medicine, lactate is one of the main components of lactated Ringer's solution and Hartmann's solution. These intravenous fluids consist of sodium and potassium cations along with lactate and chloride anions in solution with distilled water, generally in concentrations isotonic with human blood. It is most commonly used for fluid resuscitation after blood loss due to trauma, surgery, or burn injury.
Contents
History
Lactic acid was refined for the first time by the Swedish chemist Carl Wilhelm Scheele in 1780 from sour milk. In 1808 Jöns Jacob Berzelius discovered that lactic acid (actually L-lactate) also is produced in muscles during exertion. Its structure was established by Johannes Wislicenus in 1873.
In 1856, Louis Pasteur discovered Lactobacillus and its role in the making of lactic acid. Lactic acid started to be produced commercially by the German pharmacy Boehringer Ingelheim in 1895.
In 2006, global production of lactic acid reached 275,000 tonnes with an average annual growth of 10%.[7]
Exercise and lactate
During power exercises such as sprinting, when the rate of demand for energy is high, lactate is produced faster than the ability of the tissues to remove it, so lactate concentration begins to rise. This is a beneficial process, since the regeneration of NAD+ ensures that energy production is maintained and exercise can continue. The increased lactate produced can be removed in a number of ways, including:
- Oxidation to pyruvate by well-oxygenated muscle cells.[citation needed]
- Pyruvate is then directly used to fuel the Krebs cycle.
- Conversion to glucose via gluconeogenesis in the liver and release back into the circulation; see the Cori cycle.[8]
- If not released, the glucose can be used to build up the liver's glycogen stores if they are empty.
Contrary to popular belief, this increased concentration of lactate does not directly cause acidosis, nor is it responsible for delayed onset muscle soreness.[9] This is because lactate itself is not capable of releasing a proton,[9] and, second, the acidic form of lactate, lactic acid, "is not produced in muscle".[10] Analysis of the glycolytic pathway in humans indicates that there are not enough hydrogen ions present in the glycolytic intermediates to produce lactic or any other acid.
The effect of lactate on acidosis has been the topic of many recent conferences in the field of exercise physiology. Robergs et al. have accurately chased the proton movement that occurs during glycolysis. However, in doing so, they have suggested that [H+] is an independent variable that determines its own concentration. A recent review by Lindinger et al.[10] has been written to rebut the stoichiometric approach used by Robergs et al.[9] In using this stoichiometric process, Robergs et al. have ignored the causative factors (independent variables) of the concentration of hydrogen ions (denoted [H+]). These factors are strong ion difference [SID], PCO2, and weak acid buffers. Lactate is a strong anion, and causes a reduction in [SID], which causes an increase in [H+] to maintain electroneutrality. PCO2 also causes an increase in [H+]. During exercise, the intramuscular lactate concentration and PCO2 increase, causing an increase in [H+], and, thus, a decrease in pH.[11] (See Le Chatelier's principle)
During intense exercise, the respiratory chain cannot keep up with the amount of hydrogen atoms that join to form NADH. NAD+ is required to oxidize 3-phosphoglyceraldehyde in order to maintain the production of anaerobic energy during glycolysis. During anaerobic glycolysis, NAD + “frees up” when extra nonoxidized hydrogens combine with a pyruvate molecule and then form lactate. If this does not occur, glycolysis comes to a stop. However, there is lactate being continually formed at rest and during moderate exercise. This occurs due to the metabolism of red blood cells that do not have mitochondria and limitations resulting from enzyme activity that occurs in muscle fibers that contain a high glycolytic capacity.[12]
 Reference ranges for blood tests, comparing blood content of lactate (shown in violet at center-right) with other constituents.
Reference ranges for blood tests, comparing blood content of lactate (shown in violet at center-right) with other constituents.
Brain metabolism
Although glucose is usually assumed to be the main energy source for living tissues, there are some indications that it is lactate, and not glucose, that is preferentially metabolized by neurons in the brain of several mammals species (the notable ones being mice, rats, and humans).[13][14] According to the lactate-shuttling hypothesis, glial cells are responsible for transforming glucose into lactate, and for providing lactate to the neurons.[15][16] Because of this local metabolic activity of glial cells, the extracellular fluid immediately surrounding neurons strongly differs in composition from the blood or cerebro-spinal fluid, being much richer with lactate, as it was found in microdialysis studies.[13]
The role of lactate for brain metabolism seems to be even more important at early stages of development (prenatal and early postnatal), with lactate at these stages having higher concentrations in body liquids, and being utilized by the brain even more preferentially over glucose.[13] It was also hypothesized that lactate may exert a strong action over GABAergic networks in the developing brain, making them more inhibitory than it was previously assumed,[17] acting either through better support of metabolites,[13] or alterations in base intracellular pH levels,[18][19] or both.[20]
A more recent paper by Zilberter's group looked directly at the energy metabolism features in brain slices of immature mice and showed that beta-hydroxybutyrate, lactate and pyruvate acted as oxidative energy substrates causing an increase in the NAD(P)H oxidation phase, that glucose was insufficient as an energy carrier during intense synaptic activity and finally, that lactate can be an efficient energy substrate capable of sustaining and enhancing brain aerobic energy metabolism in vitro. The paper was positively commented by Kasischke (2011): "The study by Ivanov et al. (2011) also provides novel data on biphasic NAD(P)H fluorescence transients, an important physiological response to neural activation that has been reproduced in many studies and that is believed to originate predominately from activity-induced concentration changes to the cellular NADH pools."
Polymer precursor
Main article: polylactic acidTwo molecules of lactic acid can be dehydrated to lactide, a cyclic lactone. A variety of catalysts can polymerize lactide to either heterotactic or syndiotactic polylactide, which as biodegradable polyesters with valuable (inter alia) medical properties are currently attracting much attention.
Lactic acid is used also as a monomer for producing polylactic acid (PLA), which later has developed application as biodegradable plastic. This kind of plastic is a good option for substituting conventional plastic produced from petroleum oil because of low emission of carbon dioxide. The commonly used process in producing lactic acid is via fermentation, and, later, to obtain the polylactic acid, the polymerization process follows.
Pharmaceutical and cosmetic applications
Lactic acid is also employed in pharmaceutical technology to produce water-soluble lactates from otherwise insoluble active ingredients. It finds further use in topical preparations and cosmetics to adjust acidity and for its disinfectant and keratolytic properties.
Foods
Lactic acid is found primarily in sour milk products, such as koumiss, laban, yogurt, kefir, and some cottage cheeses. The casein in fermented milk is coagulated (curdled) by lactic acid. Lactic acid is also responsible for the sour flavor of sourdough breads. This acid is used in beer brewing to lower the pH and increase the body of the beer. Some brewers and breweries will use food grade lactic acid to lower the ph in finished beers.
In winemaking, a bacterial process, natural or controlled, is often used to convert the naturally present malic acid to lactic acid, to reduce the sharpness and for other flavor-related reasons. This malolactic fermentation is undertaken by the family of lactic acid bacteria.
As a food additive it is approved for use in the EU,[21] USA[22] and Australia and New Zealand;[23] it is listed by its INS number 270 or as E number E270.
Detergents
Lactic acid has gained importance in the detergent industry the last decade. It is a good descaler, soap-scum remover, and a registered anti-bacterial agent. It is also economically beneficial as well as part of a trend toward environmentally safer and natural ingredients.
See also
References
- ^ Dawson, R. M. C. et al., Data for Biochemical Research, Oxford, Clarendon Press, 1959.
- ^ a b c Online Sigma Catalogue , accessdate: June 16, 2011.
- ^ Badet, C; Thebaud, NB (2008). "Ecology of lactobacilli in the oral cavity: a review of literature". The open microbiology journal 2: 38–48. doi:10.2174/1874285800802010038. PMC 2593047. PMID 19088910. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2593047.
- ^ Nascimento, MM; Gordan, VV; Garvan, CW; Browngardt, CM; Burne, RA (2009). "Correlations of oral bacterial arginine and urea catabolism with caries experience". Oral microbiology and immunology 24 (2): 89–95. doi:10.1111/j.1399-302X.2008.00477.x. PMC 2742966. PMID 19239634. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2742966.
- ^ Aas, JA; Griffen, AL; Dardis, SR; Lee, AM; Olsen, I; Dewhirst, FE; Leys, EJ; Paster, BJ (2008). "Bacteria of dental caries in primary and permanent teeth in children and young adults". Journal of clinical microbiology 46 (4): 1407–17. doi:10.1128/JCM.01410-07. PMC 2292933. PMID 18216213. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2292933.
- ^ Caufield, PW; Li, Y; Dasanayake, A; Saxena, D (2007). "Diversity of Lactobacilli in the Oral Cavities of Young Women with Dental Caries". Caries Res. 41 (1): 2–8. doi:10.1159/000096099. PMC 2646165. PMID 17167253. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2646165.
- ^ NNFCC Renewable Chemicals Factsheet: Lactic Acid
- ^ McArdle, W. D., Katch, F. I., & Katch, V. L. (2010). Exercise physiology: Energy, nutrition, and human performance. Wolters Kluwer/Lippincott Williams & Wilkins Health. ISBN 0683057316 9780683057317.
- ^ a b c Robergs, RA; Ghiasvand, F; Parker, D (2004). "Biochemistry of exercise-induced metabolic acidosis". Am J Physiol Regul Integr Comp Physiol 287 (3): R502–R516. doi:10.1152/ajpregu.00114.2004. PMID 15308499. http://ajpregu.physiology.org/cgi/reprint/287/3/R502.pdf.
- ^ a b Lindinger, M. I. (2004). "Applying physicochemical principles to skeletal muscle acid-base status". Am J Physiol Regul Integr Comp Physiol 289 (3): R890–94. doi:10.1152/ajpregu.00225.2005.
- ^ Robergs, RA; Ghiasvand, F; Parker, D (2004). "Biochemistry of exercise-induced metabolic acidosis". Am J Physiol Regul Integr Comp Physiol 287 (3): R502-R516. doi:10.1152/ajpregu.00114.2004. PMID 15308499.
- ^ McArdle, Katch & Katch (2010). Exercise Physiology: Energy, Nutrition, and Human Performance. Wolters Kluwer/Lippincott Williams & Wilkins Health. ISBN 0683057316 9780683057317.
- ^ a b c d Zilberter Y, Zilberter T, Bregestovski P (September 2010). "Neuronal activity in vitro and the in vivo reality: the role of energy homeostasis". Trends Pharmacol. Sci. 31 (9): 394–401. doi:10.1016/j.tips.2010.06.005. PMID 20633934.
- ^ Wyss MT, Jolivet R, Buck A, Magistretti PJ, Weber B (May 2011). "In vivo evidence for lactate as a neuronal energy source". J. Neurosci. 31 (20): 7477–85. doi:10.1523/JNEUROSCI.0415-11.2011. PMID 21593331.
- ^ Gladden LB (July 2004). "Lactate metabolism: a new paradigm for the third millennium". J. Physiol. (Lond.) 558 (Pt 1): 5–30. doi:10.1113/jphysiol.2003.058701. PMC 1664920. PMID 15131240. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1664920.
- ^ Pellerin L, Bouzier-Sore AK, Aubert A, et al. (September 2007). "Activity-dependent regulation of energy metabolism by astrocytes: an update". Glia 55 (12): 1251–62. doi:10.1002/glia.20528. PMID 17659524.
- ^ Holmgren CD, Mukhtarov M, Malkov AE, Popova IY, Bregestovski P, Zilberter Y (February 2010). "Energy substrate availability as a determinant of neuronal resting potential, GABA signaling and spontaneous network activity in the neonatal cortex in vitro". J. Neurochem. 112 (4): 900–12. doi:10.1111/j.1471-4159.2009.06506.x. PMID 19943846.
- ^ Tyzio R, Allene C, Nardou R, et al. (January 2011). "Depolarizing actions of GABA in immature neurons depend neither on ketone bodies nor on pyruvate". J. Neurosci. 31 (1): 34–45. doi:10.1523/JNEUROSCI.3314-10.2011. PMID 21209187.
- ^ Ruusuvuori E, Kirilkin I, Pandya N, Kaila K (November 2010). "Spontaneous network events driven by depolarizing GABA action in neonatal hippocampal slices are not attributable to deficient mitochondrial energy metabolism". J. Neurosci. 30 (46): 15638–42. doi:10.1523/JNEUROSCI.3355-10.2010. PMID 21084619.
- ^ Khakhalin AS (May 2011). "Questioning the depolarizing effects of GABA during early brain development". J Neurophysiol. doi:10.1152/jn.00293.2011. PMID 21593390.
- ^ UK Food Standards Agency: "Current EU approved additives and their E Numbers". http://www.food.gov.uk/safereating/chemsafe/additivesbranch/enumberlist. Retrieved 2011-10-27.
- ^ US Food and Drug Administration: "Listing of Food Additives Status Part II". http://www.fda.gov/Food/FoodIngredientsPackaging/FoodAdditives/ucm191033.htm#ftnT. Retrieved 2011-10-27.
- ^ Australia New Zealand Food Standards Code"Standard 1.2.4 - Labelling of ingredients". http://www.comlaw.gov.au/Details/F2011C00827. Retrieved 2011-10-27.
External links
Categories:- Food acidity regulators
- Hydroxy acids
- Exercise physiology
- Preservatives


Wikimedia Foundation. 2010.