- Glutathione
-
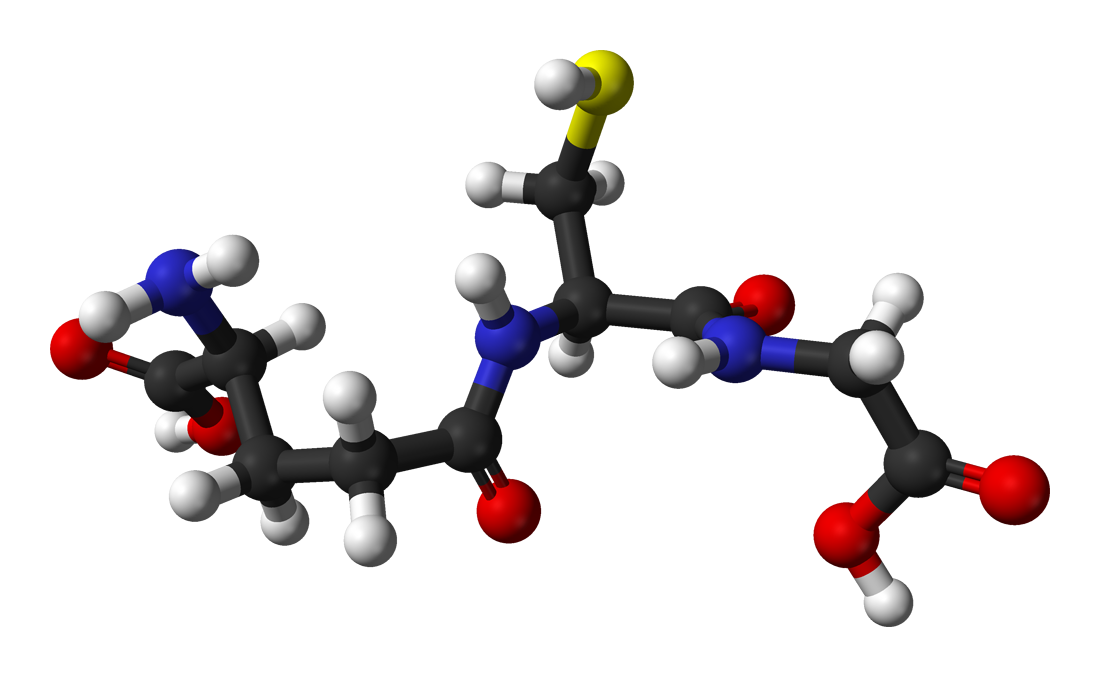
Glutathione[1] 
 (2S)-2-amino-4-{[(1R)-1-[(carboxymethyl)carbamoyl]-2-sulfanylethyl]carbamoyl}butanoic acidOther namesγ-L-Glutamyl-L-cysteinylglycine
(2S)-2-amino-4-{[(1R)-1-[(carboxymethyl)carbamoyl]-2-sulfanylethyl]carbamoyl}butanoic acidOther namesγ-L-Glutamyl-L-cysteinylglycine
(2S)-2-Amino-5-[[(2R)-1-(carboxymethylamino)-1-oxo- 3-sulfanylpropan-2-yl]amino]-5-oxopentanoic acidIdentifiers Abbreviations GSH CAS number 70-18-8 
PubChem 124886 ChemSpider 111188 UNII GAN16C9B8O DrugBank DB00143 KEGG C00051 MeSH Glutathione ChEBI CHEBI:60836 ChEMBL CHEMBL1543 Jmol-3D images Image 1 - C(CC(=O)N[C@@H](CS)C(=O)NCC(=O)O)[C@@H](C(=O)O)N
- InChI=1S/C10H17N3O6S/c11-5(10(18)19)1-2-7(14)13-6(4-20)9(17)12-3-8(15)16/h5-6,20H,1-4,11H2,(H,12,17)(H,13,14)(H,15,16)(H,18,19)/t5-,6-/m0/s1
Key: RWSXRVCMGQZWBV-WDSKDSINSA-N
InChI=1/C10H17N3O6S/c11-5(10(18)19)1-2-7(14)13-6(4-20)9(17)12-3-8(15)16/h5-6,20H,1-4,11H2,(H,12,17)(H,13,14)(H,15,16)(H,18,19)/t5-,6-/m0/s1
Key: RWSXRVCMGQZWBV-WDSKDSINBK
Properties Molecular formula C10H17N3O6S Molar mass 307.32 g/mol Melting point 195 °C, 468 K, 383 °F
Solubility in water Freely soluble[1] Solubility in methanol, diethyl ether Insoluble  (verify) (what is:
(verify) (what is:  /
/ ?)
?)
Except where noted otherwise, data are given for materials in their standard state (at 25 °C, 100 kPa)Infobox references Glutathione (GSH) is a tripeptide that contains an unusual peptide linkage between the amine group of cysteine (which is attached by normal peptide linkage to a glycine) and the carboxyl group of the glutamate side-chain. It is an antioxidant, preventing damage to important cellular components caused by reactive oxygen species such as free radicals and peroxides.[2]
Thiol groups are reducing agents, existing at a concentration of approximately 5 mM in animal cells. Glutathione reduces disulfide bonds formed within cytoplasmic proteins to cysteines by serving as an electron donor. In the process, glutathione is converted to its oxidized form glutathione disulfide (GSSG), also called L(-)-Glutathione.
Glutathione is found almost exclusively in its reduced form, since the enzyme that reverts it from its oxidized form, glutathione reductase, is constitutively active and inducible upon oxidative stress. In fact, the ratio of reduced glutathione to oxidized glutathione within cells is often used as a measure of cellular toxicity.[3]
Contents
Biosynthesis
Glutathione is not an essential nutrient (meaning it does not have to be obtained via food), since it can be synthesized in the body from the amino acids L-cysteine, L-glutamic acid, and glycine. The sulfhydryl (thiol) group (SH) of cysteine serves as a proton donor and is responsible for the biological activity of glutathione. Provision of this amino acid is the rate-limiting factor in glutathione synthesis by the cells, since cysteine is relatively rare in foodstuffs. Furthermore, if released as the free amino acid, cysteine is toxic and spontaneously catabolized in the gastrointestinal tract and blood plasma.[4]
Glutathione is synthesized in two adenosine triphosphate-dependent steps:
- First, gamma-glutamylcysteine is synthesized from L-glutamate and cysteine via the enzyme gamma-glutamylcysteine synthetase (a.k.a. glutamate cysteine ligase, GCL). This reaction is the rate-limiting step in glutathione synthesis.[5]
- Second, glycine is added to the C-terminal of gamma-glutamylcysteine via the enzyme glutathione synthetase.
Animal glutamate cysteine ligase (GCL) is a heterodimeric enzyme composed of a catalytic (GCLC) and modulatory (GCLM) subunit. GCLC constitutes all the enzymatic activity, whereas GCLM increases the catalytic efficiency of GCLC. Mice lacking GCLC (i.e., all de novo GSH synthesis) die before birth.[6] Mice lacking GCLM demonstrate no outward phenotype, but exhibit marked decrease in GSH and increased sensitivity to toxic insults.[7][8][9]
While all cells in the human body are capable of synthesizing glutathione, liver glutathione synthesis has been shown to be essential. Mice with genetically-induced loss of GCLC (i.e., GSH synthesis) only in the liver die within 1 month of birth.[10]
The plant glutamate cysteine ligase (GCL) is a redox-sensitive homodimeric enzyme, conserved in the plant kingdom.[11] In an oxidizing environment, intermolecular disulfide bridges are formed and the enzyme switches to the dimeric active state. The mid-point potential of the critical cysteine pair is -318 mV. In addition to the redox-dependent control is the plant GCL enzyme feedback inhibited by GSH.[12] GCL is exclusively located in plastids, and glutathione synthetase is dual-targeted to plastids and cytosol, thus are GSH and gamma-glutamylcysteine exported from the plastids.[13] Both glutathione biosynthesis enzymes are essential in plants; knock-outs of GCL and GS are lethal to embryo and seedling.[14]
The biosynthesis pathway for glutathione is found in some bacteria, like cyanobacteria and proteobacteria, but is missing in many other bacteria. Most eukaryotes synthesize glutathione, including humans, but some do not, such as Leguminosae, Entamoeba, and Giardia. The only archaea that make glutathione are halobacteria.[15][16]
Function
Glutathione exists in reduced (GSH) and oxidized (GSSG) states. In the reduced state, the thiol group of cysteine is able to donate a reducing equivalent (H++ e-) to other unstable molecules, such as reactive oxygen species. In donating an electron, glutathione itself becomes reactive, but readily reacts with another reactive glutathione to form glutathione disulfide (GSSG). Such a reaction is possible due to the relatively high concentration of glutathione in cells (up to 5 mM in the liver). GSH can be regenerated from GSSG by the enzyme glutathione reductase.
In healthy cells and tissue, more than 90% of the total glutathione pool is in the reduced form (GSH) and less than 10% exists in the disulfide form (GSSG). An increased GSSG-to-GSH ratio is considered indicative of oxidative stress.
Glutathione has multiple functions:
- It is the major endogenous antioxidant produced by the cells, participating directly in the neutralization of free radicals and reactive oxygen compounds, as well as maintaining exogenous antioxidants such as vitamins C and E in their reduced (active) forms.[17]
- Regulation of the nitric oxide cycle, which is critical for life but can be problematic if unregulated[18]
- It is used in metabolic and biochemical reactions such as DNA synthesis and repair, protein synthesis, prostaglandin synthesis, amino acid transport, and enzyme activation. Thus, every system in the body can be affected by the state of the glutathione system, especially the immune system, the nervous system, the gastrointestinal system and the lungs.[4]
Function in animals
GSH is known as a substrate in both conjugation reactions and reduction reactions, catalyzed by glutathione S-transferase enzymes in cytosol, microsomes, and mitochondria. However, it is also capable of participating in non-enzymatic conjugation with some chemicals.
In the case of N-acetyl-p-benzoquinone imine (NAPQI), the reactive cytochrome P450-reactive metabolite formed by paracetamol (or acetaminophen as it is known in the US), which becomes toxic when GSH is depleted by an overdose of acetaminophen, glutathione is an essential antidote to overdose. Glutathione conjugates to NAPQI and helps to detoxify it. In this capacity, it protects cellular protein thiol groups, which would otherwise become covalently modified; when all GSH has been spent, NAPQI begins to react with the cellular proteins, killing the cells in the process. The preferred treatment for an overdose of this painkiller is the administration (usually in atomized form) of N-acetyl-L-cysteine (often as a trademarked preparation called Mucomyst® [1]), which is processed by cells to L-cysteine and used in the de novo synthesis of GSH.
Glutathione (GSH) participates in leukotriene synthesis and is a cofactor for the enzyme glutathione peroxidase. It is also important as a hydrophilic molecule that is added to lipophilic toxins and waste in the liver during biotransformation before they can become part of the bile. Glutathione is also needed for the detoxification of methylglyoxal, a toxin produced as a by-product of metabolism.
This detoxification reaction is carried out by the glyoxalase system. Glyoxalase I (EC 4.4.1.5) catalyzes the conversion of methylglyoxal and reduced glutathione to S-D-lactoyl-glutathione. Glyoxalase II (EC 3.1.2.6) catalyzes the hydrolysis of S-D-lactoyl-glutathione to glutathione and D-lactic acid.
Glutathione has recently been used as an inhibitor of melanin in the cosmetics industry. In countries like Japan and the Philippines, this product is sold as a whitening soap. Glutathione competitively inhibits melanin synthesis in the reaction of tyrosinase and L-DOPA by interrupting L-DOPA's ability to bind to tyrosinase during melanin synthesis. The inhibition of melanin synthesis was reversed by increasing the concentration of L-DOPA, but not by increasing tyrosinase. Although the synthesized melanin was aggregated within 1 h, the aggregation was inhibited by the addition of glutathione. These results indicate that glutathione inhibits the synthesis and agglutination of melanin by interrupting the function of L-DOPA."[19]
Function in plants
In plants, glutathione is crucial for biotic and abiotic stress management. It is a pivotal component of the glutathione-ascorbate cycle, a system that reduces poisonous hydrogen peroxide.[20] It is the precursor of phytochelatins, glutathione oligomeres that chelate heavy metals such as cadmium.[21] Glutathione is required for efficient defence against plant pathogens such as Pseudomonas syringae and Phytophthora brassicae.[22] APS reductase, an enzyme of the sulfur assimilation pathway uses glutathione as electron donor. Other enzymes using glutathione as substrate are glutaredoxin, these small oxidoreductases are involved in flower development, salicylic acid and plant defence signalling.[23]
Supplementation
Raising GSH levels through direct supplementation of glutathione is difficult. Research suggests that glutathione taken orally is not well absorbed across the gastrointestinal tract. In a study of acute oral administration of a very large dose (3 grams) of oral glutathione, Witschi and coworkers found "it is not possible to increase circulating glutathione to a clinically beneficial extent by the oral administration of a single dose of 3 g of glutathione."[24][25]
Calcitriol, the active metabolite of vitamin D synthesized in the kidney, increases glutathione levels in the brain and appears to be a catalyst for glutathione production.[26]
In addition, plasma and liver GSH concentrations can be raised by administration of certain supplements that serve as GSH precursors. N-acetylcysteine, commonly referred to as NAC, is the most bioavailable precursor of glutathione.[27] Other supplements, including S-adenosylmethionine (SAMe)[28][29][30] and whey protein[31][32][33][34][35][36] have also been shown to increase glutathione content within the cell.
NAC is available both as a drug and as a generic supplement. Alpha lipoic acid has also been shown to restore intracellular glutathione.[37][38] Melatonin has been shown to stimulate a related enzyme, glutathione peroxidase,[39] and silymarin, an extract of the seeds of the milk thistle plant (Silybum marianum), has also demonstrated an ability to replenish glutathione levels.[40][41]
Glutathione is a tightly regulated intracellular constituent, and is limited in its production by negative feedback inhibition of its own synthesis through the enzyme gamma-glutamylcysteine synthetase, thus greatly minimizing any possibility of overdosage. Glutathione augmentation using presursors of glutathione synthesis or intravenous glutathione is a strategy developed to address states of glutathione deficiency, high oxidative stress, immune deficiency, and xenobiotic overload in which glutathione plays a part in the detoxification of the xenobiotic in question (especially through the hepatic route). Glutathione deficiency states include, but are not limited to, HIV/AIDS, chemical and infectious hepatitis, myalgic encephalomyelitis chronic fatigue syndrome ME / CFS,[42][43][44] prostate and other cancers, cataracts, Alzheimer's disease, Parkinson's disease, chronic obstructive pulmonary disease, asthma, radiation poisoning, malnutritive states, arduous physical stress, and aging, and has been associated with suboptimal immune response. Many clinical pathologies are associated with oxidative stress and are elaborated upon in numerous medical references.,[4][45][46]
Low glutathione is also strongly implicated in wasting and negative nitrogen balance,[47] as seen in cancer, AIDS, sepsis, trauma, burns and even athletic overtraining. Glutathione supplementation can oppose this process, and in AIDS, for example, result in improved survival rates.[48] However, studies in many of these conditions have not been able to differentiate between low glutathione as a result of acutely (as in septic patients) or chronically (as in HIV) increased oxidative stress, and increased pathology as a result of preexisting deficiencies.
Schizophrenia and bipolar disorder are associated with lowered glutathione. Accruing data suggest that oxidative stress may be a factor underlying the pathophysiology of bipolar disorder (BD), major depressive disorder (MDD), and schizophrenia (SCZ). Glutathione (GSH) is the major free radical scavenger in the brain.[49] Diminished GSH levels elevate cellular vulnerability towards oxidative stress; characterized by accumulating reactive oxygen species. Replenishment of glutathione using N-acetyl cysteine has been shown to reduce symptoms of both disorders.[citation needed]
Cancer
Preliminary results indicate glutathione changes the level of reactive oxygen species in isolated cells grown in a laboratory,[50][51] which may reduce cancer development.[52] [53] None of these tests were performed in humans.
However, once a cancer has already developed, by conferring resistance to a number of chemotherapeutic drugs, elevated levels of glutathione in tumour cells are able to protect cancerous cells in bone marrow, breast, colon, larynx, and lung cancers.[54]
Pathology
Excess glutamate at synapses, which may be released in conditions such as traumatic brain injury, can prevent the uptake of cysteine, a necessary building-block of glutathione. Without the protection from oxidative injury afforded by glutathione, cells may be damaged or killed.[55]
Methods to determine glutathione
Reduced glutathione may be visualized using Ellman's reagent or bimane derivates such as monobromobimane. The monobromobimane method is more sensitive. In this procedure, cells are lysed and thiols extracted using a HCl buffer. The thiols are then reduced with dithiothreitol (DTT) and labelled by monobromobimane. Monobromobimane becomes fluorescent after binding to GSH. The thiols are then separated by HPLC and the fluorescence quantified with a fluorescence detector. Bimane may also be used to quantify glutathione in vivo. The quantification is done by confocal laser scanning microscopy after application of the dye to living cells.[56] Another approach, which allows to measure the glutathione redox potential at a high spatial and temporal resolution in living cells is based on redox imaging using the redox-sensitive green fluorescent protein (roGFP)[57] or redox sensitive yellow fluorescent protein (rxYFP) [58]
See also
- Glutathione synthetase deficiency
- Ophthalmic acid
- roGFP, a tool to measure the cellular glutathione redox potential
- Glutathione-ascorbate cycle
- Bacterial glutathione transferase
- Thioredoxin Cysteine-containing small proteins with very similar functions as reducing agents
References
- ^ a b Merck Index, 11th Edition, 4369
- ^ Pompella, A; Visvikis, A; Paolicchi, A; De Tata, V; Casini, AF (2003). "The changing faces of glutathione, a cellular protagonist". Biochemical Pharmacology 66 (8): 1499–503. doi:10.1016/S0006-2952(03)00504-5. PMID 14555227.
- ^ Pastore, Anna; Piemonte, Fiorella; Locatelli, Mattia; Russo, Anna Lo; Gaeta, Laura Maria; Tozzi, Giulia; Federici, Giorgio (2003). "Determination of blood total, reduced, and oxidized glutathione in pediatric subjects". Clinical Chemistry 47 (8): 1467–9. PMID 11468240.
- ^ a b c http://www.drugs.com/pdr/immunocal-powder-sachets.html
- ^ http://www.sciencedirect.com/science/article/pii/S000326970300143X, someone please correct Wiki notation
- ^ Dalton, T; Dieter, MZ; Yang, Y; Shertzer, HG; Nebert, DW (2000). "Knockout of the Mouse Glutamate Cysteine Ligase Catalytic Subunit (Gclc) Gene: Embryonic Lethal When Homozygous, and Proposed Model for Moderate Glutathione Deficiency When Heterozygous". Biochemical and Biophysical Research Communications 279 (2): 324–9. doi:10.1006/bbrc.2000.3930. PMID 11118286.
- ^ Yang, Y.; Dieter, MZ; Chen, Y; Shertzer, HG; Nebert, DW; Dalton, TP (2002). "Initial characterization of the glutamate-cysteine ligase modifier subunit Gclm(-/-) knockout mouse. Novel model system for a severely compromised oxidative stress response". Journal of Biological Chemistry 277 (51): 49446–52. doi:10.1074/jbc.M209372200. PMID 12384496.
- ^ Giordano, G; Afsharinejad, Z; Guizzetti, M; Vitalone, A; Kavanagh, T; Costa, L (2007). "Organophosphorus insecticides chlorpyrifos and diazinon and oxidative stress in neuronal cells in a genetic model of glutathione deficiency". Toxicology and Applied Pharmacology 219 (2–3): 181–9. doi:10.1016/j.taap.2006.09.016. PMID 17084875.
- ^ McConnachie, L. A.; Mohar, I.; Hudson, F. N.; Ware, C. B.; Ladiges, W. C.; Fernandez, C.; Chatterton-Kirchmeier, S.; White, C. C. et al. (2007). "Glutamate Cysteine Ligase Modifier Subunit Deficiency and Gender as Determinants of Acetaminophen-Induced Hepatotoxicity in Mice". Toxicological Sciences 99 (2): 628–36. doi:10.1093/toxsci/kfm165. PMID 17584759.
- ^ Chen, Ying; Yang, Yi; Miller, Marian L.; Shen, Dongxiao; Shertzer, Howard G.; Stringer, Keith F.; Wang, Bin; Schneider, Scott N. et al. (2007). "Hepatocyte-specificGclcdeletion leads to rapid onset of steatosis with mitochondrial injury and liver failure". Hepatology 45 (5): 1118–28. doi:10.1002/hep.21635. PMID 17464988.
- ^ Hothorn, M.; Wachter, A; Gromes, R; Stuwe, T; Rausch, T; Scheffzek, K (2006). "Structural Basis for the Redox Control of Plant Glutamate Cysteine Ligase". Journal of Biological Chemistry 281 (37): 27557–65. doi:10.1074/jbc.M602770200. PMID 16766527.
- ^ Hicks, L. M.; Cahoon, R. E.; Bonner, E. R.; Rivard, R. S.; Sheffield, J.; Jez, J. M. (2007). "Thiol-Based Regulation of Redox-Active Glutamate-Cysteine Ligase from Arabidopsis thaliana". The Plant Cell Online 19 (8): 2653–61. doi:10.1105/tpc.107.052597. PMC 2002632. PMID 17766407. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2002632.
- ^ Wachter, Andreas; Wolf, Sebastian; Steininger, Heike; Bogs, Jochen; Rausch, Thomas (2004). "Differential targeting of GSH1 and GSH2 is achieved by multiple transcription initiation: implications for the compartmentation of glutathione biosynthesis in the Brassicaceae". The Plant Journal 41 (1): 15–30. doi:10.1111/j.1365-313X.2004.02269.x. PMID 15610346.
- ^ Pasternak, Maciej; Lim, Benson; Wirtz, Markus; Hell, RüDiger; Cobbett, Christopher S.; Meyer, Andreas J. (2007). "Restricting glutathione biosynthesis to the cytosol is sufficient for normal plant development". The Plant Journal 53 (6): 999–1012. doi:10.1111/j.1365-313X.2007.03389.x. PMID 18088327.
- ^ Copley, Shelley D; Dhillon, Jasvinder K (2002). Genome Biology 3 (5): research0025.1. doi:10.1186/gb-2002-3-5-research0025.
- ^ Grill D, Tausz T, De Kok LJ (2001). Significance of glutathione in plant adaptation to the environment. Springer. ISBN 1402001789. http://books.google.com/?id=aX2eJf1i67IC&pg=PA13.
- ^ Scholz RW. Graham KS. Gumpricht E. Reddy CC. Mechanism of interaction of vitamin E and glutathione in the protection against membrane lipid peroxidation. Ann NY Acad Sci 1989:570:514-7. Hughes RE. Reduction of dehydroascorbic acid by animal tissues.Nature 1964:203:1068-9.
- ^ Clementi, Emilio; Brown, Guy Charles; Feelisch, Martin; Moncada, Salvador; Bugg, S; O'Connell, MJ; Goldsbrough, PB; Cobbett, CS (1999). "Phytochelatin synthase genes from Arabidopsis and the yeast Schizosaccharomyces pombe". The Plant cell 11 (6): 1153–64. doi:10.1105/tpc.11.6.1153. JSTOR 3870806. PMC 144235. PMID 10368185. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=144235.
- ^ Matsuki, Mitsuo; Watanabe, Toshihiko; Ogasawara, Ayako; Mikami, Takeshi; Matsumoto, Tatsuji (2008). "Inhibitory Mechanism of Melanin Synthesis by Glutathione". Yakugaku Zasshi 128 (8): 1203–7. doi:10.1248/yakushi.128.1203. PMID 18670186.
- ^ Noctor, Graham; Foyer, Christine H. (1998). "ASCORBATE AND GLUTATHIONE: Keeping Active Oxygen Under Control". Annual Review of Plant Physiology and Plant Molecular Biology 49: 249–279. doi:10.1146/annurev.arplant.49.1.249. PMID 15012235.
- ^ Ha, S.-B.; Smith, AP; Howden, R; Dietrich, WM; Bugg, S; O'Connell, MJ; Goldsbrough, PB; Cobbett, CS (1999). "Phytochelatin Synthase Genes from Arabidopsis and the Yeast Schizosaccharomyces pombe". The Plant Cell Online 11 (6): 1153–64. doi:10.1105/tpc.11.6.1153. PMC 144235. PMID 10368185. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=144235.
- ^ Parisy, Vincent; Poinssot, Benoit; Owsianowski, Lucas; Buchala, Antony; Glazebrook, Jane; Mauch, Felix (2006). "Identification of PAD2 as a γ-glutamylcysteine synthetase highlights the importance of glutathione in disease resistance of Arabidopsis". The Plant Journal 49 (1): 159–72. doi:10.1111/j.1365-313X.2006.02938.x. PMID 17144898.
- ^ Rouhier, Nicolas; Lemaire, StéPhane D.; Jacquot, Jean-Pierre (2008). "The Role of Glutathione in Photosynthetic Organisms: Emerging Functions for Glutaredoxins and Glutathionylation". Annual Review of Plant Biology 59: 143–66. doi:10.1146/annurev.arplant.59.032607.092811. PMID 18444899.
- ^ Witschi, A.; Reddy, S.; Stofer, B.; Lauterburg, B. H. (1992). "The systemic availability of oral glutathione". European Journal of Clinical Pharmacology 43 (6): 667–9. doi:10.1007/BF02284971. PMID 1362956.
- ^ AIDS Line Update
- ^ Garcion, E; Wionbarbot, N; Monteromenei, C; Berger, F; Wion, D (2002). "New clues about vitamin D functions in the nervous system". Trends in Endocrinology and Metabolism 13 (3): 100–5. doi:10.1016/S1043-2760(01)00547-1. PMID 11893522.
- ^ Gross, Clark L.; Innace, Joy K.; Hovatter, Renee C.; Meier, Henry L.; Smith, William J. (1993). "Biochemical manipulation of intracellular glutathione levels influences cytotoxicity to isolated human lymphocytes by sulfur mustard". Cell Biology and Toxicology 9 (3): 259–67. doi:10.1007/BF00755604. PMID 8299004.
- ^ Lieber, Charles S. (2002). "S-adenosyl-L-methionine: its role in the treatment of liver disorders". The American journal of clinical nutrition 76 (5): 1183S–7S. PMID 12418503.
- ^ Vendemiale, G.; Altomare, E.; Trizio, T.; Le Grazie, C.; Di Padova, C.; Salerno, M. T.; Carrieri, V.; Albano, O. (1989). "Effects of Oral S-Adenosyl-l-Methionine on Hepatic Glutathione in Patients with Liver Disease". Scandinavian Journal of Gastroenterology 24 (4): 407–15. doi:10.3109/00365528909093067. PMID 2781235.
- ^ Loguercio, C; Nardi, G; Argenzio, F; Aurilio, C; Petrone, E; Grella, A; Del Vecchio Blanco, C; Coltorti, M (1994). "Effect of S-adenosyl-L-methionine administration on red blood cell cysteine and glutathione levels in alcoholic patients with and without liver disease". Alcohol and alcoholism (Oxford, Oxfordshire) 29 (5): 597–604. PMID 7811344.
- ^ Micke, P.; Beeh, K. M.; Schlaak, J. F.; Buhl, R. (2001). "Oral supplementation with whey proteins increases plasma glutathione levels of HIV-infected patients". European Journal of Clinical Investigation 31 (2): 171–8. doi:10.1046/j.1365-2362.2001.00781.x. PMID 11168457.
- ^ Moreno, Y. F.; Sgarbieri, VC; Da Silva, MN; Toro, AA; Vilela, MM (2006). "Features of Whey Protein Concentrate Supplementation in Children with Rapidly Progressive HIV Infection". Journal of Tropical Pediatrics 52 (1): 34–8. doi:10.1093/tropej/fmi074. PMID 16014759.
- ^ Grey, V; Mohammed, SR; Smountas, AA; Bahlool, R; Lands, LC (2003). "Improved glutathione status in young adult patients with cystic fibrosis supplemented with whey protein". Journal of Cystic Fibrosis 2 (4): 195–8. doi:10.1016/S1569-1993(03)00097-3. PMID 15463873.
- ^ Micke, P.; Beeh, K. M.; Buhl, R. (2002). "Effects of long-term supplementation with whey proteins on plasma glutathione levels of HIV-infected patients". European Journal of Nutrition 41 (1): 12–8. doi:10.1007/s003940200001. PMID 11990003.
- ^ Bounous, G; Baruchel, S; Falutz, J; Gold, P (1993). "Whey proteins as a food supplement in HIV-seropositive individuals". Clinical and investigative medicine. Medecine clinique et experimentale 16 (3): 204–9. PMID 8365048.
- ^ Bounous, G; Gold, P (1991). "The biological activity of undenatured dietary whey proteins: role of glutathione". Clinical and investigative medicine. Medecine clinique et experimentale 14 (4): 296–309. PMID 1782728.
- ^ Shay, Kate Petersen; Moreau, Régis F.; Smith, Eric J.; Smith, Anthony R.; Hagen, Tory M. (2009). "Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential". Biochimica et Biophysica Acta (BBA) - General Subjects 1790 (10): 1149–60. doi:10.1016/j.bbagen.2009.07.026. PMC 2756298. PMID 19664690. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2756298.
- ^ Busse, E; Zimmer, G; Schopohl, B; Kornhuber, B (1992). "Influence of alpha-lipoic acid on intracellular glutathione in vitro and in vivo". Arzneimittel-Forschung 42 (6): 829–31. PMID 1418040.
- ^ Barlowwalden, L; Reiter, R; Abe, M; Pablos, M; Menendezpelaez, A; Chen, L; Poeggeler, B (1995). "Melatonin stimulates brain glutathione peroxidase activity". Neurochemistry International 26 (5): 497–502. doi:10.1016/0197-0186(94)00154-M. PMID 7492947.
- ^ Nencini, C; Giorgi, G; Micheli, L (2007). "Protective effect of silymarin on oxidative stress in rat brain". Phytomedicine 14 (2–3): 129–35. doi:10.1016/j.phymed.2006.02.005. PMID 16638633.
- ^ Valenzuela, Alfonso; Aspillaga, MóNica; Vial, Soledad; Guerra, Ricardo (2007). "Selectivity of Silymarin on the Increase of the Glutathione Content in Different Tissues of the Rat". Planta Medica 55 (5): 420–2. doi:10.1055/s-2006-962056. PMID 2813578.
- ^ Enlander, Derek (2002). "CFS Treatment using Glutathione in Immunoprop". The CFS HandBook: 58–62.
- ^ Bounous, G; Molson, J (1999). "Competition for glutathione precursors between the immune system and the skeletal muscle: Pathogenesis of chronic fatigue syndrome". Med Hypotheses. 53(4) (oct): 347–9.
- ^ Richards, RS; Roberts, TK (2000). "Blood parameters indicative of oxidative stress are associated with symptom expression in chronic fatigue syndrome". Redox Rep. 1 (5): 35–41.
- ^ Glutathione: Information for Physicians: http://www.nutritionadvisor.com/glutathione.html
- ^ Benefits of Glutathione Enhancement in Disease or Stress: Pulmonary Disease http://www.fda.gov/ohrms/dockets/ac/00/slides/3652s1_05/tsld018.htm
- ^ Dröge, Wulf; Holm, Eggert (1997). "Role of cysteine and glutathione in HIV infection and other diseases associated with muscle wasting and immunological dysfunction". The FASEB journal : official publication of the Federation of American Societies for Experimental Biology 11 (13): 1077–89. PMID 9367343. http://www.fasebj.org/cgi/pmidlookup?view=long&pmid=9367343.
- ^ Herzenberg, L. A.; De Rosa, SC; Dubs, JG; Roederer, M; Anderson, MT; Ela, SW; Deresinski, SC; Herzenberg, LA (1997). "Glutathione deficiency is associated with impaired survival in HIV disease". Proceedings of the National Academy of Sciences 94 (5): 1967–72. doi:10.1073/pnas.94.5.1967. PMC 20026. PMID 9050888. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=20026.
- ^ Gawryluk, JW; Wang, JF; Andreazza, AC; Shao, L; Young, LT (2011). "Decreased levels of glutathione, the major brain antioxidant, in post-mortem prefrontal cortex from patients with psychiatric disorders". The international journal of neuropsychopharmacology / official scientific journal of the Collegium Internationale Neuropsychopharmacologicum (CINP) 14 (1): 123–30. doi:10.1017/S1461145710000805. PMID 20633320.
- ^ Park (2009). "The effects of N-acetyl cysteine, buthionine sulfoximine, diethyldithiocarbamate or 3-amino-1,2,4-triazole on antimycin A-treated Calu-6 lung cells in relation to cell growth, reactive oxygen species and glutathione". Oncology Reports: 385–91. doi:10.3892/or_00000449.
- ^ Chow, H.-H. S.; Hakim, I. A.; Vining, D. R.; Crowell, J. A.; Tome, M. E.; Ranger-Moore, J.; Cordova, C. A.; Mikhael, D. M. et al. (2007). "Modulation of Human Glutathione S-Transferases by Polyphenon E Intervention". Cancer Epidemiology Biomarkers & Prevention 16 (8): 1662–6. doi:10.1158/1055-9965.EPI-06-0830.
- ^ WebMD: Whey Protein May Prevent Prostate Cancer
- ^ Glutathione Information on MedicineNet.com (a WebMD feature)
- ^ Balendiran, Ganesaratnam K.; Dabur, Rajesh; Fraser, Deborah (2004). "The role of glutathione in cancer". Cell Biochemistry and Function 22 (6): 343–52. doi:10.1002/cbf.1149. PMID 15386533.
- ^ Pereira, C; Oliveira, CR (2000). "Oxidative glutamate toxicity involves mitochondrial dysfunction and perturbation of intracellular Ca2+ homeostasis". Neuroscience Research 37 (3): 227–36. doi:10.1016/S0168-0102(00)00124-3. PMID 10940457.
- ^ Meyer, Andreas J.; May, Mike J.; Fricker, Mark (2001). "Quantitative in vivo measurement of glutathione in Arabidopsis cells". The Plant Journal 27 (1): 67–78. doi:10.1046/j.1365-313x.2001.01071.x. PMID 11489184.
- ^ Meyer, Andreas J.; Brach, Thorsten; Marty, Laurent; Kreye, Susanne; Rouhier, Nicolas; Jacquot, Jean-Pierre; Hell, RüDiger (2007). "Redox-sensitive GFP inArabidopsis thalianais a quantitative biosensor for the redox potential of the cellular glutathione redox buffer". The Plant Journal 52 (5): 973–86. doi:10.1111/j.1365-313X.2007.03280.x. PMID 17892447.
- ^ Maulucci, Giuseppe; Labate, Valentina; Mele, Marina; Panieri, Emiliano; Arcovito, Giuseppe; Galeotti, Tommaso; Østergaard, H; Winther, JR et al. (2008). "High-resolution imaging of redox signaling in live cells through an oxidation-sensitive yellow fluorescent protein". Science Signaling 1 (43): pl3. doi:10.1126/scisignal.143pl3. PMID 18957692.
Related research
- Drevet, J (2006). "The antioxidant glutathione peroxidase family and spermatozoa: A complex story". Molecular and Cellular Endocrinology 250 (1–2): 70–9. doi:10.1016/j.mce.2005.12.027. PMID 16427183.
- The Role of Glutathione in Cell Defense.
- Wu, Guoyao; Fang, Yun-Zhong; Yang, Sheng; Lupton, Joanne R.; Turner, Nancy D. (2004). "Glutathione metabolism and its implications for health". The Journal of nutrition 134 (3): 489–92. PMID 14988435. http://jn.nutrition.org/cgi/content/full/134/3/489.
Antioxidants Acetyl-L-Carnitine (ALCAR) • Alpha-Lipoic Acid (ALA) • Ascorbic Acid (Vitamin C) • Carotenoids (Vitamin A) • Curcumin • Edaravone • Polyphenols • Glutathione • Hydroxytyrosol • L-Carnitine • Ladostigil • Melatonin • N-Acetylcysteine (NAC) • N-Acetylserotonin (NAS) • Oleocanthal • Oleuropein • Rasagiline • Resveratrol • Selegiline • Selenium • Tocopherols (Vitamin E) • Tocotrienols (Vitamin E) • Tyrosol • Ubiquinone (Coenzyme Q) • Uric AcidAntidotes (V03AB) Nervous system Barbiturate overdoseBemegride • EthamivanBenzodiazepine overdoseGHB overdoseCardiovascular Other Paracetamol toxicity (Acetaminophen)OtherPrednisolone/promethazine • oxidizing agent (potassium permanganate) • iodine-131 (Potassium iodide) • Methylthioninium chloride#Emetic Enzyme cofactors Active forms TPP / ThDP (B1) · FMN, FAD (B2) · NAD+, NADH, NADP+, NADPH (B3) · Coenzyme A (B5) · PLP / P5P (B6) · Biotin (B7) · THFA / H4FA, DHFA / H2FA, MTHF (B9) · AdoCbl, MeCbl (B12) · Ascorbic Acid (C) · Phylloquinone (K1), Menaquinone (K2) · Coenzyme F420ATP · CTP · SAMe · PAPS · GSH · Coenzyme B · Cofactor F430 · Coenzyme M · Coenzyme Q · Heme / Haem (A, B, C, O) · Lipoic Acid · Methanofuran · Molybdopterin/Molybdenum cofactor · PQQ · THB / BH4 · THMPT / H4MPTBase forms M: NUT
cof, enz, met
noco, nuvi, sysi/epon, met
drug(A8/11/12)
Categories:- Thiols
- Peptides
- Antioxidants

Wikimedia Foundation. 2010.