- Pseudomonas aeruginosa
-
Pseudomonas aeruginosa 
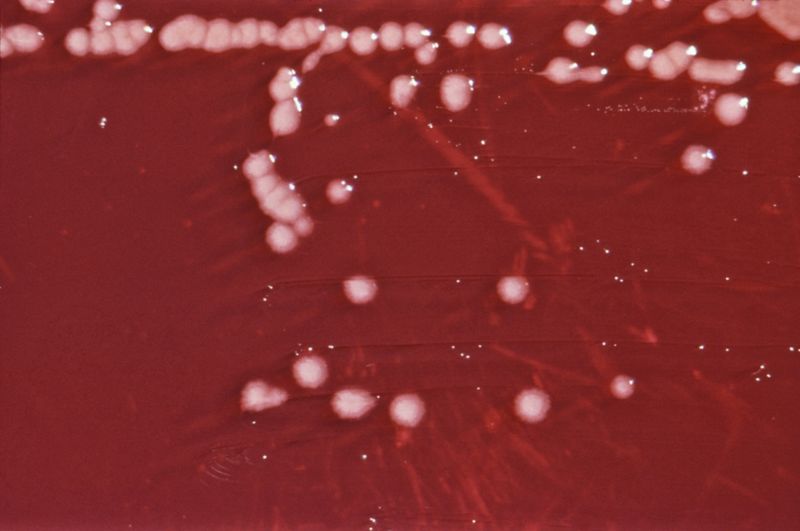
P. aeruginosa on an XLD agar plate. Scientific classification Kingdom: Bacteria Phylum: Proteobacteria Class: Gamma Proteobacteria Order: Pseudomonadales Family: Pseudomonadaceae Genus: Pseudomonas Species: P. aeruginosa Binomial name Pseudomonas aeruginosa
(Schröter 1872)
Migula 1900Type strain ATCC 10145
CCUG 551
CFBP 2466
CIP 100720
DSM 50071
JCM 5962
LMG 1242
NBRC 12689
NCCB 76039
NCIMB 8295
NCTC 10332
NRRL B-771
VKM B-588Synonyms Bacterium aeruginosum Schroeter 1872
Bacterium aeruginosum Cohn 1872
Micrococcus pyocyaneus Zopf 1884
Bacillus aeruginosus (Schroeter 1872) Trevisan 1885
Bacillus pyocyaneus (Zopf 1884) Flügge 1886
Pseudomonas pyocyanea (Zopf 1884) Migula 1895
Bacterium pyocyaneum (Zopf 1884) Lehmann and Neumann 1896
Pseudomonas polycolor Clara 1930
Pseudomonas vendrelli nomen nudum 1938Pseudomonas aeruginosa is a common bacterium that can cause disease in animals, including humans. It is found in soil, water, skin flora, and most man-made environments throughout the world. It thrives not only in normal atmospheres, but also in hypoxic atmospheres, and has, thus, colonized many natural and artificial environments. It uses a wide range of organic material for food; in animals, the versatility enables the organism to infect damaged tissues or those with reduced immunity. The symptoms of such infections are generalized inflammation and sepsis. If such colonizations occur in critical body organs, such as the lungs, the urinary tract, and kidneys, the results can be fatal.[1] Because it thrives on most surfaces, this bacterium is also found on and in medical equipment, including catheters, causing cross-infections in hospitals and clinics. It is implicated in hot-tub rash. It is also able to decompose hydrocarbons and has been used to break down tarballs and oil from oil spills.[2][3]
Contents
Identification
It is a Gram-negative, aerobic, rod-shaped bacterium with unipolar motility.[4] An opportunistic human pathogen, P. aeruginosa is also an opportunistic pathogen of plants.[5] P. aeruginosa is the type species of the genus Pseudomonas (Migula).[6]
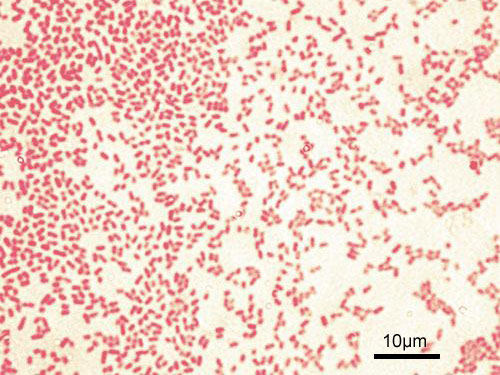
 Gram-stained Pseudomonas aeruginosa bacteria (pink-red rods)
Gram-stained Pseudomonas aeruginosa bacteria (pink-red rods)
P. aeruginosa secretes a variety of pigments, including pyocyanin (blue-green), pyoverdine (yellow-green and fluorescent), and pyorubin (red-brown). King, Ward, and Raney developed Pseudomonas Agar P (King A medium) for enhancing pyocyanin and pyorubin production, and Pseudomonas Agar F (King B medium) for enhancing fluorescein production.[7]P. aeruginosa is often preliminarily identified by its pearlescent appearance and grape-like or tortilla-like odor in vitro. Definitive clinical identification of P. aeruginosa often includes identifying the production of both pyocyanin and fluorescein, as well as its ability to grow at 42°C. P. aeruginosa is capable of growth in diesel and jet fuel, where it is known as a hydrocarbon-using microorganism (or "HUM bug"), causing microbial corrosion.[3] It creates dark, gellish mats sometimes improperly called "algae" because of their appearance.[citation needed]
Although classified as an aerobic organism, P. aeruginosa is considered by many as a facultative anaerobe, as it is well adapted to proliferate in conditions of partial or total oxygen depletion. This organism can achieve anaerobic growth with nitrate as a terminal electron acceptor, and, in its absence, it is also able to ferment arginine by substrate-level phosphorylation.[8][9] Adaptation to microaerobic or anaerobic environments is essential for certain lifestyles of P. aeruginosa, for example, during lung infection in cystic fibrosis patients, where thick layers of lung mucus and alginate surrounding mucoid bacterial cells can limit the diffusion of oxygen.[10][11][12][13]
Nomenclature
- The word Pseudomonas means "false unit", from the Greek pseudo (Greek: ψευδο, false) and monas (Latin: monas, from Greek: μονος, a single unit). The stem word mon was used early in the history of microbiology to refer to germs, e.g., Kingdom Monera.
- The species name aeruginosa is a Latin word meaning "copper rust", as seen with the oxidized copper patina on the Statue of Liberty. This also describes the blue-green bacterial pigment seen in laboratory cultures of the species. This blue-green pigment is a combination of two metabolites of P. aeruginosa, pyocyanin (blue) and pyoverdine (green), which impart the blue-green characteristic color of cultures. Pyocyanin biosynthesis is regulated by quorum sensing as in the biofilms associated with colonization of the lungs in cystic fibrosis patients. Another assertion is that the word may be derived from the Greek prefix ae- meaning "old or aged", and the suffix ruginosa means wrinkled or bumpy.[14]
- The derivations of pyocyanin and pyoverdine are of the Greek, with pyo-, meaning "pus", cyanin, meaning "blue", and verdine, meaning "green". Pyoverdine in the absence of pyocyanin is a fluorescent-yellow color.
Genomic diversity
The G+C-rich Pseudomonas aeruginosa chromosome consists of a conserved core and a variable accessory part. The core genomes of P. aeruginosa strains are largely collinear, exhibit a low rate of sequence polymorphism, and contain few loci of high sequence diversity, the most notable ones being the pyoverdine locus, the flagellar regulon, pilA, and the O-antigen biosynthesis locus. Variable segments are scattered throughout the genome, of which about one-third are immediately adjacent to tRNA or tmRNA genes. The three known hot spots of genomic diversity are caused by the integration of genomic islands of the pKLC102/PAGI-2 family into tRNALys or tRNAGly genes. The individual islands differ in their repertoire of metabolic genes, but share a set of syntenic genes that confer their horizontal spread to other clones and species. Colonization of atypical disease habitats predisposes to deletions, genome rearrangements, and accumulation of loss-of-function mutations in the P. aeruginosa chromosome. The P. aeruginosa population is characterized by a few dominant clones widespread in disease and environmental habitats. The genome is made up of clone-typical segments in core and accessory genome and of blocks in the core genome with unrestricted gene flow in the population.[15]
Cell-surface polysaccharides
Cell-surface polysaccharides play diverse roles in the bacterial "lifestyle". They serve as a barrier between the cell wall and the environment, mediate host-pathogen interactions, and form structural components of biofilms. These polysaccharides are synthesized from nucleotide-activated precursors, and, in most cases, all the enzymes necessary for biosynthesis, assembly, and transport of the completed polymer are encoded by genes organized in dedicated clusters within the genome of the organism. Lipopolysaccharide is one of the most important cell-surface polysaccharides, as it plays a key structural role in outer membrane integrity, as well as being an important mediator of host-pathogen interactions. The genetics for the biosynthesis of the so-called A-band (homopolymeric) and B-band (heteropolymeric) O antigens have been clearly defined, and much progress has been made toward understanding the biochemical pathways of their biosynthesis. The exopolysaccharide alginate is a linear copolymer of β-1,4-linked D-mannuronic acid and L-glucuronic acid residues, and is responsible for the mucoid phenotype of late-stage cystic fibrosis disease. The pel and psl loci are two recently-discovered gene clusters, which also encode exopolysaccharides found to be important for biofilm formation. A rhamnolipid is a biosurfactant whose production is tightly regulated at the transcriptional level, but the precise role that it plays in disease is not well understood at present. Protein glycosylation, in particular of pilin and flagellin, is a recent focus of research by several groups, and it has been shown to be important for adhesion and invasion during bacterial infection.[15]
Pathogenesis
 Phagocytosis of P. aeruginosa by neutrophil in patient with bloodstream infection (Gram stain)
Phagocytosis of P. aeruginosa by neutrophil in patient with bloodstream infection (Gram stain)An opportunistic, nosocomial pathogen of immunocompromised individuals, P. aeruginosa typically infects the pulmonary tract, urinary tract, burns, wounds, and also causes other blood infections.[16]
Hospital infections
Hospital infections Details and common associations High-risk groups Pneumonia Diffuse bronchopneumonia Cystic fibrosis patients Septic shock Associated with skin lesion ecthyma gangerenosum Neutropenic patients Urinary tract infection Urinary tract catheterization Gastrointestinal infection Necrotising enterocolitis (NEC) NEC, especially in premature infants and neutropenic cancer patients Skin and soft tissue infections Hemorrhage and necrosis Burns victims and patients with wound infections It is the most common cause of infections of burn injuries and of the external ear (otitis externa), and is the most frequent colonizer of medical devices (e.g., catheters). Pseudomonas can, in rare circumstances, cause community-acquired pneumonias,[17] as well as ventilator-associated pneumonias, being one of the most common agents isolated in several studies.[18] Pyocyanin is a virulence factor of the bacteria and has been known to cause death in C. elegans by oxidative stress. However, research indicates that salicylic acid can inhibit pyocyanin production.[19] One in ten hospital-acquired infections are from Pseudomonas. Cystic fibrosis patients are also predisposed to P. aeruginosa infection of the lungs. P. aeruginosa may also be a common cause of "hot-tub rash" (dermatitis), caused by lack of proper, periodic attention to water quality. The most common cause of burn infections is P. aeruginosa. Pseudomonas is also a common cause of postoperative infection in radial keratotomy surgery patients. The organism is also associated with the skin lesion ecthyma gangrenosum. Pseudomonas aeruginosa is frequently associated with osteomyelitis involving puncture wounds of the foot, believed to result from direct inoculation with P. aeruginosa via the foam padding found in tennis shoes.
Toxins
P. aeruginosa uses the virulence factor exotoxin A to ADP-ribosylate eukaryotic elongation factor 2 in the host cell, much as the diphtheria toxin does. Without elongation factor 2, eukaryotic cells cannot synthesize proteins and necrose. The release of intracellular contents induces an immunologic response in immunocompetent patients. In addition Pseudomonas aeruginosa uses an exoenzyme, ExoU, which degrades the plasma membrane of eukaryotic cells, leading to lysis.
Triggers
With low phosphate levels, P. aeruginosa has been found to activate from benign symbiont to express lethal toxins inside the intestinal tract and severely damage or kill the host, which can be mitigated by providing excess phosphate instead of antibiotics.[20]
Plants and invertebrates
In higher plants, P. aeruginosa induces symptoms of soft rot, for example in Arabidopsis thaliana (Thale cress)[21] and Lactuca sativa (lettuce).[22][23] It is also pathogenic to invertebrate animals, including the nematode Caenorhabditis elegans,[24][25] the fruit fly Drosophila[26] and the moth Galleria mellonella.[27] The associations of virulence factors are the same for plant and animal infections.[22][28]
Quorum sensing
Regulation of gene expression can occur through cell-cell communication or quorum sensing (QS) via the production of small molecules called autoinducers. QS is known to control expression of a number of virulence factors. Another form of gene regulation that allows the bacteria to rapidly adapt to surrounding changes is through environmental signaling. Recent studies have discovered anaerobiosis can significantly impact the major regulatory circuit of QS. This important link between QS and anaerobiosis has a significant impact on production of virulence factors of this organism.[15] Garlic experimentally blocks quorum sensing in P. aeruginosa.[29]
Biofilms and treatment resistance
Biofilms of P. aeruginosa can cause chronic opportunistic infections, which are a serious problem for medical care in industrialized societies, especially for immunocompromised patients and the elderly. They often cannot be treated effectively with traditional antibiotic therapy. Biofilms seem to protect these bacteria from adverse environmental factors. P. aeruginosa can cause nosocomial infections and is considered a model organism for the study of antibiotic-resistant bacteria. Researchers consider it important to learn more about the molecular mechanisms that cause the switch from planktonic growth to a biofilm phenotype and about the role of interbacterial communication in treatment-resistant bacteria such as P. aeruginosa. This should contribute to better clinical management of chronically infected patients, and should lead to the development of new drugs.[15]
Diagnosis
 Production of pyocyanin, water-soluble green pigment of Pseudomonas aeruginosa (left tube)
Production of pyocyanin, water-soluble green pigment of Pseudomonas aeruginosa (left tube)Depending on the nature of infection, an appropriate specimen is collected and sent to a bacteriology laboratory for identification. As with most bacteriological specimens, a Gram stain is performed, which may show Gram-negative rods and/or white blood cells. P. aeruginosa produces colonies with a characteristic 'grape-like' odour on bacteriological media. In mixed cultures, it can be isolated as clear colonies on MacConkey agar (as it does not ferment lactose) which will test positive for oxidase. Confirmatory tests include production of the blue-green pigment pyocyanin on cetrimide agar and growth at 42°C. A TSI slant is often used to distinguish nonfermenting Pseudomonas species from enteric pathogens in faecal specimens.
Treatment
P. aeruginosa is frequently isolated from nonsterile sites (mouth swabs, sputum, etc.), and, under these circumstances, it often represents colonization and not infection. The isolation of P. aeruginosa from nonsterile specimens should, therefore, be interpreted cautiously, and the advice of a microbiologist or infectious diseases physician/pharmacist should be sought prior to starting treatment. Often no treatment is needed.
When P. aeruginosa is isolated from a sterile site (blood, bone, deep collections), it should be taken seriously, and almost always requires treatment.[citation needed]
P. aeruginosa is naturally resistant to a large range of antibiotics and may demonstrate additional resistance after unsuccessful treatment, in particular, through modification of a porin. It should usually be possible to guide treatment according to laboratory sensitivities, rather than choosing an antibiotic empirically. If antibiotics are started empirically, then every effort should be made to obtain cultures, and the choice of antibiotic used should be reviewed when the culture results are available.
Phage therapy against P. aeruginosa remains one of the most effective treatments, which can be combined with antibiotics, has no contraindications and minimal adverse effects. Phages are produced as sterile liquid, suitable for intake, applications etc. [30] Phage therapy against ear infections caused by P. aeruginosa was reported in the journal Clinical Otolaryngology in August 2009[31]
Antibiotics that have activity against P. aeruginosa may include:
- aminoglycosides (gentamicin, amikacin, tobramycin)
- quinolones (ciprofloxacin, levofloxacin, and moxifloxacin)
- cephalosporins (ceftazidime, cefepime, cefoperazone, cefpirome, but not cefuroxime, ceftriaxone, cefotaxime)
- antipseudomonal penicillins: ureidopenicillins and carboxypenicillins (piperacillin, ticarcillin: P. aeruginosa is intrinsically resistant to all other penicillins)
- carbapenems (meropenem, imipenem, doripenem, but not ertapenem)
- polymyxins (polymyxin B and colistin)[32]
- monobactams (aztreonam)
These antibiotics must all be given by injection, with the exceptions of fluoroquinolones and aerosolized tobramycin. For this reason, in some hospitals, fluoroquinolone use is severely restricted to avoid the development of resistant strains of P. aeruginosa. In the rare occasions where infection is superficial and limited (for example, ear infections or nail infections), topical gentamicin or colistin may be used.
Antibiotic resistance
One of the most worrisome characteristics of P. aeruginosa is its low antibiotic susceptibility, which is attributable to a concerted action of multidrug efflux pumps with chromosomally-encoded antibiotic resistance genes (e.g., mexAB, mexXY etc.[33]) and the low permeability of the bacterial cellular envelopes. In addition to this intrinsic resistance, P. aeruginosa easily develops acquired resistance either by mutation in chromosomally-encoded genes or by the horizontal gene transfer of antibiotic resistance determinants. Development of multidrug resistance by P. aeruginosa isolates requires several different genetic events, including acquisition of different mutations and/or horizontal transfer of antibiotic resistance genes. Hypermutation favours the selection of mutation-driven antibiotic resistance in P. aeruginosa strains producing chronic infections, whereas the clustering of several different antibiotic resistance genes in integrons favors the concerted acquisition of antibiotic resistance determinants. Some recent studies have shown that phenotypic resistance associated to biofilm formation or to the emergence of small-colony variants may be important in the response of P. aeruginosa populations to antibiotics treatment.[15]
Phosphate trigger
Phosphate has been implicated in pathogenesis of P. aeruginosa, which is normally benign. Phosphate is required by the bacteria for normal functioning, and has been shown in experiments on two very different organisms to turn on its host.[20]
Prevention
Honey may reduce colonization of many pathogens, including P. aeruginosa.[34] Probiotic prophylaxis may prevent colonization and delay onset of pseudomonas infection in an ICU setting.[35] Immunoprophylaxis against pseudomonas is being investigated.[36]
See also
References
- ^ Balcht, Aldona & Smith, Raymond (1994). Pseudomonas Aeruginosa: Infections and Treatment. Informa Health Care. pp. 83–84. ISBN 0-8247-9210-6.
- ^ A. Y. Itah and J. P. Essien (2005). "Growth Profile and Hydrocarbonoclastic Potential of Microorganisms Isolated from Tarballs in the Bight of Bonny, Nigeria". World Journal of Microbiology and Biotechnology 21 (6–7): 1317–1322. doi:10.1007/s11274-004-6694-z.
- ^ a b AVI Biopharma (2007-01-18). "Antisense antibacterial method and compound". World Intellectual Property Organization. http://www.wipo.int/pctdb/en/wo.jsp?IA=US2006027522&DISPLAY=DESC. Retrieved 2008-10-18.
- ^ Ryan KJ, Ray CG (editors) (2004). Sherris Medical Microbiology (4th ed.). McGraw Hill. ISBN 0-8385-8529-9.
- ^ Iglewski BH (1996). Pseudomonas. In: Baron's Medical Microbiology (Baron S et al., eds.) (4th ed.). Univ of Texas Medical Branch. ISBN 0-9631172-1-1.
- ^ Anzai; Kim, H; Park, JY; Wakabayashi, H; Oyaizu, H (2000, Jul). "Phylogenetic affiliation of the pseudomonads based on 16S rRNA sequence". Int J Syst Evol Microbiol 50 (Pt 4): 1563–89. doi:10.1099/00207713-50-4-1563. PMID 10939664.
- ^ King EO, Ward MK, Raney DE (1954). "Two simple media for the demonstration of pyocyanin and fluorescein". J Lab Clin Med 44 (2): 301–7. PMID 13184240.
- ^ Palmer KL, Brown SA, Whiteley M (2007). "Membrane-bound nitrate reductase is required for anaerobic growth in cystic fibrosis sputum". J. Bacteriol. 189 (12): 4449–4455. doi:10.1128/JB.00162-07. PMC 1913347. PMID 17400735. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1913347.
- ^ Vander Wauven C, Piérard A, Kley-Raymann M, Haas D (1984). "Pseudomonas aeruginosa mutants affected in anaerobic growth on arginine: evidence for a four-gene cluster encoding the arginine deiminase pathway". J. Bacteriol. 160 (3): 928–34. PMC 215798. PMID 6438064. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=215798.
- ^ Hassett DJ (1996). "Anaerobic production of alginate by Pseudomonas aeruginosa: alginate restricts diffusion of oxygen". J. Bacteriol. 178 (24): 7322–5. PMC 178651. PMID 8955420. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=178651.
- ^ Worlitzsch D; Tarran R; Ulrich M; Schwab, Ute; Cekici, Aynur; Meyer, Keith C.; Birrer, Peter; Bellon, Gabriel et al. (2002). "Effects of reduced mucus oxygen concentration in airway Pseudomonas infections of cystic fibrosis patients". J. Clin. Invest. 109 (3): 317–325. doi:10.1172/JCI13870. PMC 150856. PMID 11827991. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=150856.
- ^ Cooper M, Tavankar GR, Williams HD (2003). "Regulation of expression of the cyanide-insensitive terminal oxidase in Pseudomonas aeruginosa". Microbiology 149 (Pt 5): 1275–1284. doi:10.1099/mic.0.26017-0. PMID 12724389.
- ^ Williams HD, Zlosnik JE, Ryall B (2007). "Oxygen, cyanide and energy generation in the cystic fibrosis pathogen Pseudomonas aeruginosa". Adv. Microb. Physiol. 52: 1–71. doi:10.1016/S0065-2911(06)52001-6. PMID 17027370.
- ^ Brown, RW (1956). Composition of Scientific Words. Smithsonian Institutional Press. ISBN 0-87474-286-2.
- ^ a b c d e Cornelis P (editor). (2008). Pseudomonas: Genomics and Molecular Biology (1st ed.). Caister Academic Press. ISBN 1904455190. http://www.horizonpress.com/pseudo.
- ^ Todar's Online Textbook of Bacteriology. Textbookofbacteriology.net (2004-06-04). Retrieved on 2011-10-09.
- ^ Fine MJ; Smith MA; Carson CA; Mutha, SS; Sankey, SS; Weissfeld, LA; Kapoor, WN (1996). "Prognosis and outcomes of patients with community-acquired pneumonia. A meta-analysis". JAMA 275 (2): 134–141. doi:10.1001/jama.275.2.134. PMID 8531309.
- ^ Diekema DJ; Pfaller MA; Jones RN; Doern, GV; Winokur, PL; Gales, AC; Sader, HS; Kugler, K et al. (1999). "Survey of bloodstream infections due to Gram-negative bacilli: frequency of occurrence and antimicrobial susceptibility of isolates collected in the United States, Canada, and Latin America for the SENTRY Antimicrobial Surveillance Program, 1997". Clin. Infect. Dis. 29 (3): 595–607. doi:10.1086/598640. PMID 10530454.
- ^ Prithiviraj B, Bais H, Weir T, Suresh B, Najarro E, Dayakar B, Schweizer H, Vivanco J (2005). "Down regulation of virulence factors of Pseudomonas aeruginosa by salicylic acid attenuates its virulence on Arabidopsis thaliana and Caenorhabditis elegans". Infect Immun 73 (9): 5319–5328. doi:10.1128/IAI.73.9.5319-5328.2005. PMC 1231131. PMID 16113247. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1231131.
- ^ a b "Research could lead to new non-antibiotic drugs to counter hospital infections" (Press release). University of Chicago Medical Center. 2009-04-14. http://news.uchicago.edu/news.php?asset_id=1589. Retrieved 2010-01-18.
- ^ Walker TS; Bais HP; Déziel E; Schweizer, HP; Rahme, LG; Fall, R; Vivanco, JM (2004). "Pseudomonas aeruginosa-plant root interactions. Pathogenicity, biofilm formation, and root exudation". Plant Physiol. 134 (1): 320–331. doi:10.1104/pp.103.027888. PMC 316311. PMID 14701912. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=316311.
- ^ a b Rahme LG, Stevens EJ, Wolfort SF, Shao J, Tompkins RG, Ausubel FM (1995). "Common virulence factors for bacterial pathogenicity in plants and animals". Science 268 (5219): 1899–1902. doi:10.1126/science.7604262. PMID 7604262.
- ^ Rahme LG; Tan MW; Le L; Wong, SM; Tompkins, RG; Calderwood, SB; Ausubel, FM (1997). "Use of model plant hosts to identify Pseudomonas aeruginosa virulence factors". Proc. Natl. Acad. Sci. U.S.A. 94 (24): 13245–13250. doi:10.1073/pnas.94.24.13245. PMC 24294. PMID 9371831. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=24294.
- ^ Mahajan-Miklos S, Tan MW, Rahme LG, Ausubel FM (1999). "Molecular mechanisms of bacterial virulence elucidated using a Pseudomonas aeruginosa-Caenorhabditis elegans pathogenesis model". Cell 96 (1): 47–56. doi:10.1016/S0092-8674(00)80958-7. PMID 9989496.
- ^ Martínez C, Pons E, Prats G, León J (2004). "Salicylic acid regulates flowering time and links defence responses and reproductive development". Plant J. 37 (2): 209–17. doi:10.1046/j.1365-313X.2003.01954.x. PMID 14690505.
- ^ D'Argenio DA, Gallagher LA, Berg CA, Manoil C (2001). "Drosophila as a model host for Pseudomonas aeruginosa infection". J. Bacteriol. 183 (4): 1466–1471. doi:10.1128/JB.183.4.1466-1471.2001. PMC 95024. PMID 11157963. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=95024.
- ^ Miyata S, Casey M, Frank DW, Ausubel FM, Drenkard E (2003). "Use of the Galleria mellonella caterpillar as a model host to study the role of the type III secretion system in Pseudomonas aeruginosa pathogenesis". Infect. Immun. 71 (5): 2404–2413. doi:10.1128/IAI.71.5.2404-2413.2003. PMC 153283. PMID 12704110. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=153283.
- ^ Rahme LG; Ausubel FM; Cao H; Drenkard, E; Goumnerov, BC; Lau, GW; Mahajan-Miklos, S; Plotnikova, J et al. (2000). "Plants and animals share functionally common bacterial virulence factors". Proc. Natl. Acad. Sci. U.S.A. 97 (16): 8815–8821. doi:10.1073/pnas.97.16.8815. PMC 34017. PMID 10922040. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=34017.
- ^ Thomas Bjarnsholt,1 Peter Østrup Jensen,2 Thomas B. Rasmussen,1 Lars Christophersen,2 Henrik Calum,2 Morten Hentzer,3 Hans-Petter Hougen,4 Jørgen Rygaard,5 Claus Moser,2 Leo Eberl,6 Niels Høiby2 and Michael Givskov1 (2005). "Garlic blocks quorum sensing and promotes rapid clearing of pulmonary Pseudomonas aeruginosa infections". Microbiology 151 (4): 3873–3880. doi:10.1099/mic.0.27955-0. PMID 16339933.
- ^ A. Sulakvelidze, Z. Alavidze and J.G. Morris, Jr. (2001). "Bacteriophage therapy". Antimicrobial Agents and Chemotherapy 45 (3): 649–659. doi:10.1128/AAC.45.3.649-659.2001.
- ^ Wright, A.; Hawkins, C.; Anggård, E.; Harper, D. (2009). "A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; a preliminary report of efficacy". Clinical Otolaryngology 34 (4): 349–357. doi:10.1111/j.1749-4486.2009.01973.x. PMID 19673983.
- ^ Hachem RY; Chemaly RF; Ahmar CA; Jiang, Y.; Boktour, M. R.; Rjaili, G. A.; Bodey, G. P.; Raad, I. I. (2007). "Colistin is effective in treatment of infections caused by multidrug-resistant Pseudomonas aeruginosa in cancer patients". Antimicrob. Agents Chemother. 51 (6): 1905–1911. doi:10.1128/AAC.01015-06. PMC 1891378. PMID 17387153. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1891378.
- ^ Poole, K. (2004). "Efflux-mediated multiresistance in Gram-negative bacteria". Clinical Microbiology and Infection 10 (1): 12–26. doi:10.1111/j.1469-0691.2004.00763.x. PMID 14706082.
- ^ Kwakman PH, Van den Akker JP, Güçlü A, Aslami H, Binnekade JM, de Boer L, Boszhard L, Paulus F, Middelhoek P, te Velde AA, Vandenbroucke-Grauls CM, Schultz MJ, Zaat SA. (2008). "Medical-grade honey kills antibiotic-resistant bacteria in vitro and eradicates skin colonization". Clin Infect Dis. 46 (11): 1677–82. PMID 18433338. http://www.bfactory.nl/artikelen/2008%20Kwakman%20et%20al%20in%20CID.pdf.
- ^ Forestier C, Guelon D, Cluytens V, Gillart T, Sirot J, de Champs C. (2008). "Oral probiotic and prevention of Pseudomonas aeruginosa infections: a randomized, double-blind, placebo-controlled pilot study in ICU-patients". Crit Care 12 (3): R69. PMID 18489775.
- ^ Döring G, Pier GB (2008). "Vaccines and immunotherapy against Pseudomonas aeruginosa". Vaccine 26 (8): 1011–24. doi:10.1016/j.vaccine.2007.12.007. PMID 18242792.
Breidenstein EB, de la Fuente-Núñez C, Hancock RE. Pseudomonas aeruginosa: all roads lead to resistance. Trends Microbiol. 2011;19:419-26.
Categories:- Disinfectants
- Gram negative bacteria
- Pseudomonadales


Wikimedia Foundation. 2010.