- Lung
-
"Lunged" redirects here. For other uses, see Lunge.
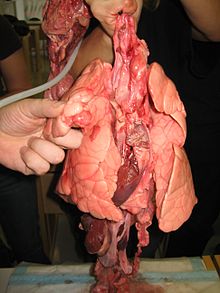 The lungs of a pig
The lungs of a pig

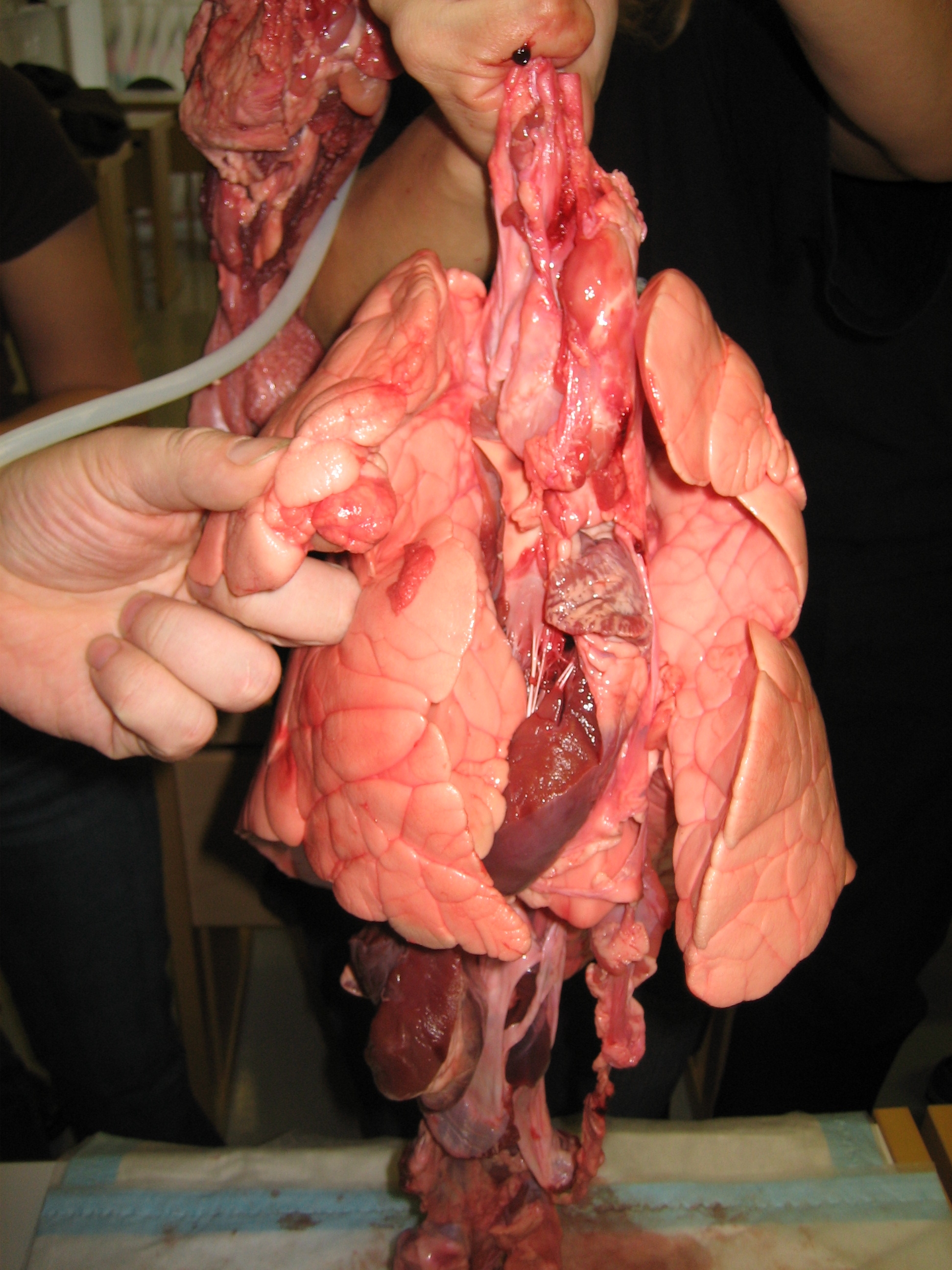
 Air enters and leaves the lungs via a conduit of cartilaginous passageways—the bronchi and bronchioles. In this image, lung tissue has been dissected away to reveal the bronchioles[1]
Air enters and leaves the lungs via a conduit of cartilaginous passageways—the bronchi and bronchioles. In this image, lung tissue has been dissected away to reveal the bronchioles[1]The lung is the essential respiration organ in many air-breathing animals, including most tetrapods, a few fish and a few snails. In mammals and the more complex life forms, the two lungs are located near the backbone on either side of the heart. Their principal function is to transport oxygen from the atmosphere into the bloodstream, and to release carbon dioxide from the bloodstream into the atmosphere. This exchange of gases is accomplished in the mosaic of specialized cells that form millions of tiny, exceptionally thin-walled air sacs called alveoli.
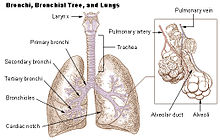
To completely explain the anatomy of the lungs, it is necessary to discuss the passage of air through the mouth to the alveoli. Once air progresses through the mouth or nose, it travels through the oropharynx, nasopharynx, the larynx, the trachea, and a progressively subdividing system of bronchi and bronchioles until it finally reaches the alveoli where the gas exchange of carbon dioxide and oxygen takes place.[2]
The drawing and expulsion of air (ventilation) is driven by muscular action; in early tetrapods, air was driven into the lungs by the pharyngeal muscles via buccal pumping, whereas in reptiles, birds and mammals a more complicated musculoskeletal system is used.
Medical terms related to the lung often begin with pulmo-, such as in the (adjectival form: pulmonary) or from the Latin pulmonarius ("of the lungs"), or with pneumo- (from Greek πνεύμων "lung").
Contents
Mammalian lungs
The lungs of mammals have a spongy and soft texture and are honeycombed with epithelium, having a much larger surface area in total than the outer surface area of the lung itself. The lungs of humans are a typical example of this type of lung.
Breathing is largely driven by the muscular diaphragm at the bottom of the thorax. Contraction of the diaphragm pulls the bottom of the cavity in which the lung is enclosed downward, increasing volume and thus decreasing pressure, causing air to flow into the airways. Air enters through the oral and nasal cavities; it flows through the pharynx, then the larynx and into the trachea, which branches out into the main bronchi and then subsequent divisions. During normal breathing, expiration is passive and no muscles are contracted (the diaphragm relaxes). The rib cage itself is also able to expand and contract to some degree, through the action of other respiratory and accessory respiratory muscles. As a result, air is transported into or expelled out of the lungs. This type of lung is known as a bellows lung as it resembles a blacksmith's bellows.[3]
Anatomy
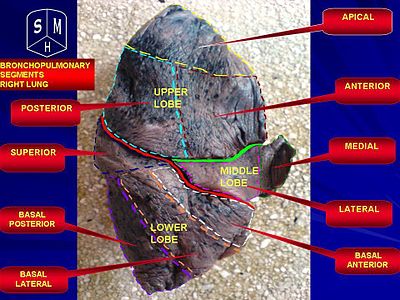 Human right lung
Human right lungIn humans, the trachea divides into the two main bronchi that enter the roots of the lungs. The bronchi continue to divide within the lung, and after multiple divisions, give rise to bronchioles. The bronchial tree continues branching until it reaches the level of terminal bronchioles, which lead to alveolar sacs. Alveolar sacs are made up of clusters of alveoli, like individual grapes within a bunch. The individual alveoli are tightly wrapped in blood vessels and it is here that gas exchange actually occurs. Deoxygenated blood from the heart is pumped through the pulmonary artery to the lungs, where oxygen diffuses into blood and is exchanged for carbon dioxide in the hemoglobin of the erythrocytes. The oxygen-rich blood returns to the heart via the pulmonary veins to be pumped back into systemic circulation.
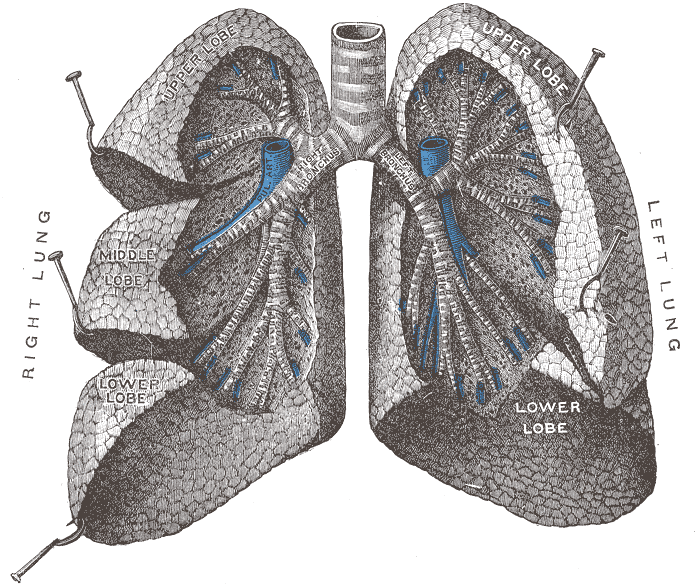
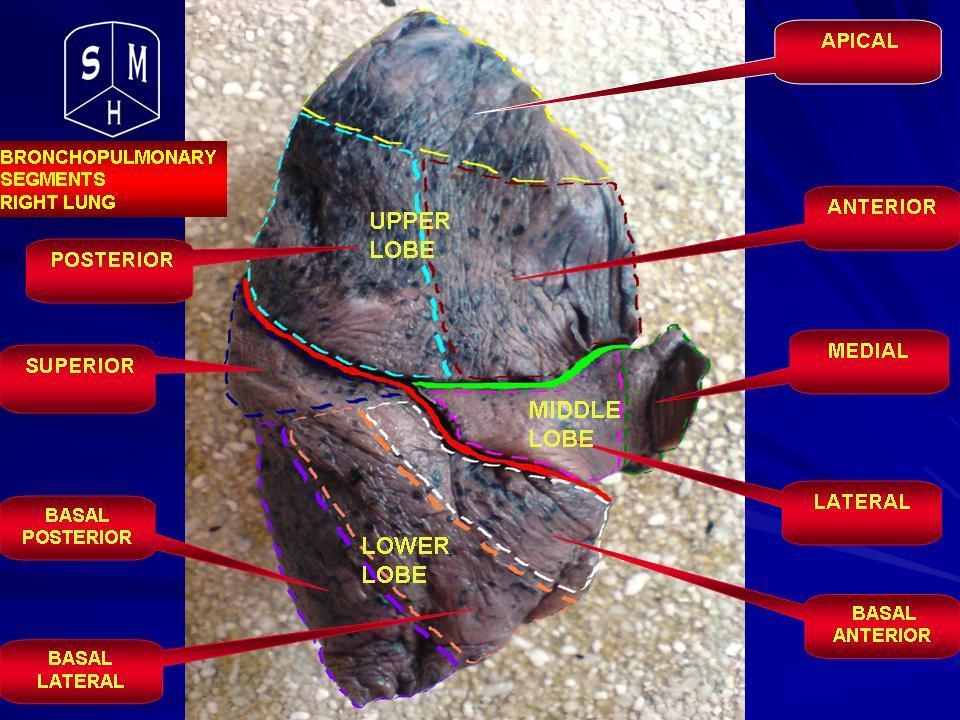
Human lungs are located in two cavities on either side of the heart. Though similar in appearance, the two are not identical. Both are separated into lobes by fissures, with three lobes on the right and two on the left. The lobes are further divided into segments and then into lobules, hexagonal divisions of the lungs that are the smallest subdivision visible to the naked eye. The connective tissue that divides lobules is often blackened in smokers. The medial border of the right lung is nearly vertical, while the left lung contains a cardiac notch. The cardiac notch is a concave impression molded to accommodate the shape of the heart.
Each lobe is surrounded by a pleural cavity, which consists of two pleurae. The parietal pleura lies against the rib cage, and the visceral pleura lies on the surface of the lungs. In between the pleura is pleural fluid. The pleural cavity helps the lubricate the lungs, as well as providing surface tension to keep the lung surface in contact with the rib cage.
Lungs are to a certain extent 'overbuilt' and have a tremendous reserve volume as compared to the oxygen exchange requirements when at rest. Such excess capacity is one of the reasons that individuals can smoke for years without having a noticeable decrease in lung function while still or moving slowly; in situations like these only a small portion of the lungs are actually perfused with blood for gas exchange. Destruction of too many alveoli over time leads to the condition emphysema, which is associated with extreme shortness of breath. As oxygen requirements increase due to exercise, a greater volume of the lungs is perfused, allowing the body to match its CO2/O2 exchange requirements. Additionally, due to the excess capacity, it is possible for humans to live with only one lung, with the one compensating for the other's loss.
The environment of the lung is very moist, which makes it hospitable for bacteria. Many respiratory illnesses are the result of bacterial or viral infection of the lungs. Inflammation of the lungs is known as pneumonia; inflammation of the pleura surrounding the lungs is known as pleurisy.
Vital capacity is the maximum volume of air that a person can exhale after maximum inhalation; it can be measured with a spirometer. In combination with other physiological measurements, the vital capacity can help make a diagnosis of underlying lung disease.
The lung parenchyma is strictly used to refer solely to alveolar tissue with respiratory bronchioles, alveolar ducts and terminal bronchioles.[4] However, it often includes any form of lung tissue, also including bronchioles, bronchi, blood vessels and lung interstitium.[4]
Non respiratory functions
In addition to their function in respiration, the lungs also:
- Alter the pH of blood by facilitating alterations in the partial pressure of carbon dioxide
- Filter out small blood clots formed in veins
- Filter out gas micro-bubbles occurring in the venous blood stream such as those created after scuba diving during decompression.[5]
- Influence the concentration of some biologic substances and drugs used in medicine in blood
- Convert angiotensin I to angiotensin II by the action of angiotensin-converting enzyme
- May serve as a layer of soft, shock-absorbent protection for the heart, which the lungs flank and nearly enclose.
- Immunoglobulin-A is secreted in the bronchial secretion and protects against respiratory infections.
- Maintain sterility by producing mucus containing antimicrobial compounds.[6] Mucus contains glycoproteins, e.g. mucins, lactoferrin,[7] lysozyme, lactoperoxidase.[8][9] We find also on the epithelium Dual oxidase 2[10][11][12] proteins generating hydrogen peroxide, useful for hypothiocyanite endogenous antimicrobial synthesis. Function not in place in cystic fibrosis patient lungs.[13][14]
- Ciliary escalator action is an important defence system against air-borne infection.The dust particles and bacteria in the inhaled air are caught in the mucous layer present at the mucosal surface of respiratory passages and are moved up towards pharynx by the rhythmic upward beating action of the cilia.
- Provide airflow for the creation of vocal sounds.
- Thermoregulation (panting)
Avian lungs
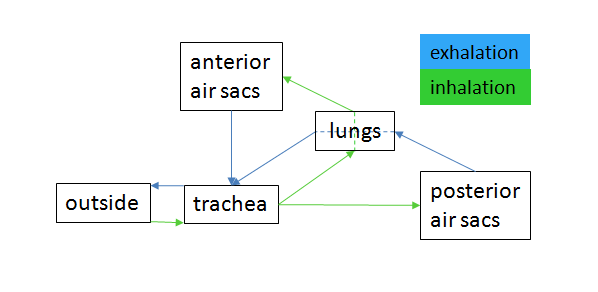
 Bird respiration air flow schematic
Bird respiration air flow schematicAvian lungs do not have alveoli as mammalian lungs do, they have Faveolar lungs. They contain millions of tiny passages known as para-bronchi, connected at both ends by the dorsobronchi. The airflow through the avian lung always travels in the same direction – posterior to anterior. This is in contrast to the mammalian system, in which the direction of airflow in the lung is tidal, reversing between inhalation and exhalation. By utilizing a unidirectional flow of air, avian lungs are able to extract a greater concentration of oxygen from inhaled air. Birds are thus equipped to fly at altitudes at which mammals would succumb to hypoxia. This also allows them to sustain a higher metabolic rate than an equivalent weight mammal.[15]
The lungs of birds are relatively small, but are connected to 8-9 air sacs that extend through much of the body, and are in turn connected to air spaces within the bones. The air sacs are smooth-walled, and do not themselves contribute much to respiration, but they do help to maintain the airflow through the lungs as air is forced through them by the movement of the ribs and flight muscles.[16]
Because of the complexity of the system, misunderstanding is common and it is incorrectly believed that it takes two breathing cycles for air to pass entirely through a bird's respiratory system. Air is not stored in either the posterior or anterior sacs between respiration cycles, air moves continuously from the posterior to the anterior of the lungs throughout respiration. This type of lung construction is called a circulatory lung, as distinct from the bellows lung possessed by other animals.[15]
Reptilian lungs
Reptilian lungs are typically ventilated by a combination of expansion and contraction of the ribs via axial muscles and buccal pumping. Crocodilians also rely on the hepatic piston method, in which the liver is pulled back by a muscle anchored to the pubic bone (part of the pelvis), which in turn pulls the bottom of the lungs backward, expanding them. Turtles, which are unable to move their ribs, instead use their forelimbs and pectoral girdle to force air in and out of the lungs.[16]
The lung of most reptiles has a single bronchus running down the centre, from which numerous branches reach out to individual pockets throughout the lungs. These pockets are similar to, but much larger and fewer in number than, mammalian alveoli, and give the lung a sponge-like texture. In tuataras, snakes, and some lizards, the lungs are simpler in structure, similar to that of typical amphibians.[16]
Snakes and limbless lizards typically possess only the right lung as a major respiratory organ; the left lung is greatly reduced, or even absent. Amphisbaenians, however, have the opposite arrangement, with a major left lung, and a reduced or absent right lung.[16]
Amphibian lungs
The lungs of most frogs and other amphibians are simple balloon-like structures, with gas exchange limited to the outer surface area of the lung. This is not a very efficient arrangement, but amphibians have low metabolic demands and also frequently supplement their oxygen supply by diffusion across the moist outer skin of their bodies. Unlike mammals, which use a breathing system driven by negative pressure, amphibians employ positive pressure.[16] The majority of salamander species are lungless salamanders, which respirate through their skin and tissues lining their mouth. The only other known lungless tetrapods are also amphibians; the Bornean Flat-headed Frog (Barbourula kalimantanensis) and Atretochoana eiselti, a caecilian.
The lungs of amphibians typically have a few narrow septa of soft tissue around the outer walls, increasing the respiratory surface area and giving the lung a honey-comb appearance. In some salamanders even these are lacking, and the lung has a smooth wall. In caecilians, as in snakes, only the right lung attains any size or development.[16]
Lungfish
The lungs of lungfish are similar to those of amphibians, with few, if any, internal septa. In Polypterus and the Australian lungfish, there is only a single lung, albeit divided into two lobes in the former case. Other lungfish, however, have two lungs, which are located in the upper part of the body, with the connecting duct curving round and above the esophagus. The blood supply also twists around the esophagus, suggesting that the lungs originally evolved in the ventral part of the body, as in other vertebrates.[16]
Invertebrate lungs
Some invertebrates have "lungs" that serve a similar respiratory purpose as, but are not evolutionarily related to, vertebrate lungs. Some arachnids have structures called "book lungs" used for atmospheric gas exchange. The Coconut crab uses structures called Branchiostegal lungs to breathe air and indeed will drown in water, hence it breathes on land and holds its breath underwater. The Pulmonata are an order of snails and slugs that have developed "lungs".
Origins of the vertebrate lung
The lungs of today's terrestrial vertebrates and the gas bladders of today's fish are believed to have evolved from simple sacs (outpocketings) of the esophagus that allowed early fish to gulp air under oxygen-poor conditions.[17] These outpocketings first arose in the bony fish; in some of the ray-finned fish the sacs evolved into gas bladders, while in other ray-finned fish (such as the gar, bichir and amia) as well as the lobe-finned fish they evolved into lungs.[17] The lobe-finned fish gave rise to the land-based tetrapods. Thus, the lungs of vertebrates are homologous to the gas bladders of fish (but not to their gills). This is reflected by the fact that the lungs of a fetus also develop from an outpocketing of the esophagus and in the case of gas bladders, this connection to the gut continues to exist as the pneumatic duct in more "primitive" teleosts, and is lost in the higher orders. (This is an instance of correlation between ontogeny and phylogeny.) No known animals have both a gas bladder and lungs.
See also
- Alveolar-capillary barrier
- American Lung Association
- Asthma
- Borders of the lung
- Bronchitis
- Bronchus
- Cardiothoracic surgery
- Chronic obstructive pulmonary disease
- Drowning
- Dry drowning
- Emphysema
- Gills
- Left lung
- Liquid breathing
- Lung cancer
- Lung volumes
- Mechanical ventilation
- Pneumothorax
- Pneumonia
- Pulmonary contusion
- Pulmonology
- Right lung
- tuberculosis
Further reading
- Lung Function Fundamentals. http://www.anaesthetist.com/icu/organs/lung/lungfx.htm
- Dr D.R. Johnson: Introductory anatomy, respiratory system
- Franlink Institute Online: The Respiratory System
- Lungs 'best in late afternoon'
- Chronic Respiratory Disease - leading research and articles on respiratory disease.
- Avian lungs and respiration
Footnotes
- ^ a b Gray's Anatomy of the Human Body, 20th ed. 1918.
- ^ Wienberger, Cockrill, Mandel. Principles of Pulmonary Medicine. Elsevier Science.[verification needed]
- ^ Maton, Anthea; Jean Hopkins, Charles William McLaughlin, Susan Johnson, Maryanna Quon Warner, David LaHart, Jill D. Wright1 (1993). Human Biology and Health. Englewood Cliffs, New Jersey, USA: Prentice Hall. ISBN 0-13-981176-1. OCLC 32308337.[page needed]
- ^ a b medilexicon.com > Medical Dictionary - 'Parenchyma Of Lung' In turn citing: Stedman's Medical Dictionary. 2006
- ^ Wienke B.R. : "Decompression theory"[verification needed]
- ^ Travis SM, Conway BA, Zabner J, et al. (May 1999). "Activity of abundant antimicrobials of the human airway". American Journal of Respiratory Cell and Molecular Biology 20 (5): 872–9. PMID 10226057. http://ajrcmb.atsjournals.org/cgi/pmidlookup?view=long&pmid=10226057.
- ^ Rogan MP, Taggart CC, Greene CM, Murphy PG, O'Neill SJ, McElvaney NG (October 2004). "Loss of microbicidal activity and increased formation of biofilm due to decreased lactoferrin activity in patients with cystic fibrosis". The Journal of Infectious Diseases 190 (7): 1245–53. doi:10.1086/423821. PMID 15346334.
- ^ Wijkstrom-Frei C, El-Chemaly S, Ali-Rachedi R, et al. (August 2003). "Lactoperoxidase and human airway host defense". American Journal of Respiratory Cell and Molecular Biology 29 (2): 206–12. doi:10.1165/rcmb.2002-0152OC. PMID 12626341.
- ^ Conner GE, Salathe M, Forteza R (December 2002). "Lactoperoxidase and hydrogen peroxide metabolism in the airway". American Journal of Respiratory and Critical Care Medicine 166 (12 Pt 2): S57–61. doi:10.1164/rccm.2206018. PMID 12471090.
- ^ Fischer H (October 2009). "Mechanisms and Function of DUOX in Epithelia of the Lung". Antioxidants & Redox Signaling 11 (10): 2453–65. doi:10.1089/ARS.2009.2558. PMC 2823369. PMID 19358684. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2823369.
- ^ Rada B, Leto TL (2008). "Oxidative innate immune defenses by Nox/Duox family NADPH Oxidases". Contributions to Microbiology. Contributions to Microbiology 15: 164–87. doi:10.1159/000136357. ISBN 978-3-8055-8548-4. PMC 2776633. PMID 18511861. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2776633.
- ^ Rada B, Lekstrom K, Damian S, Dupuy C, Leto TL (October 2008). "The Pseudomonas toxin pyocyanin inhibits the Dual oxidase-based antimicrobial system as it imposes oxidative stress on airway epithelial cells". Journal of Immunology 181 (7): 4883–93. PMC 2776642. PMID 18802092. http://www.jimmunol.org/cgi/pmidlookup?view=long&pmid=18802092.
- ^ Moskwa P, Lorentzen D, Excoffon KJ, et al. (January 2007). "A Novel Host Defense System of Airways Is Defective in Cystic Fibrosis". American Journal of Respiratory and Critical Care Medicine 175 (2): 174–83. doi:10.1164/rccm.200607-1029OC. PMC 2720149. PMID 17082494. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2720149.
- ^ Conner GE, Wijkstrom-Frei C, Randell SH, Fernandez VE, Salathe M (January 2007). "The Lactoperoxidase System Links Anion Transport To Host Defense in Cystic Fibrosis". FEBS Letters 581 (2): 271–8. doi:10.1016/j.febslet.2006.12.025. PMC 1851694. PMID 17204267. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1851694.
- ^ a b Ritchson, G. "BIO 554/754 - Ornithology: Avian respiration". Department of Biological Sciences, Eastern Kentucky University. http://www.people.eku.edu/ritchisong/birdrespiration.html. Retrieved 2009-04-23.
- ^ a b c d e f g Romer, Alfred Sherwood; Parsons, Thomas S. (1977). The Vertebrate Body. Philadelphia, PA: Holt-Saunders International. pp. 330–334. ISBN 0-03-910284-X.
- ^ a b Colleen Farmer (1997). "Did lungs and the intracardiac shunt evolve to oxygenate the heart in vertebrates". Paleobiology. http://www.biology.utah.edu/farmer/publications%20pdf/1997%20Paleobiology23.pdf.
Human systems and organs TA 2–4:
MSBone (Carpus · Collar bone (clavicle) · Thigh bone (femur) · Fibula · Humerus · Mandible · Metacarpus · Metatarsus · Ossicles · Patella · Phalanges · Radius · Skull (cranium) · Tarsus · Tibia · Ulna · Rib · Vertebra · Pelvis · Sternum) · CartilageTA 5–11:
splanchnic/
viscusMouth (Salivary gland, Tongue) · upper GI (Oropharynx, Laryngopharynx, Esophagus, Stomach) · lower GI (Small intestine, Appendix, Colon, Rectum, Anus) · accessory (Liver, Biliary tract, Pancreas)TA 12–16 Blood
(Non-TA)General anatomy: systems and organs, regional anatomy, planes and lines, superficial axial anatomy, superficial anatomy of limbsCategories:- Lung
- Organs


Wikimedia Foundation. 2010.