- Frog
-
For other uses, see Frog (disambiguation).
Frogs
Temporal range: Triassic–present
Australian Green Tree Frog (Litoria caerulea) Scientific classification Kingdom: Animalia Phylum: Chordata Class: Amphibia Subclass: Lissamphibia Order: Anura
Merrem, 1820Suborders Archaeobatrachia
Mesobatrachia
Neobatrachia
-
List of Anuran families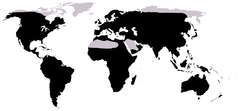
Native distribution of frogs (in black) Frogs are amphibians in the order Anura (meaning "tail-less", from Greek an-, without + oura, tail), formerly referred to as Salientia (Latin salere (salio), "to jump"). Most frogs are characterized by a short body, webbed digits (fingers or toes), protruding eyes and the absence of a tail. Frogs are widely known as exceptional jumpers, and many of the anatomical characteristics of frogs, particularly their long, powerful legs, are adaptations to improve jumping performance. Due to their permeable skin, frogs are often semi-aquatic or inhabit humid areas, but move easily on land. They typically lay their eggs in puddles, ponds or lakes, and their larvae, called tadpoles, have gills and develop in water. Adult frogs follow a carnivorous diet, mostly of arthropods, annelids and gastropods. Frogs are most noticeable by their call, which can be widely heard during the night or day, mainly in their mating season.
The distribution of frogs ranges from tropic to subarctic regions, but most species are found in tropical rainforests. Consisting of more than 5,000 species described, they are among the most diverse groups of vertebrates. However, populations of certain frog species are declining significantly.
A popular distinction is often made between frogs and toads on the basis of their appearance, but this has no taxonomic basis. (Members of the anuran family Bufonidae are called true toads, but many species from other families are also called toads.) In addition to their ecological importance, frogs have many cultural roles, such as in literature, symbolism and religion, and they are also valued as food and as pets.
Contents
Etymology and terminology
The name frog derives from Old English frogga, (compare Old Norse frauki, German Frosch, older Dutch spelling kikvorsch), cognate with Sanskrit plava (frog), probably deriving from Proto-Indo-European praw = "to jump".[1]
A distinction is often made between frogs and toads on the basis of their appearance, caused by the convergent adaptation among so-called toads to dry environments; however, this distinction has no taxonomic basis. The only family exclusively given the common name "toad" is Bufonidae, but many species from other families are also called "toads," and the species within the toad genus Atelopus are referred to as "harlequin frogs".
Taxonomy
For more details on this topic, see List of Anuran families.The order Anura contains 4,810 species[2] in 33 families, of which the Leptodactylidae (1100 spp.), Hylidae (800 spp.) and Ranidae (750 spp.) are the richest in species. About 88% of amphibian species are frogs.
 European Fire-bellied Toad (Bombina bombina)
European Fire-bellied Toad (Bombina bombina)
 Young American bullfrog found in a stream in New Jersey
Young American bullfrog found in a stream in New JerseyThe use of the common names "frog" and "toad" has no taxonomic justification. From a taxonomic perspective, all members of the order Anura are frogs, but only members of the family Bufonidae are considered "true toads". The use of the term "frog" in common names usually refers to species that are aquatic or semi-aquatic with smooth and/or moist skins, and the term "toad" generally refers to species that tend to be terrestrial with dry, warty skin. An exception is the fire-bellied toad (Bombina bombina): while its skin is slightly warty, it prefers a watery habitat.
Frogs and toads are broadly classified into three suborders: Archaeobatrachia, which includes four families of primitive frogs; Mesobatrachia, which includes five families of more evolutionary intermediate frogs; and Neobatrachia, by far the largest group, which contains the remaining 24 families of "modern" frogs, including most common species throughout the world. Neobatrachia is further divided into the Hyloidea and Ranoidea.[3] This classification is based on such morphological features as the number of vertebrae, the structure of the pectoral girdle, and the morphology of tadpoles. While this classification is largely accepted, relationships among families of frogs are still debated. Future studies of molecular genetics should soon provide further insights to the evolutionary relationships among anuran families.[4]
Some species of anurans hybridise readily. For instance, the Edible Frog (Rana esculenta) is a hybrid of the Pool Frog (R. lessonae) and the Marsh Frog (R. ridibunda). Bombina bombina and Bombina variegata similarly form hybrids, although these are less fertile, giving rise to a hybrid zone.
Morphology and physiology
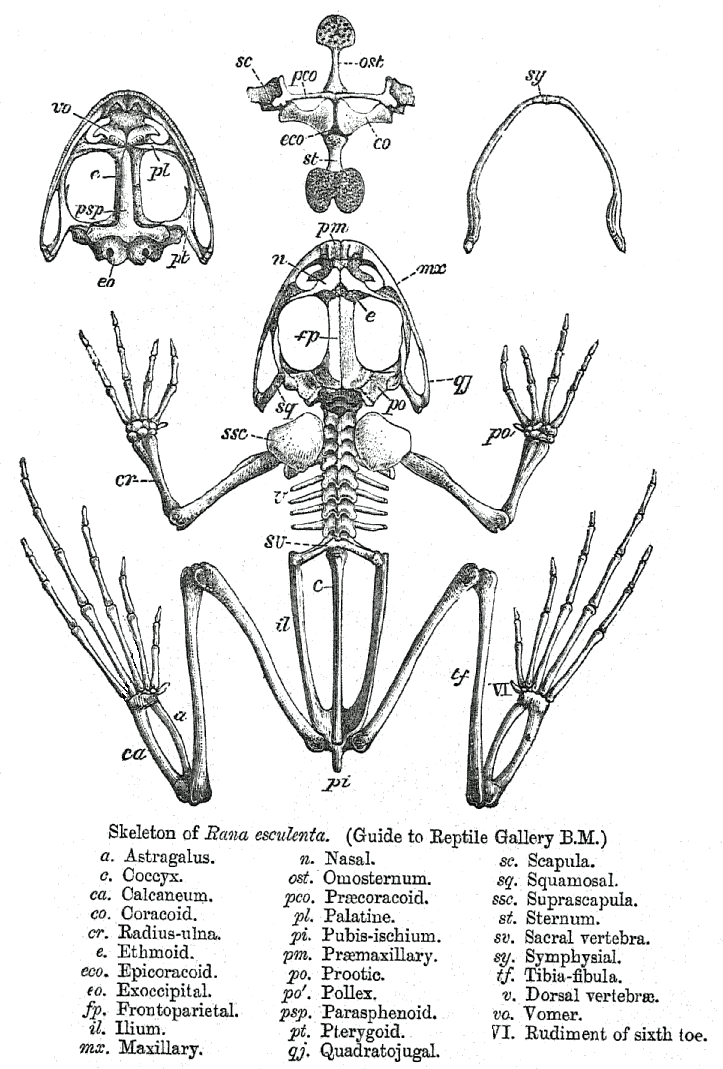
 Skeleton of Rana
Skeleton of RanaThe morphology of frogs is unique among amphibians. Compared with the other two groups of amphibians, (salamanders and caecilians), frogs are unusual because they lack tails as adults and their legs are more suited to jumping than walking. The physiology of frogs is generally like that of other amphibians (and differs from other terrestrial vertebrates) because oxygen can pass through their highly permeable skin. This unique feature allows frogs to "breathe" largely through their skin.[citation needed] Because the oxygen is dissolved in an aqueous film on the skin and passes from there to the blood, the skin must remain moist at all times; this makes frogs susceptible to many toxins in the environment, some of which can similarly dissolve in the layer of water and be passed into their bloodstream. This may be the cause of the decline in frog populations.[5][6][7][8][9][10][11][12][13]
Many characteristics are not shared by all of the approximately 5,250 described frog species. However, some general characteristics distinguish them from other amphibians. Frogs are usually well suited to jumping, with long hind legs and elongated ankle bones. They have a short vertebral column, with no more than ten free vertebrae, followed by a fused tailbone (urostyle or coccyx), typically resulting in a tailless phenotype.[citation needed]
Frogs range in size from 10 mm (0.39 in) (Brachycephalus didactylus of Brazil and Eleutherodactylus iberia of Cuba) to 300 mm (12 in) (goliath frog, Conraua goliath, of Cameroon). The skin hangs loosely on the body because of the lack of loose connective tissue. Skin texture varies: it can be smooth, warty or folded. Frogs have three eyelid membranes: one is transparent to protect the eyes underwater, and two vary from translucent to opaque. Frogs have a tympanum on each side of the head, which is involved in hearing and, in some species, is covered by skin. Most frogs have teeth, specifically pedicellate teeth in which the crown is separated from the root by fibrous tissue. Most only have teeth on the edge of the upper jaw (maxillary teeth) as well as vomerine teeth on the roof of their mouth. They do not have any teeth on their lower jaw, so they usually swallow their food whole. The teeth are mainly used to hold the prey and keep it in place till they can get a good grip on it and swallow their meal, assisted by retracting their eyes into their head.[14] True toads lack any teeth at all, and some species (Pyxicephalus) which prey on relatively large organisms (including mice and other frogs) have cone shaped projections of bone, called odontoid processes, at the front of the lower jaw which function like teeth.[2]
Feet and legs
 Tyler's Tree Frog (Litoria tyleri) illustrates large toe pads and webbed feet
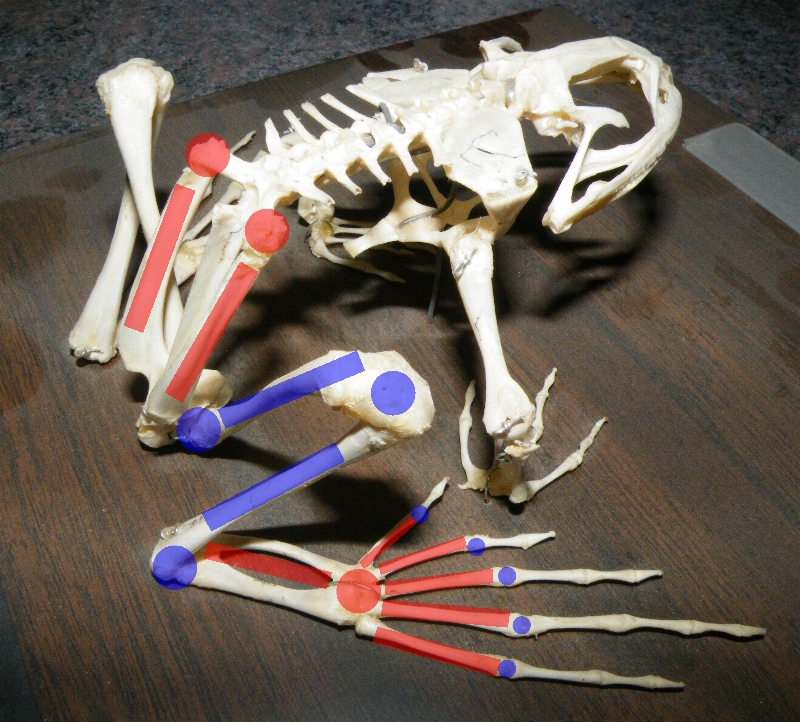
Tyler's Tree Frog (Litoria tyleri) illustrates large toe pads and webbed feet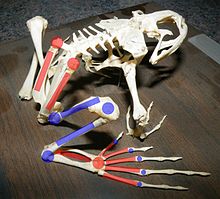 A bullfrog skeleton, showing elongate limb bones and extra joints. Red marks indicate bones which have been substantially elongated in frogs and joints which have become mobile. Blue indicates joints and bones which have not been modified or only somewhat elongated.
A bullfrog skeleton, showing elongate limb bones and extra joints. Red marks indicate bones which have been substantially elongated in frogs and joints which have become mobile. Blue indicates joints and bones which have not been modified or only somewhat elongated.The structure of the feet and legs varies greatly among frog species, depending in part on whether they live primarily on the ground, in water, in trees, or in burrows. Frogs must be able to move quickly through their environment to catch prey and escape predators, and numerous adaptations help them do so.
Many frogs, especially those that live in water, have webbed toes. The degree to which the toes are webbed is directly proportional to the amount of time the species lives in the water. For example, the completely aquatic African dwarf frog (Hymenochirus sp.) has fully webbed toes, whereas the toes of White's tree frog (Litoria caerulea), an arboreal species, are only a half or a quarter webbed.
 A frog gripping on a palm frond.
A frog gripping on a palm frond.Arboreal frogs have "toe pads" to help grip vertical surfaces. These pads, located on the ends of the toes, do not work by suction. Rather, the surface of the pad consists of interlocking cells, with a small gap between adjacent cells. When the frog applies pressure to the toe pads, the interlocking cells grip irregularities on the substrate. The small gaps between the cells drain away all but a thin layer of moisture on the pad, and maintain a grip through capillarity. This allows the frog to grip smooth surfaces, and does not function when the pads are excessively wet.[15]
In many arboreal frogs, a small "intercalary structure" in each toe increases the surface area touching the substrate. Furthermore, since hopping through trees can be dangerous, many arboreal frogs have hip joints that allow both hopping and walking. Some frogs that live high in trees even possess an elaborate degree of webbing between their toes, as do aquatic frogs. In these arboreal frogs, the webs allow the frogs to "parachute" or control their glide from one position in the canopy to another.[16]
Ground-dwelling frogs generally lack the adaptations of aquatic and arboreal frogs. Most have smaller toe pads, if any, and little webbing. Some burrowing frogs have a toe extension—a metatarsal tubercle—that helps them to burrow. The hind legs of ground dwellers are more muscular than those of aqueous and tree-dwelling frogs.
Sometimes during the tadpole stage, one of the animal's rear leg stubs is eaten by a dragonfly nymph. In some of these cases, the full leg grows anyway, and in other cases, it does not, although the frog may still live out its normal lifespan with only three legs. Other times, a parasitic flatworm called Riberoria trematodes digs into the rear of a tadpole, where it rearranges the limb bud cells, which sometimes causes the frog to have extra legs.[17]
Jumping
 Rainforest Rocket Frog jumping
Rainforest Rocket Frog jumpingFrogs are generally recognized as exceptional jumpers, and the best jumper of all vertebrates. The Australian rocket frog, Litoria nasuta, can leap over 50 times its body length (5.5 cm), resulting in jumps of over 2 meters. The acceleration of the jump may be up to twice gravity. There are tremendous differences between species in jumping capability, but within a species, jump distance increases with increasing size, but relative jumping distance (body-lengths jumped) decreases.
While frog species can use a variety of locomotor modes (running, walking, gliding, swimming, and climbing), more are either proficient at jumping or descended from ancestors who were, with much of the musculo-skeletal morphology modified for this purpose. The tibia, fibula and tarsals have been fused into a single, strong bone, as have the radius and ulna in the forelimbs (which must absorb the impact of landing). The metatarsals have become elongated to add to the leg length and allow the frog to push against the ground for longer during a jump. The illium has elongated and formed a mobile joint with the sacrum which, in specialist jumpers such as Ranids or Hylids, functions as an additional limb joint to further power the leaps. This elongation of the limbs results in the frog being able to apply force to the ground for longer during a jump, which in turn results in a longer, faster jump.[citation needed]
The muscular system has been similarly modified. The hind limbs of the ancestor of frogs presumably contained pairs of muscles which would act in opposition (one muscle to flex the knee, a different muscle to extend it), as is seen in most other limbed animals. However, in modern frogs, almost all muscles have been modified to contribute to the action of jumping, with only a few small muscles remaining to bring the limb back to the starting position and maintain posture. The muscles have also been greatly enlarged, with the muscles involved in jumping accounting for over 17% of the total mass of the frog.
In some extremely capable jumpers, such as the cuban tree frog, the peak power exerted during a jump can exceed what muscle is capable of producing. Currently, it is hypothesized that frogs are storing muscular energy by stretching their tendons like springs, then triggering the release all at once, allowing the frog to increase the energy of its jump beyond the limits of muscle-powered acceleration. A similar mechanism has already been documented in locusts and grasshoppers.[18]
Skin
 Pouched Frog (Assa darlingtoni) camouflaged against leaf litter
Pouched Frog (Assa darlingtoni) camouflaged against leaf litter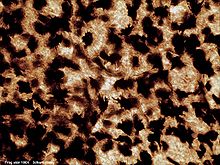 Microscopic view of frog skin
Microscopic view of frog skinMany frogs are able to absorb water and oxygen directly through the skin, especially around the pelvic area. However, the permeability of a frog's skin can also result in water loss. Some tree frogs reduce water loss with a waterproof layer of skin. Others have adapted behaviours to conserve water, including engaging in nocturnal activity and resting in a water-conserving position. This position involves the frog lying with its toes and fingers tucked under its body and chin, respectively, with no gap between the body and substrate. Some frog species will also rest in large groups, touching the skin of the neighbouring frog. This reduces the amount of skin exposed to the air or a dry surface, and thus reduces water loss. These adaptations only reduce water loss enough for a predominantly arboreal existence, and are not suitable for arid conditions.
Camouflage is a common defensive mechanism in frogs. Most camouflaged frogs are nocturnal, which adds to their ability to hide. Nocturnal frogs usually find the ideal camouflaged position during the day to sleep. Some frogs have the ability to change colour, but this is usually restricted to shades of one or two colours. For example, White's tree frog varies in shades of green and brown. Features such as warts and skin folds are usually found on ground-dwelling frogs, where a smooth skin would not disguise them effectively. Arboreal frogs usually have smooth skin, enabling them to disguise themselves as leaves.[citation needed]
Certain frogs change colour between night and day, as light and moisture stimulate the pigment cells and cause them to expand or contract.
Poison
Many frogs contain mild toxins that make them unpalatable to potential predators. For example, all toads have large poison glands—the parotoid glands—located behind the eyes, on the top of the head. Some frogs, such as some poison dart frogs, are especially toxic. The chemical makeup of toxins in frogs varies from irritants to hallucinogens, convulsants, nerve poisons, and vasoconstrictors. Many predators of frogs have adapted to tolerate high levels of these poisons. Others, including humans, may be severely affected.
 Oophaga pumilio, a poison dart frog, contains numerous alkaloids which deter predators
Oophaga pumilio, a poison dart frog, contains numerous alkaloids which deter predatorsSome frogs obtain poisons from the ants and other arthropods they eat;[19] others, such as the Australian Corroboree Frogs (Pseudophryne corroboree and Pseudophryne pengilleyi), can manufacture an alkaloid not derived from their diet.[20] Some native people of South America extract poison from the poison dart frogs and apply it to their darts for hunting,[21] although few species are toxic enough to be used for this purpose. It was previously a misconception the poison was placed on arrows rather than darts. The common name of these frogs was thus changed from "poison arrow frog" to "poison dart frog" in the early 1980s. Poisonous frogs tend to advertise their toxicity with bright colours, an adaptive strategy known as aposematism. There are at least two non-poisonous species of frogs in tropical America (Eleutherodactylus gaigei and Lithodytes lineatus) that mimic the colouration of dart poison frogs' coloration for self-protection (Batesian mimicry).[22][23]
Because frog toxins are extraordinarily diverse, they have raised the interest of biochemists as a "natural pharmacy". The alkaloid epibatidine, a painkiller 200 times more potent than morphine, is found in some species of poison dart frogs. Other chemicals isolated from the skin of frogs may offer resistance to HIV infection.[24] Arrow and dart poisons are under active investigation for their potential as therapeutic drugs.[25]
The skin secretions of some toads, such as the Colorado River toad and cane toad, contain bufotoxins, some of which, such as bufotenin, are psychoactive, and have therefore been used as recreational drugs. Typically, the skin secretions are dried and smoked. Skin licking is especially dangerous, and appears to constitute an urban myth. See psychoactive toad.
Respiration and circulation
The skin of a frog is permeable to oxygen and carbon dioxide, as well as to water. There are a number of blood vessels near the surface of the skin. When a frog is underwater, oxygen is transmitted through the skin directly into the bloodstream. On land, adult frogs use their lungs to breathe. Their lungs are similar to those of humans, but the chest muscles are not involved in respiration, and there are no ribs or diaphragm to support breathing. Frogs breathe by taking air in through the nostrils (which often have valves which close when the frog is submerged), causing the throat to puff out, then compressing the floor of the mouth, which forces the air into the lungs. In August 2007 an aquatic frog named Barbourula kalimantanensis was discovered in a remote part of Indonesia. The Bornean Flat-headed Frog (B. kalimantanensis) is the first species of frog known to science without lungs.
Frogs are known for their three-chambered heart, which they share with all tetrapods except birds, crocodilians and mammals. In the three-chambered heart, oxygenated blood from the lungs and de-oxygenated blood from the respiring tissues enter by separate atria, and are directed via a spiral valve to the appropriate vessel—aorta for oxygenated blood and pulmonary artery for deoxygenated blood. This special structure is essential to keeping the mixing of the two types of blood to a minimum, which enables frogs to have higher metabolic rates, and to be more active than otherwise.
Some species of frog have remarkable adaptations that allow them to survive in oxygen deficient water. The lake titicaca frog (Telmatobius culeus) is one such species and to survive in the poorly oxygenated waters of Lake Titicaca it has incredibly wrinkly skin that increases its surface area to enhance gas exchange. This frog will also do 'push-ups' on the lake bed to increase the flow of water around its body.[26]
Digestion and excretion
The frog's digestive system begins with the mouth. Frogs have teeth along their upper jaw called the maxillary teeth, which are used to grind food before swallowing. These teeth are very weak, and cannot be used to catch or harm agile prey. Instead, the frog uses its sticky tongue to catch food (such as flies or other insects). The food then moves through the esophagus into the stomach. The food then proceeds to the small intestine (duodenum and ileum) where most digestion occurs. Frogs carry pancreatic juice from the pancreas, and bile (produced by the liver) through the gallbladder from the liver to the small intestine, where the fluids digest the food and extract the nutrients. When the food passes into the large intestine, the water is reabsorbed and wastes are routed to the cloaca. All wastes exit the body through the cloaca and the cloacal vent.
Nervous system
The frog has a highly developed nervous system which consists of a brain, spinal cord and nerves. Many parts of the frog's brain correspond with those of humans. The medulla oblongata regulates respiration, digestion, and other automatic functions. Muscular coordination and posture are controlled by the cerebellum. The relative size of the cerebrum of a frog is much smaller than that of a human. Frogs have ten cranial nerves (nerves which pass information from the outside directly to the brain) and ten pairs of spinal nerves (nerves which pass information from extremities to the brain through the spinal cord). By contrast, all amniotes (mammals, birds and reptiles) have twelve cranial nerves. Frogs do not have external ears; the eardrums (tympanic membranes) are directly exposed. As in all animals, the ear contains semicircular canals which help control balance and orientation. Due to their short cochlea, frogs use electrical tuning to expand their range of audible frequencies.
Natural history
The life cycle of frogs, like that of other amphibians, consists of four main stages: egg, tadpole, metamorphosis and adult. The reliance of frogs on an aquatic environment for the egg and tadpole stages gives rise to a variety of breeding behaviours that include the well-known mating calls used by the males of most species to attract females to the bodies of water that they have chosen for breeding. Some frogs also look after their eggs—and in some cases even the tadpoles—for some time after laying.
Life cycle
 Frogspawn
Frogspawn Frogspawn development
Frogspawn development Tadpole of Haswell's Froglet (Paracrinia haswelli
Tadpole of Haswell's Froglet (Paracrinia haswelliThe life cycle of a frog starts with an egg. A female generally lays gelatinous egg masses containing thousands of eggs, in water. Each anuran species lays eggs in a distinctive, identifiable manner. An example are the long strings of eggs laid by the common American toad. The eggs are highly vulnerable to predation, so frogs have evolved many techniques to ensure the survival of the next generation. In colder areas the embryo is black to absorb more heat from the sun, which speeds up the development. Most commonly, this involves synchronous reproduction. Many individuals will breed at the same time, overwhelming the actions of predators; the majority of the offspring will still die due to predation, but there is a greater chance some will survive. Another way in which some species avoid the predators and pathogens eggs are exposed to in ponds is to lay eggs on leaves above the pond, with a gelatinous coating designed to retain moisture. In these species the tadpoles drop into the water upon hatching. The eggs of some species laid out of water can detect vibrations of nearby predatory wasps or snakes, and will hatch early to avoid being eaten.[27] Some species, such as the Cane Toad (Bufo marinus), lay poisonous eggs to minimise predation. While the length of the egg stage depends on the species and environmental conditions, aquatic eggs generally hatch within one week. Other species go through their whole larval phase inside the eggs or the mother, or they have direct development. Unlike salamanders and newts, frogs and toads never become sexually mature while still in their larval stage.
Eggs hatch and continue life as tadpoles (occasionally known as polliwogs), which typically have oval bodies and long, vertically flattened tails. At least one species (Nannophrys ceylonensis) has tadpoles that are semi-terrestrial and live among wet rocks,[28][29] but as a general rule, free living larvae are fully aquatic. They lack eyelids and have a cartilaginous skeleton, a lateral line system, gills for respiration (external gills at first, internal gills later) and tails with dorsal and ventral folds of skin for swimming.[30] From pretty early onward they develop a gill pouch that covers the gills and the front legs and also the lungs are developed in an early stage as an accessory breathing organ. Some species which go through the metamorphosis inside the egg and hatch to small frogs never develop gills, instead there are specialised areas of skin that takes care of the respiration. Tadpoles also lack true teeth, but the jaws in most species usually have two elongate, parallel rows of small keratinized structures called keradonts in the upper jaw while the lower jaw has three rows of keradonts, surrounded by a horny beak, but the number of rows can be lower or absent, or much higher.[31] Tadpoles are typically herbivorous, feeding mostly on algae, including diatoms filtered from the water through the gills. Some species are carnivorous at the tadpole stage, eating insects, smaller tadpoles, and fish. Cannibalism has been observed among tadpoles. Early developers who gain legs may be eaten by the others, so the late bloomers survive longer. This has been observed in England in the species Rana temporaria (common frog).[32]
Tadpoles are highly vulnerable to predation by fish, newts, predatory diving beetles and birds such as kingfishers. Poisonous tadpoles are present in many species, such as Cane Toads. The tadpole stage may be as short as a week, or tadpoles may overwinter and metamorphose the following year in some species, such as the midwife toad (Alytes obstetricans) and the common spadefoot (Pelobates fuscus). In the Pipidae, with the exception for Hymenochirus, the tadpoles have paired anterior barbels which make them resemble small catfish.[33]
With the exception of the base of the tail, where a few vertebral structures develop to give rise to the urostyle later in life, the tail lacks the completely solid, segmental, skeletal elements of cartilage or bony tissue that are so typical for other vertebrates, although it does contain a notochord
At the end of the tadpole stage, frogs undergo metamorphosis, in which they undergo a transition into the adult form. This metamorphosis last typically only 24 hours and consists of:
 Larva of the common frog Rana temporaria a day before metamorphisis
Larva of the common frog Rana temporaria a day before metamorphisis Common frog - Metamorphosis stage. Notice the deformed jaws, large eyes and the remains of the gill pouch.
Common frog - Metamorphosis stage. Notice the deformed jaws, large eyes and the remains of the gill pouch. Young frog with tail remains after metamorphosis
Young frog with tail remains after metamorphosis Adult leopard frog
Adult leopard frog- The disappearance of the gill pouch, making the front legs visible.
- The transformation of the jaws into the big jaws of predatory frogs (most tadpoles are scrapers of algae or are filter feeders)
- The transformation of the digestive system: the long spiral gut of the larva is being replaced by the typical short gut of a predator.
- An adaptation of the nervous system for stereoscopic vision, locomotion and feeding
- A quick growth and movement of the eyes to higher up the skull and the formation of eyelids.
- Formation of skin glands, thickening of the skin and loss of the lateral line system
- An eardrum is developed to lock the middle ear.
The disappearance of the tail is somewhat later (occurs at higher thyroxin levels) and after the tail has been resorbed the animals are ready to leave the water. The material of the tail is being used for a quick growth of the legs. The disappearing of the larval structures is a regulated process called apoptosis.
 Incident of frog cannibalism
Incident of frog cannibalismAfter metamorphosis, young adults may leave the water and disperse into terrestrial habitats, or continue to live in the aquatic habitat as adults. Almost all species of frogs are carnivorous as adults, eating invertebrates such as arthropods, annelids and gastropods. A few of the larger species may eat prey such as small mammals, fish and smaller frogs. Some frogs use their sticky tongues to catch fast-moving prey, while others capture their prey and force it into their mouths with their hands. However, there are a very few species of frogs that primarily eat plants.[34] Adult frogs are themselves preyed upon by birds, large fish, snakes, otters, foxes, badgers, coatis, and other animals. Frogs are also eaten by people (see section on uses in agriculture and research, below).
Frogs and toads can live for many years; though little is known about their life span in the wild, captive frogs and toads are recorded living up to 40 years.[35]
Frogs from temperate climates hibernate through the winter, and 4 species are known to freeze during this time, most notably Rana sylvatica.[36]
Reproduction of frogs
Once adult frogs reach maturity, they will assemble at a water source such as a pond or stream to breed. Many frogs return to the bodies of water where they were born, often resulting in annual migrations involving thousands of frogs. In continental Europe, a large proportion of migrating frogs used to die on roads, before special fences and tunnels were built for them.

 A Male and Female common toad in amplexus. The black strands are eggs released into open water minutes after birth.
A Male and Female common toad in amplexus. The black strands are eggs released into open water minutes after birth.Once at the breeding ground, male frogs call to attract a mate, collectively becoming a chorus of frogs. The call is unique to the species, and will attract females of that species. Some species have satellite males who do not call, but intercept females that are approaching a calling male.
The male and female frogs then undergo amplexus. This involves the male mounting the female and gripping her (sometimes with special nuptial pads) tightly. Fertilization is external: the egg and sperm meet outside of the body. The female releases her eggs, which the male frog covers with a sperm solution. The eggs then swell and develop a protective coating. The eggs are typically brown or black, with a clear, gelatin-like covering.
Most temperate species of frogs reproduce between late autumn and early spring. In the UK, most common frog populations produce frogspawn in February, although there is wide variation in timing. Water temperatures at this time of year are relatively low, typically between four and 10 degrees Celsius. Reproducing in these conditions helps the developing tadpoles because dissolved oxygen concentrations in the water are highest at cold temperatures. More importantly, reproducing early in the season ensures that appropriate food is available to the developing frogs at the right time.
Parental care
 Colour plate from Ernst Haeckel's 1904 Kunstformen der Natur, depicting frog species that include two examples of parental care.
Colour plate from Ernst Haeckel's 1904 Kunstformen der Natur, depicting frog species that include two examples of parental care.Although care of offspring is poorly understood in frogs, it is estimated that up to 20% of amphibian species may care for their young in one way or another, and there is a great diversity of parental behaviours.[37] Some species of poison dart frog lay eggs on the forest floor and protect them, guarding the eggs from predation and keeping them moist. The frog will urinate on them if they become too dry. After hatching, a parent (the sex depends upon the species) will move them, on its back, to a water-holding bromeliad. The parent then feeds them by laying unfertilized eggs in the bromeliad until the young have metamorphosed. Other frogs carry the eggs and tadpoles on their hind legs or back (e.g. the midwife toads, Alytes spp.). Some frogs even protect their offspring inside their own bodies. The male Australian Pouched Frog (Assa darlingtoni) has pouches along its side in which the tadpoles reside until metamorphosis. The female Gastric-brooding Frogs (genus Rheobatrachus) from Australia, now probably extinct, swallows its tadpoles, which then develop in the stomach. To do this, the Gastric-brooding Frog must stop secreting stomach acid and suppress peristalsis (contractions of the stomach). Darwin's Frog (Rhinoderma darwinii) from Chile puts the tadpoles in its vocal sac (see next section) for development. Some species of frog will leave a 'babysitter' to watch over the frogspawn until it hatches.
The evolution of parental care in frogs is driven primarily by the size of the water body in which they breed. There is an inverse relationship between the level of parental care in a frog species and the size of the body of water—frogs that breed in smaller water bodies tend to have more complex parental care behaviors.[38] Water body size shows this strong relationship with parental care because it encompasses several important variables that interact to select for parental care: predation, desiccation, competition, and resource limitation. Because predation of eggs and larvae is high in large water bodies, a number of frog species evolved terrestrial oviposition. Once eggs are deposited on land, the desiccating terrestrial environment demands uniparental care in the form of egg hydration to ensure egg survival.[39] The subsequent need to transport hatched tadpoles to a water source requires an even more intense form of uniparental care. In small water bodies where predators are mostly absent, such as phytotelmata (water-filled leaf axils or small woody cavities), inter-tadpole competition becomes the variable that constrains tadpole survival. Certain frogs species avoid this competition by evolving the use of smaller phytotelmata as tadpole deposition sites.[40] However, while these smaller tadpole rearing sites are free of competition, they also lack nutrients. Because they do not have sufficient nutrients to support a tadpole without parental provisioning behavior, frog species that transitioned from the use of larger to smaller phytotelmata have evolved trophic (unfertilized) egg laying. In this complex form of biparental care, the female provides her offspring with nutritive eggs. While each of these variables select for different behaviors, they correlate with the size of a species' tadpole-rearing site and influence the degree of parental care displayed by a species.
Call
 A male Dendropsophus microcephalus displaying its vocal sac during its call.
A male Dendropsophus microcephalus displaying its vocal sac during its call.Some frog calls are so loud, they can be heard up to a mile away.[41] The call of a frog is unique to its species. Frogs call by passing air through the larynx in the throat. In most calling frogs, the sound is amplified by one or more vocal sacs, membranes of skin under the throat or on the corner of the mouth that distend during the amplification of the call. The field of neuroethology studies the neurocircuitry that underlies frog audition.
Some frogs lack vocal sacs, such as those from the genera Heleioporus and Neobatrachus, but these species can still produce a loud call. Their buccal cavity is enlarged and dome-shaped, acting as a resonance chamber that amplifies their call. Species of frog without vocal sacs and that do not have a loud call tend to inhabit areas close to flowing water. The noise of flowing water overpowers any call, so they must communicate by other means.
The main reason for calling is to allow males to attract a mate. Males call either individually or in a group called a chorus. Females of many frog species, for example Polypedates leucomystax, produce calls reciprocal to the males', which act as the catalyst for the enhancement of reproductive activity in a breeding colony.[42] A male frog emits a release call when mounted by another male. Tropical species also have a rain call that they make on the basis of humidity cues prior to a rain shower. Many species also have a territorial call that is used to chase away other males. All of these calls are emitted with the mouth of the frog closed.
A distress call, emitted by some frogs when they are in danger, is produced with the mouth open, resulting in a higher-pitched call. The effectiveness of the call is unknown; however, it is suspected the call intrigues the predator until another animal is attracted, distracting them enough for its escape.
Many species of frog have deep calls, or croaks. The English onomatopoeic spelling is "ribbit". The croak of the American bullfrog (Rana catesbiana) is sometimes spelt "jug o' rum".[43] Other examples are Ancient Greek brekekekex koax koax for probably Rana ridibunda, and the description in Rigveda 7:103.6 gómāyur éko ajámāyur ékaħ = "one has a voice like a cow's, one has a voice like a goat's".
Distribution and conservation status
 See also: Decline in amphibian populations
See also: Decline in amphibian populationsThe habitat of frogs extends almost worldwide, but they do not occur in Antarctica and are not present on many oceanic islands.[44][45] The greatest diversity of frogs occurs in the tropical areas of the world, where water is readily available, suiting frogs' requirements due to their skin. Some frogs inhabit arid areas such as deserts, where water may not be easily accessible, and rely on specific adaptations to survive. The Australian genus Cyclorana and the American genus Pternohyla will bury themselves underground, create a water-impervious cocoon and hibernate during dry periods. Once it rains, they emerge, find a temporary pond and breed. Egg and tadpole development is very fast in comparison to most other frogs so that breeding is complete before the pond dries up. Some frog species are adapted to a cold environment; for instance the wood frog, whose habitat extends north of the Arctic Circle, buries itself in the ground during winter when much of its body freezes.
 Golden toad (Bufo periglenes) – last seen in 1989
Golden toad (Bufo periglenes) – last seen in 1989Frog populations have declined dramatically since the 1950s: more than one third of species are believed to be threatened with extinction and more than 120 species are suspected to be extinct since the 1980s.[46] Among these species are the golden toad of Costa Rica and the Gastric-brooding frogs of Australia. Habitat loss is a significant cause of frog population decline, as are pollutants, climate change, the introduction of non-indigenous predators/competitors, and emerging infectious diseases including chytridiomycosis. Many environmental scientists believe that amphibians, including frogs, are excellent biological indicators of broader ecosystem health because of their intermediate position in food webs, permeable skins, and typically biphasic life (aquatic larvae and terrestrial adults).[47] It appears that it is the species with both aquatic eggs and aquatic larvae that are most affected by the decline, while those with direct development are the most resistant.[48]
A Canadian study conducted in 2006, suggested heavy traffic near frog habitats as a large threat to frog populations.[49] In a few cases, captive breeding programs have been attempted to alleviate the pressure on frog populations, and these have proved successful.[50][51] In 2007, it was reported the application of certain probiotic bacteria could protect amphibians from chytridiomycosis.[52] One current project, The Panama Amphibian Rescue and Conservation Project, has subsequently been developed in order to rescue species at risk of chytridiomycosis in eastern Panama, and to develop field applications of this probiotic cure.[53]
Zoos and aquariums around the world named 2008 the Year of the Frog, to draw attention to the conservation issues.[54]
Evolution
 A fossilized frog from the Czech Republic, possibly Palaeobatrachus gigas
A fossilized frog from the Czech Republic, possibly Palaeobatrachus gigasUntil the discovery of the Early Permian Gerobatrachus hottoni in 2008, a stem-batrachian with many salamander-like characteristics, the earliest known proto-frog was Triadobatrachus massinoti, from the 250 million year old early Triassic of Madagascar.[55] The skull is frog-like, being broad with large eye sockets, but the fossil has features diverging from modern amphibia. These include a different ilium, a longer body with more vertebrae, and separate vertebrae in its tail (whereas in modern frogs, the tail vertebrae are fused, and known as the urostyle or coccyx). The tibia and fibula bones are unfused and separate, making it probable Triadobatrachus was not an efficient leaper.
Another fossil frog, Prosalirus bitis, was discovered in 1995. The remains were recovered from Arizona's Kayenta Formation, which dates back to the Early Jurassic epoch,[56] somewhat younger than Triadobatrachus. Like Triadobatrachus, Prosalirus did not have greatly enlarged legs, but had the typical three-pronged pelvic structure. Unlike Triadobatrachus, Prosalirus had already lost nearly all of its tail[citation needed] and was well adapted for jumping.[57]
The earliest known "true frog" is Vieraella herbsti, from the early Jurassic (188–213 million years ago). It is known only from the dorsal and ventral impressions of a single animal and was estimated to be 33 mm (1.3 in) from snout to vent. Notobatrachus degiustoi from the middle Jurassic is slightly younger, about 155–170 million years old. It is likely the evolution of modern Anura was completed by the Jurassic period. The main evolutionary changes involved the shortening of the body and the loss of the tail.
The earliest full fossil record of a modern frog is of sanyanlichan, which lived 125 million years ago[58] and had all modern frog features, but bore 9 presacral vertebrae instead of the 8 of modern frogs.[59]
Frog fossils have been found on all continents except Antarctica, but biogeographic evidence suggests they inhabited Antarctica when it was warmer.[60]
Uses in agriculture and research
For more details on this topic, see Animal testing on frogs.Frogs are raised commercially for several purposes. Frogs are used as a food source; frog legs are a delicacy in China, France, Indonesia, the Philippines, the north of Greece and in many parts of the American South, especially Louisiana. Dead frogs are sometimes used for dissections in high school and university anatomy classes, often after being injected with coloured plastics to enhance the contrast between the organs. This practice has declined in recent years with the increasing concerns about animal welfare.
Frogs have served as important model organisms throughout the history of science. Eighteenth-century biologist Luigi Galvani discovered the link between electricity and the nervous system through studying frogs. The African clawed frog or platanna (Xenopus laevis) was first widely used in laboratories in pregnancy assays in the first half of the 20th century. When human chorionic gonadotropin, a hormone found in substantial quantities in the urine of pregnant women, is injected into a female X. laevis, it induces them to lay eggs. In 1952, Robert Briggs and Thomas J. King cloned a frog by somatic cell nuclear transfer, the same technique later used to create Dolly the Sheep, their experiment was the first time successful nuclear transplantation had been accomplished in metazoans.[61]
Frogs are used in cloning research and other branches of embryology because frogs are among the closest living relatives of man to lack egg shells characteristic of most other vertebrates, and therefore facilitate observations of early development. Although alternative pregnancy assays have been developed, biologists continue to use Xenopus as a model organism in developmental biology because it is easy to raise in captivity and has a large and easily manipulatable embryo. Recently, X. laevis is increasingly being displaced by its smaller relative X. tropicalis, which reaches its reproductive age in five months rather than one to two years (as in X. laevis),[62] facilitating faster studies across generations. The genome sequence of X. tropicalis will probably be completed by 2015 at the latest.[63]
Cultural beliefs
For more details on this topic, see Frogs in popular culture. Moche Frog 200 A.D. Larco Museum Collection Lima, Peru
Moche Frog 200 A.D. Larco Museum Collection Lima, PeruFrogs feature prominently in folklore, fairy tales and popular culture. They tend to be portrayed as benign, ugly, clumsy, but with hidden talents. Examples include Michigan J. Frog, The Frog Prince, and Kermit the Frog. Michigan J. Frog, featured in the Warner Brothers cartoon One Froggy Evening, only performs his singing and dancing routine for his owner. Once another person looks at him, he will return to a frog-like pose. "The Frog Prince" is a fairy tale of a frog who turns into a handsome prince once kissed. Kermit the Frog, on the other hand, is a conscientious and disciplined character of The Muppet Show and Sesame Street; while openly friendly and greatly talented, he is often portrayed as cringing at the fanciful behavior of more flamboyant characters.
The Moche people of ancient Peru worshipped animals and often depicted frogs in their art.[64] In Panama local legend promised luck to anyone who spotted a golden frog in the wild and some believed that when Panamanian Golden Frogs died, they would turn into a gold talisman, known as a huaca. Today, despite being extinct in the wild, Panamanian Golden Frogs remain an important cultural symbol and can be found on decorative cloth molas made by the Kuna Indians, on T-shirts, as inlaid design on a new overpass in Panama City and even on lottery tickets.[65]
See also
References
Notes
- ^ "Indo-European etymology database". http://ehl.santafe.edu/cgi-bin/response.cgi?root=config&morpho=0&basename=/data/ie/piet&first=2221.
- ^ a b Pough et al. 1992. Herpetology: Third Edition. Pearson Prentice Hall:Pearson Education, Inc., 2002.
- ^ Ford, L.S.; D.C. Cannatella (1993). "The major clades of frogs". Herpetological Monographs 7: 94–117. doi:10.2307/1466954. JSTOR 1466954.
- ^ Faivovich, J.; C.F.B. Haddad, P.C.A. Garcia, D.R. Frost, J.A. Campbell, and W.C. Wheeler (2005). "Systematic review of the frog family Hylidae, with special reference to Hylinae: Phylogenetic analysis and revision". Bulletin of the American Museum of Natural History 294: 1–240. doi:10.1206/0003-0090(2005)294[0001:SROTFF]2.0.CO;2.
- ^ Blaustein, Andrew R & Pieter TJ Johnson (2003). "The complexity of deformed amphibians" (PDF). Front. Ecol. Environ. 1 (2): 87–94. doi:10.1890/1540-9295(2003)001[0087:TCODA]2.0.CO;2. http://tiee.ecoed.net/vol/v2/issues/frontier_sets/amphibians/pdf/Frontiers-Blaustein-Johnson.pdf.
- ^ Burkhart, James G.; Gerald Ankley, Heidi Bell, Hillary Carpenter, Douglas Fort, David Gardiner, Henry Gardner, Robert Hale, Judy C. Helgen, Paul Jepson, Douglas Johnson, Michael Lannoo, David Lee, Joseph Lary, Rick Levey, Joseph Magner, Carol Meteyer, Michael D. Shelby, and George Lucier (2000). "Strategies for Assessing the Implications of Malformed Frogs for Environmental Health". Environmental Health Perspectives 108 (1): 83. doi:10.2307/3454299. JSTOR 3454299. PMC 1637865. PMID 10620528. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1637865.
- ^ Relyea, R.A. (2004). "The impact of insecticides and herbicides on the biodiversity and productivity of aquatic communities". Ecological Applications 15 (2): 618–627. doi:10.1890/03-5342.
- ^ Relyea, R.A. (2005). "The lethal impact of Roundup on aquatic and terrestrial amphibians". Ecological Applications 15 (4): 1118–1124. doi:10.1890/04-1291.
- ^ Hayes, T. B.; Collins, A.; Lee, M.; Mendoza, M.; Noriega, N.; Stuart, A. A.; Vonk, A. (2002). "Hermaphroditic demasculinized frogs after exposure to the herbicide atrazine at low ecologically relevant doses". Proc. Natl. Acad. Sci. 99 (8): 5476–5480. doi:10.1073/pnas.082121499. PMC 122794. PMID 11960004. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=122794.
- ^ "Pollutants change 'he' frogs into 'she' frogs" (in Yahoo! News). 2007. Archived from the original on 2007-03-02. http://web.archive.org/web/20070302073022/http://news.yahoo.com/s/afp/20070227/sc_afp/scienceenvironmentanimalssexfrogs_070227120242. Retrieved 2007-03-01.
- ^ Johnson PTJ, Jonathan M. Chase, Katherine L. Dosch,Richard B. Hartson, Jackson A. Gross,Don J. Larson, Daniel R. Sutherland and Stephen R. Carpenter (2007). "Aquatic eutrophication promotes pathogenic infection in amphibians". PNAS 104 (40): 15781–15786. doi:10.1073/pnas.0707763104. PMC 2000446. PMID 17893332. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2000446.
- ^ Grossi, Mark (24 July 2001). "Sierra Frogs Fall Silent". The Fresno Bee. http://www.yosemite.org/newsroom/clips2001/July/072401.html. Retrieved 2008-10-02.[dead link]
- ^ Dohm, M.R.; Mautz, WJ; Andrade, JA; Gellert, KS; Salas-Ferguson, LJ; Nicolaisen, N; Fujie, N (2005). "Effects of ozone exposure on nonspecific phagocytic capacity of pulmonary macrophages from an amphibian, Bufo marinus". Environmental Toxicology and Chemistry 24 (1): 205–210. doi:10.1897/04-040R.1. PMID 15683185.
- ^ Levine, R. P.; Monroy, JA; Brainerd, EL (2004-03-15). "Contribution of eye retraction to swallowing performance in the northern leopard frog, Rana pipiens". Journal of Experimental Biology 207 (Pt 8): 1361–8. doi:10.1242/jeb.00885. PMID 15010487. http://jeb.biologists.org/cgi/reprint/207/8/1361. Retrieved 2010-03-18.
- ^ Emerson, S.B.; Diehl, D. (1980). "Toe pad morphology and mechanisms of sticking in frogs". Biol. J. Linn. Soc. 13 (3): 199–216. doi:10.1111/j.1095-8312.1980.tb00082.x.
- ^ Harvey, M. B; A. J. Pemberton, and E. N. Smith (2002). "New and poorly known parachuting frogs (Rhacophoridae : Rhacophorus) from Sumatra and Java". Herpetological Monographs 16: 46–92. doi:10.1655/0733-1347(2002)016[0046:NAPKPF]2.0.CO;2.
- ^ Walker, Matt (June 25, 2009). "Legless frogs mystery solved". BBC News. http://news.bbc.co.uk/earth/hi/earth_news/newsid_8116000/8116692.stm.
- ^ Scott, J. (2005). "The locust jump: an integrated laboratory investigation". AJP: Advances in Physiology Education 29 (1): 21. doi:10.1152/advan.00037.2004. http://advan.physiology.org/cgi/content/full/29/1/21#R13. Retrieved 2010-03-18.
- ^ Saporito, R.A.; H.M. Garraffo, M.A. Donnelly, A.L. Edwards, J.T. Longino, and J.W. Daly (2004). "Formicine ants: An arthropod source for the pumiliotoxin alkaloids of dendrobatid poison frogs". Proceedings of the National Academy of Science 101 (21): 8045–8050. doi:10.1073/pnas.0402365101. PMC 419554. PMID 15128938. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=419554.
- ^ Smith, B. P.; Tyler M. J., Kaneko T., Garraffo H. M., Spande T. F., Daly J. W. (2002). "Evidence for biosynthesis of pseudophrynamine alkaloids by an Australian myobatrachid frog (pseudophryne) and for sequestration of dietary pumiliotoxins". J Nat Prod 65 (4): 439–47. doi:10.1021/np010506a. PMID 11975476.
- ^ Myers, C.W.; J.W. Daly (1983). "Dart-poison frogs". Scientific American 248 (2): 120–133. doi:10.1038/scientificamerican0283-120. PMID 6836257.
- ^ Savage, J. M. (2002). The Amphibians and Reptiles of Costa Rica. University of Chicago Press, Chicago. ISBN 0916984168.
- ^ Duellman, W. E. (1978). "The Biology of an Equatorial Herpetofauna in Amazonian Ecuador" (PDF). University of Kansas Museum of Natural History Miscellaneous Publication 65: 1–352. http://200.31.31.2/Recursos/publicaciones/Cientifica/dulleman1978.pdf.
- ^ VanCompernolle, Scott. E.; R. J. Taylor, K. Oswald-Richter, J. Jiang, B. E. Youree, J. H. Bowie, M. J. Tyler, M. Conlon, D. Wade, C. Aiken, T. S. Dermody, V. N. KewalRamani, L. A. Rollins-Smith and D. Unutmaz (2005). "Antimicrobial peptides from amphibian skin potently inhibit Human Immunodeficiency Virus infection and transfer of virus from dendritic cells to T cells". Journal of Virology 79 (18): 11598–11606. doi:10.1128/JVI.79.18.11598-11606.2005. PMC 1212620. PMID 16140737. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1212620.
- ^ Phillipe, G.; Angenot L. (2005). "Recent developments in the field of arrow and dart poisons". J Ethnopharmacol 100(1–2) (1-2): 85–91. doi:10.1016/j.jep.2005.05.022. PMID 15993556.
- ^ Piper, Ross (2007). Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals. Greenwood Press. ISBN 0313339228.
- ^ Warkentin, K.M. (1995). "Adaptive plasticity in hatching age: a response to predation risk trade-offs". Proceedings of the National Academy of Sciences 92: 3507–3510. doi:10.1073/pnas.92.8.3507.
- ^ "Ontogenetic Changes in Diet and Intestinal Morphology in Semi-Terrestrial Tadpoles of Nannophrys ceylonensis (Dicroglossidae)". http://www.encyclopedia.com/doc/1P.html.
- ^ "Nannophrys ceylonensis - Sri Lanka rock frog". Amphibiaweb.org. 2005-05-10. http://amphibiaweb.org/cgi-bin/amphib_query?query_src=aw_lists_genera_&table=amphib&where-genus=Nannophrys&where-species=ceylonensis. Retrieved 2009-03-06.
- ^ "Anura :: From tadpole to adult - Britannica Online Encyclopedia". Britannica.com. http://www.britannica.com/eb/article-40603/Anura. Retrieved 2008-11-03.
- ^ "Larvae: Information from". Answers.com. http://www.answers.com/topic/larvae. Retrieved 2010-03-18.
- ^ Frogs Found in the U.K.. Retrieved 18 July 2007.[dead link]
- ^ "AmphibiaWeb - Pipidae". Amphibiaweb.org. http://amphibiaweb.org/lists/Pipidae.shtml. Retrieved 2008-11-03.
- ^ Silva, H. R.; Britto-Pereira M. C., & Caramaschi U. (1989). "Frugivory and Seed Dispersal by Hyla truncata, a Neotropical Treefrog". Copeia 1989 (3): 781–783. doi:10.2307/1445517. JSTOR 1445517.
- ^ Curry-Lindahl (1966). Biegler. see also http://www.pondturtle.com/lfrog.html#Bufo
- ^ Storey, KB (1990). "Life in a frozen state: adaptive strategies for natural freeze tolerance in amphibians and reptiles.". American Journal of Physiology 258 (3 Pt 2): 559–568. PMID 2180324.
- ^ Crump, M.L. (1996). "Parental care among the Amphibia". Advances in the Study of Behavior 25: 109–144. doi:10.1016/S0065-3454(08)60331-9.
- ^ Brown, J. L.; Morales, V.; Summers, K. (2010). "A key ecological trait drove the evolution of biparental care and monogamy in an amphibian". American Naturalist 175 (4): 436–446. doi:10.1086/650727. PMID 20180700.
- ^ Sheridan, Jennifer A.; Ocock, Joanne F. (2008). "Parental Care in Chiromantis hansenae (Anura: Rhacophoridae)". Copeia 2008: 733–736. doi:10.1643/CH-07-225. http://findarticles.com/p/articles/mi_7513/is_200812/ai_n32310466/.
- ^ Brown, J. L.; Morales, V.; Summers, K. (2008a). "Divergence in parental care, habitat selection and larval life history between two species of peruvian poison frogs: An experimental analysis". Journal of Evolutionary Biology 21 (6): 1534–1543. doi:10.1111/j.1420-9101.2008.01609.x. PMID 18811668.
- ^ See, for instance, Ohio's Toads and Frogs by the Ohio Department of Natural Resources. Retrieved 18 July 2007.
- ^ Roy, Debjani (1997). "Communication signals and sexual selection in amphibians" (PDF). Current Science 72: 923–927. http://www.ias.ac.in/jarch/currsci/72/00000944.pdf.
- ^ Hilton, Bill Jr. (1986-06-08). Jug-o-Rum: Call of the Amorous Bullfrog. 1. Hilton Pond Center for Piedmont Natural History. http://www.hiltonpond.org/PNBullfrog860608.html. Retrieved 2008-06-26.
- ^ "Freaky Frogs," at National Geographic Explorer. Retrieved 18 July 2007.
- ^ Evolution Encyclopedia, Volume 3: Geographical Distribution. Retrieved 18 July 2007.
- ^ Stuart, S.N.; J.S. Chanson, N.A. Cox, B.E. Young, A.S.L. Rodrigues, D.L. Fischman, and R.W. Waller (2004). "Status and trends of amphibian declines and extinctions worldwide". Science 306 (5702): 1783–1786. doi:10.1126/science.1103538. PMID 15486254.
- ^ Phillips, Kathryn (1994). Tracking the Vanishing Frogs. New York: Penguin Books. ISBN 0-14-024646-0.
- ^ Lips, Karen R. (2008). "Decline of a Tropical Montane Amphibian Fauna". Conservation Biology 12: 106–117. doi:10.1111/j.1523-1739.1998.96359.x. JSTOR 2387466. http://www.jcu.edu.au/school/phtm/PHTM/frogs/papers/lips-1998.pdf.
- ^ New Scientist (July 7, 2006). "Frog population decrease mostly due to traffic". New Scientist (advance online). http://www.newscientist.com/article/dn9506-frogs-toads-and-automobiles--a-fatal-combination.html.
- ^ Richard Black (2005-10-02). "New frog centre for London Zoo". BBC News. http://news.bbc.co.uk/1/hi/sci/tech/4298050.stm. Retrieved 2008-11-03.
- ^ "National recovery plan for the Southern Corroboree Frog (Pseudophryne corroboree): 5. Previous Recovery Actions". Environment.gov.au. http://www.environment.gov.au/biodiversity/threatened/publications/recovery/p-corroboree/part4.html. Retrieved 2008-11-03.
- ^ "Bacteria show promise in fending off global amphibian killer". Physorg.com. http://www.physorg.com/news99134333.html. Retrieved 2008-11-03.
- ^ "Project launched to fight frog-killing fungus". London: http://www.guardian.co.uk.+2008-01-23. http://www.guardian.co.uk/world/feedarticle/8502055. Retrieved 2009-07-19.
- ^ MacNeill, R., Saving Kermit, Canadian Geographic Magazine: April 2008, p. 19
- ^ Cannatella, David (1995). "Triadobatrachus massinoti". Tree of Life. http://www.tolweb.org/Triadobatrachus_massinoti/16962. Retrieved 2008-06-26.
- ^ Weishampel, David B; et al (2004). "Dinosaur distribution (Early Jurassic, North America)." In: Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. Pp. 530-532. ISBN 0-520-24209-2.
- ^ Foster, J. (2007). "Anura (Frogs)". Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 135–136. ISBN 0253348706.
- ^ "China Yields East Asia's Earliest Fossilized Frog". People's Daily. 2001-11-20. http://english.peopledaily.com.cn/200111/20/eng20011120_84925.shtml. Retrieved 2008-06-26.
- ^ "Chinese frog discovery sheds light on amphibians' evolution". The Dhamurian Society (Australian Broadcasting Company). 2001-11-20. http://www.dhamurian.org.au/zoology/chinesefrog.html. Retrieved 2008-06-26.
- ^ Evans, S. E.; Jones, M. E. H.; Krause, D. W. (2008). "A giant frog with South American affinities from the Late Cretaceous of Madagascar". Proceedings of the National Academy of Sciences 105 (8): 2951–6. doi:10.1073/pnas.0707599105. PMC 2268566. PMID 18287076. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2268566.
- ^ "Robert W. Briggs Biographical Memoir". http://newton.nap.edu/html/biomems/rbriggs.html. Retrieved 2006-04-22.
- ^ "Developing the potential of Xenopus tropicalis as a genetic model". http://grants.nih.gov/grants/guide/rfa-files/RFA-HD-01-008.html. Retrieved 2006-03-09.
- ^ "Joint Genome Institute - Xenopus tropicalis Home". http://genome.jgi-psf.org/Xentr4/Xentr4.home.html. Retrieved 2006-03-03.
- ^ Berrin, Katherine; Larco Museum (1997). The Spirit of Ancient Peru: Treasures from the Museo Arqueológico Rafael Larco Herrera. New York: Thames and Hudson. ISBN 0500018022.
- ^ Gratwicke, B (2009). "The Panamanian Golden Frog". Panama Amphibian Rescue and Conservation Project blog. http://amphibianrescue.org/?p=192.
Bibliography
- Beltz, Ellin (2005). Frogs: Inside their Remarkable World. Firefly Books. ISBN 1552978699.
- Cogger, H.G.; R.G. Zweifel, and D. Kirschner (2004). Encyclopedia of Reptiles & Amphibians Second Edition. Fog City Press. ISBN 1-877019-69-0.
- Estes, R., and O. A. Reig. (1973). "The early fossil record of frogs: a review of the evidence." pp. 11–63 In J. L. Vial (Ed.), Evolutionary Biology of the Anurans: Contemporary Research on Major Problems. University of Missouri Press, Columbia.
- Gissi, Carmela; Diego San Mauro, Graziano Pesole and Rafael Zardoya (February 2006). "Mitochondrial phylogeny of Anura (Amphibia): A case study of congruent phylogenetic reconstruction using amino acid and nucleotide characters". Gene 366 (2): 228–237. doi:10.1016/j.gene.2005.07.034. PMID 16307849.
- Holman, J. A (2004). Fossil Frogs and Toads of North America. Indiana University Press. ISBN 0-253-34280-5.
- San Mauro, Diego; Miguel Vences, Marina Alcobendas, Rafael Zardoya and Axel Meyer (May 2005). "Initial diversification of living amphibians predated the breakup of Pangaea". American Naturalist 165 (5): 590–599. doi:10.1086/429523. PMID 15795855. http://www.journals.uchicago.edu/doi/abs/10.1086/429523.
- Tyler, M. J. (1994). Australian Frogs A Natural History. Reed Books. ISBN 0-7301-0468-0.
External links
- AmphibiaWeb
- Gallery of Frogs - Photography and images of various species of frogs
- The Whole Frog Project - Virtual frog dissection and anatomy
- Disappearance of toads, frogs has some scientists worried San Francisco Chronicle, 20 April 1992
- Recording UK frogspawn sightings - Springwatch 2006
- Amphibian photo gallery by scientific name - Features many unusual frogs
- Scientific American: Researchers Pinpoint Source of Poison Frogs' Deadly Defenses
- Frogwatch USA - Volunteer frog and toad monitoring program by National Wildlife Federation and USGS, includes links to frog calls of the United States
- Media
- Time-lapse video showing the egg's development until hatching
- Frog calls - Short video clips of calling frogs and interviews with scientists about frog issues
- Frog vocalisations from around the world - From the British Library Sound Archive
- Frog calls - From Manitoba, Canada
- Frog calls - From eastern and central North America
Extant amphibian positions by subclass Lissamphibia Extant anuran families by suborder Archaeobatrachia 
Mesobatrachia Neobatrachia Amphignathodontidae · Aromobatidae · Arthroleptidae · Brachycephalidae · Bufonidae · Centrolenidae · Craugastoridae · Dendrobatidae · Heleophrynidae · Hemiphractidae · Hemisotidae · Hylidae · Hyperoliidae · Leptodactylidae · Mantellidae · Microhylidae · Myobatrachidae · Petropedetidae · Pyxicephalidae · Ranidae · Rhacophoridae · Rhinodermatidae · SooglossidaeCategories:- Frogs
- Animals that can change color
























Wikimedia Foundation. 2010.