- Cytoskeleton
-
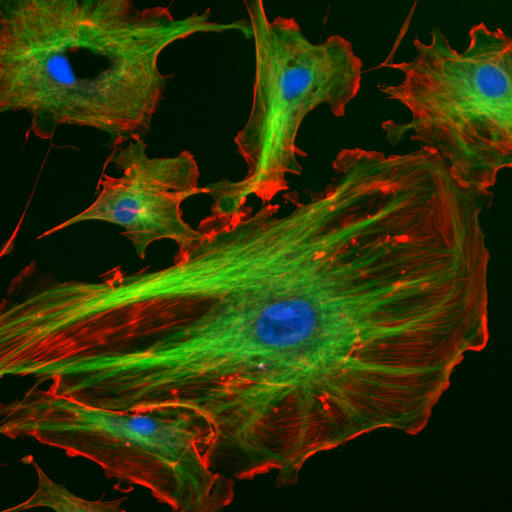
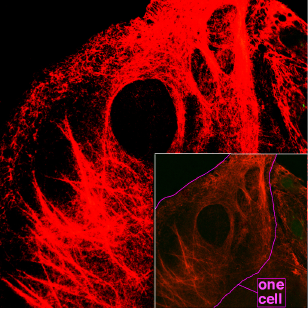
 The eukaryotic cytoskeleton. Actin filaments are shown in red, microtubules in green, and the nuclei are in blue.
The eukaryotic cytoskeleton. Actin filaments are shown in red, microtubules in green, and the nuclei are in blue.
The cytoskeleton (also CSK) is a cellular "scaffolding" or "skeleton" contained within a cell's cytoplasm and is made out of protein. The cytoskeleton is present in all cells; it was once thought to be unique to eukaryotes, but recent research has identified the prokaryotic cytoskeleton. It has structures such as flagella, cilia and lamellipodia and plays important roles in both intracellular transport (the movement of vesicles and organelles, for example) and cellular division. In 1903 Nikolai K Koltsov proposed that the shape of cells was determined by a network of tubules which he termed the cytoskeleton. The concept of a protein mosaic that dynamically coordinated cytoplasmic biochemistry was proposed by Rudolph Peters in 1929 [1] while the term (cytosquelette, in French) was first introduced by French embryologist Paul Wintrebert in 1931.[2]
Contents
The eukaryotic cytoskeleton
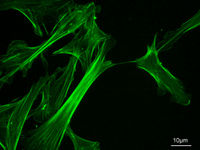
Eukaryotic cells contain three main kinds of cytoskeletal filaments, which are microfilaments, intermediate filaments, and microtubules. The cytoskeleton provides the cell with structure and shape, and by excluding macromolecules from some of the cytosol it adds to the level of macromolecular crowding in this compartment.[3] Cytoskeletal elements interact extensively and intimately with cellular membranes.[4]
Microfilaments (actin filaments)
These are the thinnest filaments of the cytoskeleton. They are composed of linear polymers of actin subunits, and generate force by elongation at one end of the filament coupled with shrinkage at the other, causing net movement of the intervening strand. They also act as tracks for the movement of myosin molecules that attach to the microfilament and "walk" along them. Actin structures are controlled by the Rho family of small GTP-binding proteins such as Rho itself for contractile acto-myosin filaments ("stress fibers"), Rac for lamellipodia and Cdc42 for filopodia.
Intermediate filaments
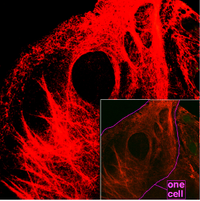 Microscopy of keratin filaments inside cells.
Microscopy of keratin filaments inside cells.These filaments, around 10 nanometers in diameter, are more stable (strongly bound) than actin filaments, and heterogeneous constituents of the cytoskeleton. Like actin filaments, they function in the maintenance of cell-shape by bearing tension (microtubules, by contrast, resist compression. It may be useful to think of micro- and intermediate filaments as cables, and of microtubules as cellular support beams). Intermediate filaments organize the internal tridimensional structure of the cell, anchoring organelles and serving as structural components of the nuclear lamina and sarcomeres. They also participate in some cell-cell and cell-matrix junctions.
Different intermediate filaments are:
- made of vimentins, being the common structural support of many cells.
- made of keratin, found in skin cells, hair and nails.
- neurofilaments of neural cells.
- made of lamin, giving structural support to the nuclear envelope.
Microtubules
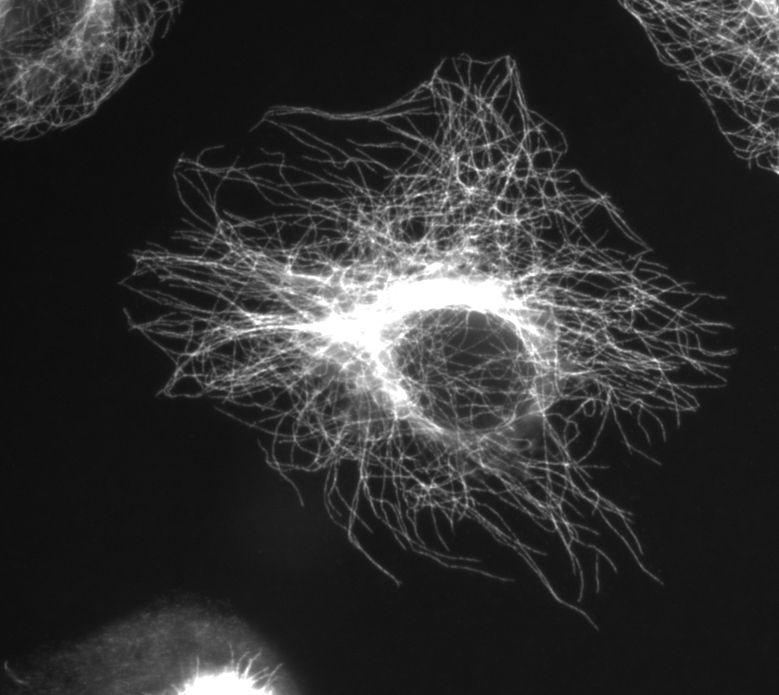
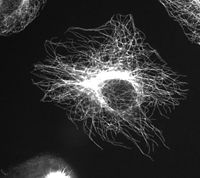 Microtubules in a gel fixated cell.
Microtubules in a gel fixated cell.Microtubules are hollow cylinders about 23 nm in diameter (lumen = approximately 15 nm in diameter), most commonly comprising 13 protofilaments which, in turn, are polymers of alpha and beta tubulin. They have a very dynamic behaviour, binding GTP for polymerization. They are commonly organized by the centrosome.
In nine triplet sets (star-shaped), they form the centrioles, and in nine doublets oriented about two additional microtubules (wheel-shaped) they form cilia and flagella. The latter formation is commonly referred to as a "9+2" arrangement, where in each doublet is connected to another by the protein dynein. As both flagella and cilia are structural components of the cell, and are maintained by microtubules, they can be considered part of the cytoskeleton.
They play key roles in:
- intracellular transport (associated with dyneins and kinesins, they transport organelles like mitochondria or vesicles).
- the axoneme of cilia and flagella.
- the mitotic spindle.
- synthesis of the cell wall in plants.
Comparison
Cytoskeleton type[5] Diameter (nm)[6] Structure Subunit examples[5] Microfilaments 6 double helix actin Intermediate filaments 10 two anti-parallel helices/dimers, forming tetramers - vimentin (mesenchyme)
- glial fibrillary acidic protein (glial cells)
- neurofilament proteins (neuronal processes)
- keratins (epithelial cells)
- nuclear lamins
Microtubules 23 protofilaments, in turn consisting of tubulin subunits α- and β-tubulin The prokaryotic cytoskeleton
The cytoskeleton was previously thought to be a feature only of eukaryotic cells, but homologues to all the major proteins of the eukaryotic cytoskeleton have recently been found in prokaryotes.[7] Although the evolutionary relationships are so distant that they are not obvious from protein sequence comparisons alone, the similarity of their three-dimensional structures and similar functions in maintaining cell shape and polarity provides strong evidence that the eukaryotic and prokaryotic cytoskeletons are truly homologous.[8] However, some structures in the bacterial cytoskeleton may have yet to be identified.[9]
FtsZ
FtsZ was the first protein of the prokaryotic cytoskeleton to be identified. Like tubulin, FtsZ forms filaments in the presence of GTP, but these filaments do not group into tubules. During cell division, FtsZ is the first protein to move to the division site, and is essential for recruiting other proteins that synthesize the new cell wall between the dividing cells.
MreB and ParM
Prokaryotic actin-like proteins, such as MreB, are involved in the maintenance of cell shape. All non-spherical bacteria have genes encoding actin-like proteins, and these proteins form a helical network beneath the cell membrane that guides the proteins involved in cell wall biosynthesis.
Some plasmids encode a partitioning system that involves an actin-like protein ParM. Filaments of ParM exhibit dynamic instability, and may partition plasmid DNA into the dividing daughter cells by a mechanism analogous to that used by microtubules during eukaryotic mitosis.
Crescentin
The bacterium Caulobacter crescentus contains a third 3rd protein, crescentin, that is related to the intermediate filaments of eukaryotic cells. Crescentin is also involved in maintaining cell shape, such as helical and vibrioid forms of bacteria, but the mechanism by which it does this is currently unclear.[10]
History
Microtrabeculae
A fourth eukaryotic cytoskeletal element, microtrabeculae, was proposed by Keith Porter based on images obtained from high-voltage electron microscopy of whole cells in the 1970s.[11] The images showed short, filamentous structures of unknown molecular composition associated with known cytoplasmic structures. Porter proposed that this microtrabecular structure represented a novel filamentous network distinct from microtubules, filamentous actin, or intermediate filaments. It is now generally accepted that microtrabeculae are nothing more than an artifact of certain types of fixation treatment, although we have yet to fully understand the complexity of the cell's cytoskeleton.[12]
References
- ^ Peters RA. The Harben Lectures, 1929. Reprinted in: Peters, R. A. (1963) Biochemical lesions and lethal synthesis,p. 216. Pergamon Press, Oxford..
- ^ Frixione E (June 2000). "Recurring views on the structure and function of the cytoskeleton: a 300-year epic". Cell motility and the cytoskeleton 46 (2): 73–94. doi:10.1002/1097-0169(200006)46:2<73::AID-CM1>3.0.CO;2-0. PMID 10891854.
- ^ Minton AP (October 1992). "Confinement as a determinant of macromolecular structure and reactivity". Biophys. J. 63 (4): 1090–100. Bibcode 1992BpJ....63.1090M. doi:10.1016/S0006-3495(92)81663-6. PMC 1262248. PMID 1420928. http://www.biophysj.org/cgi/reprint/63/4/1090.
- ^ Doherty GJ and McMahon HT (2008). "Mediation, Modulation and Consequences of Membrane-Cytoskeleton Interactions". Annual Review of Biophysics 37: 65–95. doi:10.1146/annurev.biophys.37.032807.125912. PMID 18573073. http://arjournals.annualreviews.org/doi/abs/10.1146/annurev.biophys.37.032807.125912.
- ^ a b Unless else specified in boxes, then ref is:Walter F., PhD. Boron (2003). Medical Physiology: A Cellular And Molecular Approaoch. Elsevier/Saunders. pp. 1300. ISBN 1-4160-2328-3. Page 25
- ^ Fuchs E, Cleveland DW (January 1998). "A structural scaffolding of intermediate filaments in health and disease". Science 279 (5350): 514–9. doi:10.1126/science.279.5350.514. PMID 9438837. http://www.sciencemag.org/cgi/pmidlookup?view=long&pmid=9438837.
- ^ Shih YL, Rothfield L (2006). "The bacterial cytoskeleton". Microbiol. Mol. Biol. Rev. 70 (3): 729–54. doi:10.1128/MMBR.00017-06. PMC 1594594. PMID 16959967. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1594594.
- ^ Michie KA, Löwe J (2006). "Dynamic filaments of the bacterial cytoskeleton". Annu. Rev. Biochem. 75: 467–92. doi:10.1146/annurev.biochem.75.103004.142452. PMID 16756499. http://www2.mrc-lmb.cam.ac.uk/SS/Lowe_J/group/PDF/annrev2006.pdf.[dead link]
- ^ Briegel A, Dias DP, Li Z, Jensen RB, Frangakis AS, Jensen GJ (October 2006). "Multiple large filament bundles observed in Caulobacter crescentus by electron cryotomography". Mol. Microbiol. 62 (1): 5–14. doi:10.1111/j.1365-2958.2006.05355.x. PMID 16987173.
- ^ Ausmees N, Kuhn JR, Jacobs-Wagner C (December 2003). "The bacterial cytoskeleton: an intermediate filament-like function in cell shape". Cell 115 (6): 705–13. doi:10.1016/S0092-8674(03)00935-8. PMID 14675535.
- ^ Wolosewick JJ, Porter KR (July 1979). "Microtrabecular lattice of the cytoplasmic ground substance. Artifact or reality". J. Cell Biol. 82 (1): 114–39. doi:10.1083/jcb.82.1.114. PMC 2110423. PMID 479294. http://www.jcb.org/cgi/pmidlookup?view=long&pmid=479294.
- ^ Heuser J (2002). "Whatever happened to the 'microtrabecular concept'?". Biol Cell 94 (9): 561–96. doi:10.1016/S0248-4900(02)00013-8. PMID 12732437.
External links
- Cytoskeleton, Cell Motility and Motors - The Virtual Library of Biochemistry and Cell Biology
- Cytoskeleton database, clinical trials, recent literature, lab registry ...
- Animation of leukocyte adhesion (Animation with some images of actin and microtubule assembly and dynamics.)
- http://cellix.imba.oeaw.ac.at/ Cytoskeleton and cell motility including videos
Structures of the cell / organelles (TH H1.00.01.2-3) Endomembrane system Cytoskeleton Endosymbionts Other internal External Proteins of the cytoskeleton Human I (MYO1A, MYO1B, MYO1C, MYO1D, MYO1E, MYO1F, MYO1G, MYO1H) · II (MYH1, MYH2, MYH3, MYH4, MYH6, MYH7, MYH7B, MYH8, MYH9, MYH10, MYH11, MYH13, MYH14, MYH15, MYH16) · III (MYO3A, MYO3B) · V (MYO5A, MYO5B, MYO5C) · VI (MYO6) · VII (MYO7A, MYO7B) · IX (MYO9A, MYO9B) · X (MYO10) · XV (MYO15A) · XVIII (MYO18A, MYO18B) · LC (MYL1, MYL2, MYL3, MYL4, MYL5, MYL6, MYL6B, MYL7, MYL9, MYLIP, MYLK, MYLK2, MYLL1)OtherOtherEpithelial keratins
(soft alpha-keratins)Ungrouped alphaNot alphaType 3Type 4Type 5OtherOtherNonhuman Categories:- Cell anatomy
- Cytoskeleton

Wikimedia Foundation. 2010.