- Insect
-
For the Breed 77 album, see Insects (album).
Insect
Temporal range: 396–0 Ma Early Devonian[1] (but see text) – Recent
Clockwise from top left: dancefly (Empis livida), long-nosed weevil (Rhinotia hemistictus), mole cricket (Gryllotalpa brachyptera), German wasp (Vespula germanica), emperor gum moth (Opodiphthera eucalypti), assassin bug (Harpactorinae) Scientific classification 
Kingdom: Animalia Phylum: Arthropoda Subphylum: Hexapoda Class: Insecta
Linnaeus, 1758Subclasses and orders - Dicondylia
Insects (from Latin insectum, a calque of Greek ἔντομον [éntomon], "cut into sections") are a class of living creatures within the arthropods that have a chitinous exoskeleton, a three-part body (head, thorax, and abdomen), three pairs of jointed legs, compound eyes, and two antennae. They are among the most diverse groups of animals on the planet, including more than a million described species and represent more than half of all known living organisms.[2][3] The number of extant species is estimated at between six and ten million,[2][4][5] and potentially represent over 90% of the differing metazoan life forms on Earth.[6] Insects may be found in nearly all environments, although only a small number of species occur in the oceans, a habitat dominated by another arthropod group, the crustaceans.
The life cycles of insects vary but most hatch from eggs. Insect growth is constrained by the inelastic exoskeleton and development involves a series of molts. The immature stages can differ from the adults in structure, habit and habitat and can include a passive pupal stage in those groups that undergo complete metamorphosis. Insects that undergo incomplete metamorphosis lack a pupal stage and adults develop through a series of nymphal stages.[7] The higher level relationship of the hexapoda is unclear. Fossilized insects of enormous size have been found from the Paleozoic Era, including giant dragonflies with wingspans of 55 to 70 cm (22–28 in). The most diverse insect groups appear to have coevolved with flowering plants.
Insects typically move about by walking, flying or occasionally swimming. As it allows for rapid yet stable movement, many insects adopt a tripedal gait in which they walk with their legs touching the ground in alternating triangles. Insects are the only invertebrates to have evolved flight. Many insects spend at least part of their life underwater, with larval adaptations that include gills and some adult insects are aquatic and have adaptations for swimming. Some species, like water striders, are capable of walking on the surface of water. Insects are mostly solitary, but some insects, such as certain bees, ants, and termites are social and live in large, well-organized colonies. Some insects, like earwigs, show maternal care, guarding their eggs and young. Insects can communicate with each other in a variety of ways. Male moths can sense the pheromones of female moths over distances of many kilometers. Other species communicate with sounds: crickets stridulate, or rub their wings together, to attract a mate and repel other males. Lampyridae in the beetle order Coleoptera communicate with light.
Humans regard certain insects as pests and attempt to control them using insecticides and a host of other techniques. Some insects damage crops by feeding on sap, leaves or fruits, a few bite humans and livestock, alive and dead, to feed on blood and some are capable of transmitting diseases to humans, pets and livestock. Nevertheless, without insects to pollinate flowers, the human race would soon run out of food because many of the crop plants that we rely on would not be able to reproduce.[8] Many other insects are considered ecologically beneficial as predators and a few provide direct economic benefit. Silkworms and bees have been used extensively by humans for the production of silk and honey, respectively.
Contents
Etymology
The word insect dates back to 1600, from the Latin word insectum meaning "with a notched or divided body," literally "cut into," from neuter plural of insectare "to cut into, to cut up," from in- "into" + secare "to cut";[9] from the fact that insects are "cut into" three sections. The word created by Pliny the Elder's loan-translation of the Greek word ἔντομος or "insect" (as in entomology), which was Aristotle's term for this class of life, also in reference to their "notched" bodies, first documented in English in 1601 in Holland's translation of Pliny. Translations of Aristotle's term also form the usual word for "insect" in Welsh (trychfil, from trychu "cut" + mil "animal"), Serbo-Croatian (zareznik, from rezati "cut"), Russian (насекомое nasekomoe, from sekat "cut"), etc.[9]
Distribution and diversity
Even though the true dimensions of species diversity remain uncertain, estimates are ranging from 1.4 to 1.8 million species. This probably represents less than 20% of all species on Earth, and with only about 20,000 new species of all organisms being described each year, it seems that most species will not remain described for many years unless there is a rapid increase in species descriptions. About 850,000–1,000,000 of all described species are insects. Of the 30 or so orders of insects, four dominate in terms of numbers of described species, with an estimated 600,000–795,000 species include Coleoptera, Diptera, Hymenoptera, and Lepidoptera. There are almost as many named species of beetle as there are of all other insects added together, or all other non-insects (plants and animals).
Comparison of the Estimated Number of Species for Vertebrates and the Four Most Species Order Described Species Average description rate (species per year) Publication effort Coleoptera 300,000–400,000 2308 0.01 Lepidoptera 110,000–120,000 642 0.03 Diptera 90,000–150,000 1048 0.04 Hymenoptera 100,000–125,000 1196 0.02 Morphology and physiology
Main articles: Insect morphology and Insect physiologyExternal
 Insect morphology
Insect morphology
A- Head B- Thorax C- Abdomen
1. antenna.
2. ocelli (lower)
3. ocelli (upper)
4. compound eye
5. brain (cerebral ganglia)
6. prothorax
7. dorsal blood vessel
8. tracheal tubes (trunk with spiracle)
9. mesothorax
10. metathorax
11. forewing
12. hindwing
13. mid-gut (stomach)
14. dorsal tube (Heart)
15. ovary
16. hind-gut (intestine, rectum & anus)
17. anus
18. oviduct
19. nerve chord (abdominal ganglia)
20. Malpighian tubes
21. tarsal pads
22. claws
23. tarsus
24. tibia
25. femur
26. trochanter
27. fore-gut (crop, gizzard)
28. thoracic ganglion
29. coxa
30. salivary gland
31. subesophageal ganglion
32. mouthpartsInsects have segmented bodies supported by an exoskeleton, a hard outer covering made mostly of chitin. The segments of the body are organized into three distinctive but interconnected units, or tagmata: a head, a thorax, and an abdomen.[10] The head supports a pair of sensory antennae, a pair of compound eyes, and, if present, one to three simple eyes (or ocelli) and three sets of variously modified appendages that form the mouthparts. The thorax has six segmented legs—one pair each for the prothorax, mesothorax and the metathorax segments making up the thorax—and, if present in the species, two or four wings. The abdomen consists of eleven segments, though in a few species of insects these segments may be fused together or reduced in size. The abdomen also contains most of the digestive, respiratory, excretory and reproductive internal structures.[11]:22–48 There is considerable variation and many adaptations in the body parts of insects especially wings, legs, antenna, mouth-parts etc.
Segmentation
The head: is enclosed in a hard, heavily sclerotized, unsegmented, exoskeletal head capsule, or epicranium, which contains most of the sensing organs, including the antennae, ocellus or eyes, and the mouthparts. Out of all the insect orders, Orthoptera displays the most features found in other insects, including the sutures and sclerites.[12] here, the vertex, or the apex (dorsal region), is situated between the compound eyes for insects with a hypognathous and opisthognathous head. In prognathous insects, the vertex is not found between the compound eyes, but rather, where the ocelli are normally. This is because the primary axis of the head is rotated 90 degrees to become parallel to the primary axis of the body. In some species this region is modified and assumes a different name.[12]:13
The thorax: is a segment composed of three sections, the prothorax, mesothorax, and the metathorax. The anterior segment, closest to the head, is the prothorax, with the major features being the first pair of legs and the pronotum. The middle segment is the mesothorax with the major features being the second pair of legs and the anterior wings. The third and most posterior segment, abutting the abdomen, is the metathorax, which features the third pair of legs and the posterior wings. Each segment is dilineated by an intersegmental suture. Each segment has four basic regions. The dorsal surface is called the tergum (or notum) to distinguish them from the abdominal terga.[11] The two lateral regions are called the pleura (singular: pleuron) and the ventral aspect is called the sternum. In turn, the notum of the prothorax is called the pronotum, the notum for the mesothorax is called the mesonotum and the notum for the metathorax is called the metanotum. Continuing with this logic, there is also the mesopleura and metapleura as well as the mesosternum and metasternum.[12]
The abdomen: is a the last segment of the insect, which typically consists of 11–12 segments and is less strongly sclerotized than the head or thorax. Each segment of the abdomen is represented by a sclerotized tergum, sternum, and perhaps a pleurite. Terga are separated from each other and from the adjacent sterna or pleura by a membrane. Spiracles are located in the pleural area. Variation of this ground plan includes the fusion of terga or terga and sterna to form continuous dorsal or ventral shields or a conical tube. Some insects bear a sclerite in the pleural area called a laterotergite. Ventral sclerites are sometimes called laterosternites. During the embryonic stage of many insects and the postembryonic stage of primitive insects, 11 abdominal segments are present. In modern insects there is a tendency toward reduction in the number of the abdominal segments, but the primitive number of 11 is maintained during embryogenesis.Variation in abdominal segment number is considerable. If the Apterygota are considered to be indicative of the ground plan for pterygotes, confusion reigns: adult Protura have 12 segments, Collembola have 6. The orthopteran family Acrididae has 11 segments, and a fossil specimen of Zoraptera has a 10-segmented abdomen.[12]
Exoskeleton
Insect outer skeleton, the cuticle, is made up of two layers: the epicuticle, which is a thin and waxy water resistant outer layer and contains no chitin, and a lower layer called the procuticle. The procuticle is chitinous and much thicker than the epicuticle and has two layers: an outer layer known as the exocuticle and an inner layer known as the endocuticle. The tough and flexible endocuticle is built from numerous layers of fibrous chitin and proteins, criss-crossing each others in a sandwich pattern, while the exocuticle is rigid and hardened.[11]:22–24 The exocuticle is greatly reduced in many soft-bodied insects (e.g., caterpillars), especially during their larval stages.
Insects are the only invertebrates to have developed active flight capability, and this has played an important role in their success.[11]:186 Their muscles are able to contract multiple times for each single nerve impulse, allowing the wings to beat faster than would ordinarily be possible. Having their muscles attached to their exoskeletons is more efficient and allows more muscle connections; crustaceans also use the same method, though all spiders use hydraulic pressure to extend their legs, a system inherited from their pre-arthropod ancestors. Unlike insects, though, most aquatic crustaceans are biomineralized with calcium carbonate extracted from the water.[13][14]
Internal
Nervous system
The nervous system of an insect can be divided into a brain and a ventral nerve cord. The head capsule is made up of six fused segments, each with a pair of ganglia, or a cluster of nerve cells outside of the brain. The first three pairs of ganglia are fused into the brain, while the three following pairs are fused into a structure of three pairs of ganglia under the insect's esophagus, called the subesophageal ganglion.[11]:57
The thoracic segments have one ganglion on each side, which are connected into a pair, one pair per segment. This arrangement is also seen in the abdomen but only in the first eight segments. Many species of insects have reduced numbers of ganglia due to fusion or reduction.[15] Some cockroaches have just six ganglia in the abdomen, whereas the wasp Vespa crabro has only two in the thorax and three in the abdomen. Some insects, like the house fly Musca domestica, have all the body ganglia fused into a single large thoracic ganglion.
At least a few insects have nociceptors, cells that detect and transmit sensations of pain.[16] This was discovered in 2003 by studying the variation in reactions of larvae of the common fruitfly Drosophila to the touch of a heated probe and an unheated one. The larvae reacted to the touch of the heated probe with a stereotypical rolling behavior that was not exhibited when the larvae were touched by the unheated probe.[17] Although nociception has been demonstrated in insects, there is not a consensus that insects feel pain consciously[18] but see Pain in invertebrates.
Digestive system
An insect uses its digestive system to extract nutrients and other substances from the food it consumes.[19] Most of this food is ingested in the form of macromolecules and other complex substances like proteins, polysaccharides, fats, and nucleic acids. These macromolecules must be broken down by catabolic reactions into smaller molecules like amino acids and simple sugars before being used by cells of the body for energy, growth, or reproduction. This break-down process is known as digestion.
The main structure of an insect's digestive system is a long enclosed tube called the alimentary canal, which runs lengthwise through the body. The alimentary canal directs food unidirectionally from the mouth to the anus. It has three sections, each of which performs a different process of digestion. In addition to the alimentary canal, insects also have paired salivary glands and salivary reservoirs. These structures usually reside in the thorax, adjacent to the foregut.[11]:70–77
The salivary glands (element 30 in numbered diagram) in an insect's mouth produce saliva. The salivary ducts lead from the glands to the reservoirs and then forward through the head to an opening called the salivarium, located behind the hypopharynx. By moving its mouthparts (element 32 in numbered diagram) the insect can mix its food with saliva. The mixture of saliva and food then travels through the salivary tubes into the mouth, where it begins to break down.[19][20] Some insects, like flies, have extra-oral digestion. Insects using extra-oral digestion expel digestive enzymes onto their food to break it down. This strategy allows insects to extract a significant proportion of the available nutrients from the food source.[21]:31 The gut is where almost all of insects' digestion takes place. It can be divided into the foregut, midgut and hindgut.
Foregut
 Stylized diagram of insect digestive tract showing malpighian tubule, from an insect of the order Orthoptera.
Stylized diagram of insect digestive tract showing malpighian tubule, from an insect of the order Orthoptera.The first section of the alimentary canal is the foregut (element 27 in numbered diagram), or stomodaeum. The foregut is lined with a cuticular lining made of chitin and proteins as protection from tough food. The foregut includes the buccal cavity (mouth), pharynx, esophagus, and Crop and proventriculus (any part may be highly modified) which both store food and signify when to continue passing onward to the midgut.[11]:70
Digestion starts in buccal cavity (mouth) as partially chewed food is broken down by saliva from the salivary glands. As the salivary glands produce fluid and carbohydrate-digesting enzymes (mostly amylases), strong muscles in the pharynx pump fluid into the buccal cavity, lubricating the food like the salivarium does, and helping blood feeders, and xylem and phloem feeders.
From there, the pharynx passes food to the esophagus, which could be just a simple tube passing it on to the crop and proventriculus, and then onward to the midgut, as in most insects. Alternately, the foregut may expand into a very enlarged crop and proventriculus, or the crop could just be a diverticulum, or fluid filled structure, as in some Diptera species.[21]:30–31
Midgut
Once food leaves the crop, it passes to the midgut (element 13 in numbered diagram), also known as the mesenteron, where the majority of digestion takes place. Microscopic projections from the midgut wall, called microvilli, increase the surface area of the wall and allow more nutrients to be absorbed; they tend to be close to the origin of the midgut. In some insects, the role of the microvilli and where they are located may vary. For example, specialized microvilli producing digestive enzymes may more likely be near the end of the midgut, and absorption near the origin or beginning of the midgut.[21]:32
Hindgut
In the hindgut (element 16 in numbered diagram), or proctodaeum, undigested food particles are joined by uric acid to form fecal pellets. The rectum absorbs 90% of the water in these fecal pellets, and the dry pellet is then eliminated through the anus (element 17), completing the process of digestion. The uric acid is formed using hemolymph waste products diffused from the Malpighian tubules (element 20). It is then emptied directly into the alimentary canal, at the junction between the midgut and hindgut. The number of Malpighian tubules possessed by a given insect varies between species, ranging from only two tubules in some insects to over 100 tubules in others.[11]:71–72, 78–80
Endocrine system
The salivary glands (element 30 in numbered diagram) in an insect's mouth produce saliva. The salivary ducts lead from the glands to the reservoirs and then forward through the head to an opening called the salivarium, located behind the hypopharynx. By moving its mouthparts (element 32 in numbered diagram) the insect can mix its food with saliva. The mixture of saliva and food then travels through the salivary tubes into the mouth, where it begins to break down.[19][22] Some insects, like flies, have extra-oral digestion. Insects using extra-oral digestion expel digestive enzymes onto their food to break it down. This strategy allows insects to extract a significant proportion of the available nutrients from the food source.[21]:31
Reproductive system
Respiratory and circulatory systems
Insect respiration is accomplished without lungs. Instead, the insect respiratory system uses a system of internal tubes and sacs through which gases either diffuse or are actively pumped, delivering oxygen directly to tissues that need it via their trachea (element 8 in numbered diagram). Since oxygen is delivered directly, the circulatory system is not used to carry oxygen, and is therefore greatly reduced. The insect circulatory system has no veins or arteries, and instead consists of little more than a single, perforated dorsal tube which pulses peristaltically. Toward the thorax, the dorsal tube (element 14) divides into chambers and acts like the insect's heart. The opposite end of the dorsal tube is like the aorta of the insect circulating the hemolymph, arthropods' fluid analog of blood, inside the body cavity.[11]:61–65[23] Air is taken in through openings on the sides of the abdomen called spiracles.
There are many different patterns of gas exchange demonstrated by different groups of insects. Gas exchange patterns in insects can range from continuous and diffusive ventilation, to discontinuous gas exchange.[11]:65–68 During continuous gas exchange, oxygen is taken in and carbon dioxide is released in a continuous cycle. In discontinuous gas exchange, however, the insect takes in oxygen while it is active and small amounts of carbon dioxide are released when the insect is at rest.[24] Diffusive ventilation is simply a form of continuous gas exchange that occurs by diffusion rather than physically taking in the oxygen. Some species of insect that are submerged also have adaptations to aid in respiration. As larvae, many insects have gills that can extract oxygen dissolved in water, while others need to rise to the water surface to replenish air supplies which may be held or trapped in special structures.[25][26]
Reproduction and development
 A pair of Simosyrphus grandicornis hoverflies mating in flight.
A pair of Simosyrphus grandicornis hoverflies mating in flight.The majority of insects hatch from eggs. The fertilization and development takes place inside the egg, enclosed by a shell (chorion). Some species of insects, like the cockroach Blaptica dubia, as well as juvenile aphids and tsetse flies, are ovoviviparous. The eggs of ovoviviparous animals develop entirely inside the female, and then hatch immediately upon being laid.[7] Some other species, such as those in the genus of cockroaches known as Diploptera, are viviparous, and thus gestate inside the mother and are born alive.[11]:129, 131, 134–135 Some insects, like parasitic wasps, show polyembryony, where a single fertilized egg divides into many and in some cases thousands of separate embryos.[11]:136–137
 The different forms of the male (top) and female (bottom) tussock moth Orgyia recens is an example of sexual dimorphism in insects.
The different forms of the male (top) and female (bottom) tussock moth Orgyia recens is an example of sexual dimorphism in insects.Other developmental and reproductive variations include haplodiploidy, polymorphism, paedomorphosis or peramorphosis, sexual dimorphism, parthenogenesis and more rarely hermaphroditism.[11]:143 In haplodiploidy, which is a type of sex-determination system, the offspring's sex is determined by the number of sets of chromosomes an individual receives. This system is typical in bees and wasps.[27] Polymorphism is the where a species may have different morphs or forms, as in the oblong winged katydid, which has four different varieties: green, pink, and yellow or tan. Some insects may retain phenotypes that are normally only seen in juveniles; this is called paedomorphosis. In peramorphosis, an opposite sort of phenomenon, insects take on previously unseen traits after they have matured into adults. Many insects display sexual dimorphism, in which males and females have notably different appearances, such as the moth Orgyia recens as an exemplar of sexual dimorphism in insects.
Some insects use parthenogenesis, a process in which the female can reproduce and give birth without having the eggs fertilized by a male. Many aphids undergo a form of parthenogenesis, called cyclical parthenogenesis, in which they alternate between one or many generations of asexual and sexual reproduction.[28][29] In summer, aphids are generally female and parthenogenetic; in the autumn, males may be produced for sexual reproduction. Other insects produced by parthenogenesis are bees, wasps, and ants, in which they spawn males. However, overall, most individuals are female, which are produced by fertilization. The males are haploid and the females are diploid.[7] More rarely, some insects display hermaphroditism, in which a given individual has both male and female reproductive organs.
Insect life-histories show adaptations to withstand cold and dry conditions. Some temperate region insects are capable of activity during winter, while some others migrate to a warmer climate or go into a state of torpor.[30] Still other insects have evolved mechanisms of diapause that allow eggs or pupae to survive these conditions.[31]
Metamorphosis
Metamorphosis in insects is the biological process of development all insects must undergo. There are two forms of metamorphosis: incomplete metamorphosis and complete metamorphosis.
Incomplete metamorphosis
Main article: HemimetabolismInsects that show hemimetabolism, or incomplete metamorphosis, change gradually by undergoing a series of molts. An insect molts when it outgrows its exoskeleton, which does not stretch and would otherwise restrict the insect's growth. The molting process begins as the insect's epidermis secretes a new epicuticle. After this new epicuticle is secreted, the epidermis releases a mixture of enzymes that digests the endocuticle and thus detaches the old cuticle. When this stage is complete, the insect makes its body swell by taking in a large quantity of water or air, which makes the old cuticle split along predefined weaknesses where the old exocuticle was thinnest.[11]:142[32] Other arthropods have a much different process and only molt; though must accommodate for the difference in exoskeleton structure and make up with other enzymes.
Immature insects that go through incomplete metamorphosis are called nymphs or in the case of dragonflies and damselflies as naiads. Nymphs are similar in form to the adult except for the presence of wings, which are not developed until adulthood. With each molt, nymphs grow larger and become more similar in appearance to adult insects.
 Like other insects that develop through incomplete metamorphosis, this Southern Hawker dragonfly molts its exoskeleton (shown above) several times during its pre-adult life.
Like other insects that develop through incomplete metamorphosis, this Southern Hawker dragonfly molts its exoskeleton (shown above) several times during its pre-adult life.
Complete metamorphosis
Main article: Holometabolism
Holometabolism, or complete metamorphosis, is where the insect changes all in four stages, an egg or embryo, a larva, a pupa, and the adult or imago. In these species, egg hatches to produce a larva, which is generally worm-like in form. This worm-like form can be one of several varieties: eruciform (caterpillar-like), scarabaeiform (grub-like), campodeiform (elongated, flattened, and active), elateriform (wireworm-like) or vermiform (maggot-like). The larva grows and eventually becomes a pupa, a stage marked by reduced movement and often sealed within a cocoon. There are three types of pupae: obtect, exarate or coarctate. Obtect pupae are compact, with the legs and other appendages enclosed. Exarate pupae have their legs and other appendages free and extended. Coarctate pupae develop inside the larval skin.[11]:151 Insects undergo considerable change in form during the pupal stage, and emerge as adults. Butterflies are a well known example of an insects that undergo complete metamorphosis, although most insects use this life cycle. Some insects have evolved this system to hypermetamorphosis.
Some of the oldest and most successful insect groups, such Endopterygota, use a system of complete metamorphosis.[11]:143 Strangely though, complete metamorphosis is unique to certain insect orders, like Diptera, Lepidoptera, and Hymenoptera, and no other arthropods undergo it, but incomplete metamorphosis.
Senses and communication
Many insects possess very sensitive and/or specialized organs of perception. Some insects such as bees can perceive ultraviolet wavelengths, or detect polarized light, while the antennae of male moths can detect the pheromones of female moths over distances of many kilometers.[33] There is a pronounced tendency for there to be a trade-off between visual acuity and chemical or tactile acuity, such that most insects with well-developed eyes have reduced or simple antennae, and vice-versa. There are a variety of different mechanisms by which insects perceive sound, while the patterns are not universal, insects can generally hear sound if they can produce it. Different insect species can have varying hearing, though most insects can hear only a narrow range of frequencies related to the frequency of the sounds they can produce. Mosquitoes have been found to hear up to 2 MHz., and some grasshoppers can hear up to 50 MHz.[34] Certain predatory and parasitic insects can detect the characteristic sounds made by their prey or hosts, respectively. For instance, some nocturnal moths can perceive the ultrasonic emissions of bats, which helps them avoid predation.[11]:87–94 Insects that feed on blood have special sensory structures that can detect infrared emissions, and use them to home in on their hosts.
Some insects display a rudimentary sense of numbers,[35] such as the solitary wasps that prey upon a single species. The mother wasp lays her eggs in individual cells and provides each egg with a number of live caterpillars on which the young feed when hatched. Some species of wasp always provide five, others twelve, and others as high as twenty-four caterpillars per cell. The number of caterpillars is different among species, but always the same for each sex of larva. The male solitary wasp in the genus Eumenes is smaller than the female, so the mother of one species supplies him with only five caterpillars; the larger female receives ten caterpillars in her cell.
Light production and vision
 Insects have compound eyes and two antennae.
Insects have compound eyes and two antennae.A few insects, such as members of the families Poduridae and Onychiuridae (Collembola), Mycetophilidae (Diptera), and the beetle families Lampyridae, Phengodidae, Elateridae and Staphylinidae are bioluminescent. The most familiar group are the fireflies, beetles of the family Lampyridae. Some species are able to control this light generation to produce flashes. The function varies with some species using them to attract mates, while others use them to lure prey. Cave dwelling larvae of Arachnocampa (Mycetophilidae, Fungus gnats) glow to lure small flying insects into sticky strands of silk.[36] Some fireflies of the genus Photuris mimic the flashing of female Photinus species to attract males of that species, which are then captured and devoured.[37] The colors of emitted light vary from dull blue (Orfelia fultoni, Mycetophilidae) to the familiar greens and the rare reds (Phrixothrix tiemanni, Phengodidae).[38]
Most insects, except some species of cave crickets, are able to perceive light and dark. Many species have acute vision capable of detecting minute movements. The eyes include simple eyes or ocelli as well as compound eyes of varying sizes. Many species are able to detect light in the infrared, ultraviolet and the visible light wavelengths. Color vision has been demonstrated in many species and phylogenetic analysis suggests that UV-green-blue trichromacy existed from at least the Devonian period between 416 and 359 million years ago.[39]
Sound production and hearing
Insects were the earliest organisms to produce and sense sounds. Insects make sounds mostly by mechanical action of appendages. In grasshoppers and crickets, this is achieved by stridulation. Cicadas make the loudest sounds among the insects by producing and amplifying sounds with special modifications to their body and musculature. The African cicada Brevisana brevis has been measured at 106.7 decibels at a distance of 50 cm (20 in).[40] Some insects, such as the hawk moths and Hedylid butterflies, can hear ultrasound and take evasive action when they sense that they have been detected by bats. Some moths produce ultrasonic clicks that were once thought to have a role in jamming bat echolocation. The ultrasonic clicks were subsequently found to be produced mostly by unpalatable moths to warn bats, just as warning colorations are used against predators that hunt by sight.[41] Some otherwise palatable moths have evolved to mimic these calls.[42]
Very low sounds are also produced in various species of Coleoptera, Hymenoptera, Lepidoptera, Mantodea, and Neuroptera. These low sounds are simply the sounds made by the insect's movement. Through microscopic stridulatory structures located on the insect's muscles and joints, the normal sounds of the insect moving are amplified and can be used to warn or communicate with other insects. Most sound-making insects also have tympanal organs that can perceive airborne sounds. Some species in Hemiptera, such as the corixids (water boatmen), are known to communicate via underwater sounds.[43] Most insects are also able to sense vibrations transmitted through surfaces. For example, an insect is caught in a spider web and struggles to escape. The vibrations it produces are sensed by the spider, who is alerted to its presence. Through these vibrations, the spider can tell where on the web the insect is located, as well as how big it is.[11]:87–94
Communication using surface-borne vibrational signals is more widespread among insects because of size constraints in producing air-borne sounds.[44] Insects cannot effectively produce low-frequency sounds, and high-frequency sounds tend to disperse more in a dense environment (such as foliage), so insects living in such environments communicate primarily using substrate-borne vibrations.[45] The mechanisms of production of vibrational signals are just as diverse as those for producing sound in insects.
Some species use vibrations for communicating within members of the same species, such as to attract mates as in the songs of the shield bug Nezara viridula.[46] Vibrations can also be used to communicate between entirely different species; lycaenid (gossamer-winged butterfly) caterpillars which are myrmecophilous (living in a mutualistic association with ants) communicate with ants in this way.[47] The Madagascar hissing cockroach has the ability to press air through its spiracles to make a hissing noise as a sign of aggression;[48] the Death's-head Hawkmoth makes a squeaking noise by forcing air out of their pharynx when agitated, which may also reduce aggressive worker honey bee behavior when the two are in close proximity.[49]
Chemical communication
In addition to the use of sound for communication, a wide range of insects have evolved chemical means for communication. These chemicals, termed semiochemicals, are often derived from plant metabolites include those meant to attract, repel and provide other kinds of information. Pheromones, a type of semiochemical, are used for attracting mates of the opposite sex, for aggregating conspecific individuals of both sexes, for deterring other individuals from approaching, to mark a trail, and to trigger aggression in nearby individuals. Allomonea benefit their producer by the effect they have upon the receiver. Kairomones benefit their receiver instead of their producer. Synomones benefit the producer and the receiver. While some chemicals are targeted at individuals of the same species, others are used for communication across species. The use of scents is especially well known to have developed in social insects.[11]:96–105
Social behavior
 A cathedral mound created by termites (Isoptera).
A cathedral mound created by termites (Isoptera).Social insects, such as termites, ants and many bees and wasps, are the most familiar species of eusocial animal.[50] They live together in large well-organized colonies that may be so tightly integrated and genetically similar that the colonies of some species are sometimes considered superorganisms. It is sometimes argued that the various species of honey bee are the only invertebrates (and indeed one of the few non-human groups) to have evolved a system of abstract symbolic communication where a behavior is used to represent and convey specific information about something in the environment. In this communication system, called dance language, the angle at which a bee dances represents a direction relative to the sun, and the length of the dance represents the distance to be flown.[11]:309–311
Only insects which live in nests or colonies demonstrate any true capacity for fine-scale spatial orientation or homing. This can allow an insect to return unerringly to a single hole a few millimeters in diameter among thousands of apparently identical holes clustered together, after a trip of up to several kilometers' distance. In a phenomenon known as philopatry, insects that hibernate have shown the ability to recall a specific location up to a year after last viewing the area of interest.[51] A few insects seasonally migrate large distances between different geographic regions (e.g., the overwintering areas of the Monarch butterfly).[11]:14
Care of young
Some bees, ants, termites and some wasps are eusocial, build nest, guard eggs and provide food for offsprings fulltime (see Eusociality). Most insects, however, lead short lives as adults, and rarely interact with one another except to mate or compete for mates. A small number exhibit some form of parental care, where they will at least guard their eggs, and sometimes continue guarding their offspring until adulthood, and possibly even feeding them. Another simple form of parental care is to construct a nest (a burrow or an actual construction, either of which may be simple or complex), store provisions in it, and lay an egg upon those provisions. The adult does not contact the growing offspring, but it nonetheless does provide food. This sort of care is typical for most species of bees and various types of wasps.[52]
Locomotion
Flight
Main article: Wing (insect)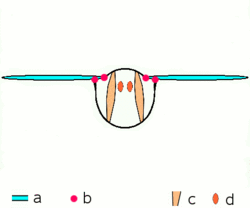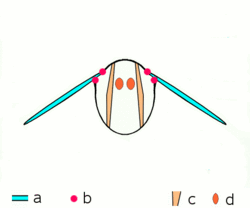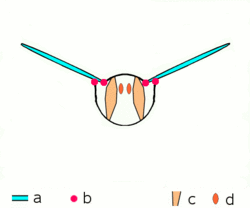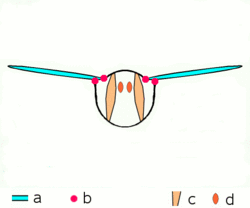 Basic motion of the insect wing in insect with an indirect flight mechanism scheme of dorsoventral cut through a thorax segment with
Basic motion of the insect wing in insect with an indirect flight mechanism scheme of dorsoventral cut through a thorax segment with
a wings
b joints
c dorsoventral muscles
d longitudinal muscles.Insects are the only group of invertebrates to have developed flight.[Note 1] The evolution of insect wings has been a subject of debate. Some entomologists suggest that the wings are from paranotal lobes, or extensions from the insect's exoskeleton called the nota, called the paranotal theory. Other theories are based on a pleural origin. These theories include suggestions that wings originated from modified gills, spiracular flaps or as from an appendage of the epicoxa. The epicoxal theory suggests the insect wings are modified epicoxal exites, a modified appendage at the base of the legs or coxa.[54] In the Carboniferous age, some of the Meganeura dragonflies had as much as a 50 cm (20 in) wide wingspan. The appearance of gigantic insects has been found to be consistent with high atmospheric oxygen. The respiratory system of insects constrains their size, however the high oxygen in the atmosphere allowed larger sizes.[55] The largest flying insects today are much smaller and include several moth species such as the Atlas moth and the White Witch (Thysania agrippina). Insect flight has been a topic of great interest in aerodynamics due partly to the inability of steady-state theories to explain the lift generated by the tiny wings of insects.
Unlike birds, many small insects are swept along by the prevailing winds[56] although many of the larger insects are known to make migrations. Aphids are known to be transported long distances by low-level jet streams.[57] As such, fine line patterns associated with converging winds within weather radar imagery, like the WSR-88D radar network, often represent large groups of insects.[58]
Walking
Many adult insects use six legs for walking and have adopted a tripedal gait. The tripedal gait allows for rapid walking while always having a stable stance and has been studied extensively in cockroaches. The legs are used in alternate triangles touching the ground. For the first step, the middle right leg and the front and rear left legs are in contact with the ground and move the insect forward, while the front and rear right leg and the middle left leg are lifted and moved forward to a new position. When they touch the ground to form a new stable triangle the other legs can be lifted and brought forward in turn and so on.[59] The purest form of the tripedal gait is seen in insects moving at high speeds. However, this type of locomotion is not rigid and insects can adapt a variety of gaits. For example, when moving slowly, turning, or avoiding obstacles, four or more feet may be touching the ground. Insects can also adapt their gait to cope with the loss of one or more limbs.
Cockroaches are among the fastest insect runners and, at full speed, adopt a bipedal run to reach a high velocity in proportion to their body size. As cockroaches move very quickly, they need to be video recorded at several hundred frames per second to reveal their gait. More sedate locomotion is seen in the stick insects or walking sticks (Phasmatodea). A few insects have evolved to walk on the surface of the water, especially the bugs of the Gerridae family, commonly known as water striders. A few species of ocean-skaters in the genus Halobates even live on the surface of open oceans, a habitat that has few insect species.[60]
Use in robotics
See also: Robot locomotion and Hexapod (robotics)Insect walking is of particular interest as an alternative form of locomotion in robots. The study of insects and bipeds has a significant impact on possible robotic methods of transport. This may allow new robots to be designed that can traverse terrain that robots with wheels may be unable to handle.[59]
Swimming
Main article: Aquatic insects The backswimmer Notonecta glauca underwater, showing its paddle-like hindleg adaptation.
The backswimmer Notonecta glauca underwater, showing its paddle-like hindleg adaptation.A large number of insects live either parts or the whole of their lives underwater. In many of the more primitive orders of insect, the immature stages are spent in an aquatic environment. Some groups of insects, like certain water beetles, have aquatic adults as well.[25]
Many of these species have adaptations to help in under-water locomotion. Water beetles and water bugs have legs adapted into paddle-like structures. Dragonfly naiads use jet propulsion, forcibly expelling water out of their rectal chamber.[61] Some species like the water striders are capable of walking on the surface of water. They can do this because their claws are not at the tips of the legs as in most insects, but recessed in a special groove further up the leg; this prevents the claws from piercing the water's surface film.[25] Other insects such as the Rove beetle Stenus are known to emit pygidial gland secretions that reduce surface tension making it possible for them to move on the surface of water by Marangoni propulsion (also known by the German term Entspannungsschwimmen).[62][63]
Phylogeny and systematics
 Evolution has produced astonishing variety in insects. Pictured are some of the possible shapes of antennae.Main article: Evolution of insects
Evolution has produced astonishing variety in insects. Pictured are some of the possible shapes of antennae.Main article: Evolution of insectsThe evolutionary relationships of insects to other animal groups remain unclear. Although more traditionally grouped with millipedes and centipedes, evidence has emerged favoring closer evolutionary ties with crustaceans. In the Pancrustacea theory, insects, together with Remipedia and Malacostraca, make up a natural clade.[64] Other terrestrial arthropods, such as centipedes, millipedes, scorpions and spiders, are sometimes confused with insects since their body plans can appear similar, sharing (as do all arthropods) a jointed exoskeleton. However, upon closer examination their features differ significantly; most noticeably they do not have the six legs characteristic of adult insects.[65]
Hexapoda (Insecta, Collembola, Diplura, Protura)
Myriapoda Diplopoda (millipedes)
Chilopoda (centipedes)
Chelicerata Eurypterida (sea scorpions: extinct)
Xiphosura (horseshoe crabs)
Pycnogonida (sea spiders)
Trilobites (extinct)
A phylogenetic tree of the arthropods and related groups[66]
The higher-level phylogeny of the arthropods continues to be a matter of debate and research. In 2008, researchers at Tufts University uncovered what they believe is the world's oldest known full-body impression of a primitive flying insect, a 300 million-year-old specimen from the Carboniferous Period.[67] The oldest definitive insect fossil is the Devonian Rhyniognatha hirsti, from the 396 million year old Rhynie chert. It may have superficially resembled a modern-day silverfish insect. This species already possessed dicondylic mandibles (two articulations in the mandible), a feature associated with winged insects, suggesting that wings may already have evolved at this time. Thus, the first insects probably appeared earlier, in the Silurian period.[1][68]
There have been four super radiations of insects: beetles (evolved ~300 million years ago), flies (evolved ~250 million years ago), moths and wasps (evolved ~150 million years ago).[citation needed] These four groups account for the majority of described species. The flies and moths along with the fleas evolved from the Mecoptera.
The origins of insect flight remain obscure, since the earliest winged insects currently known appear to have been capable fliers. Some extinct insects had an additional pair of winglets attaching to the first segment of the thorax, for a total of three pairs. As of 2009, there is no evidence that suggests that the insects were a particularly successful group of animals before they evolved to have wings.[69]
Late Carboniferous and Early Permian insect orders include both extant groups and a number of Paleozoic species, now extinct. During this era, some giant dragonfly-like forms reached wingspans of 55 to 70 cm (22 to 28 in) making them far larger than any living insect. This gigantism may have been due to higher atmospheric oxygen levels that allowed increased respiratory efficiency relative to today. The lack of flying vertebrates could have been another factor. Most extinct orders of insects developed during the Permian period that began around 270 million years ago. Many of the early groups became extinct during the Permian-Triassic extinction event, the largest mass extinction in the history of the Earth, around 252 million years ago.[70]
The remarkably successful Hymenopterans appeared as long as 146 million years ago in the Cretaceous period, but achieved their wide diversity more recently in the Cenozoic era, which began 66 million years ago. A number of highly successful insect groups evolved in conjunction with flowering plants, a powerful illustration of coevolution.[71]
Many modern insect genera developed during the Cenozoic. Insects from this period on are often found preserved in amber, often in perfect condition. The body plan, or morphology, of such specimens is thus easily compared with modern species. The study of fossilized insects is called paleoentomology.
Evolutionary relationships
Insects are prey for a variety of organisms, including terrestrial vertebrates. The earliest vertebrates on land existed 400 million years ago and were large amphibious piscivores, through gradual evolutionary change, insectivory was the next diet type to evolve.[72]
Insects were among the earliest terrestrial herbivores and acted as major selection agents on plants.[71] Plants evolved chemical defenses against this herbivory and the insects in turn evolved mechanisms to deal with plant toxins. Many insects make use of these toxins to protect themselves from their predators. Such insects often advertise their toxicity using warning colors.[71] This successful evolutionary pattern has also been utilized by mimics. Over time, this has led to complex groups of coevolved species. Conversely, some interactions between plants and insects, like pollination, are beneficial to both organisms. Coevolution has led to the development of very specific mutualisms in such systems.
Taxonomy
See also: Category:Orders of insects and Category:Insect familiesClassification Insecta Monocondylia -Archaeognatha - 470 Dicondylia Apterygota -Thysanura<200 -Monura Pterygota Paleoptera -Ephemeroptera- 2,500–<3,000 -Odonata- 6,500 Neoptera -Blattodea - 3,684–4,000 -Coleoptera - 360,000–400,000 -Dermaptera - 1,816 -Diptera - 152,956 -Embioptera - 200–300 -Grylloblattaria - 24 -Hymenoptera - 115,000 -Lepidoptera - 174,250 -Mantodea - 2,200 -Mecoptera - 481 -Megaloptera - 250–300 -Neuroptera - 5,000 -Orthoptera - 24,380 -Phasmatodea - 2,500–3,300 -Phthiraptera - 3,000–3,200 -Plecoptera - 2,274 -Siphonaptera - 2,525 -Strepsiptera - 596 -Trichoptera - 12,627 -Zoraptera - 28 -Zygentoma - 370 Cladogram of living insect groups,[73] with numbers of species in each group.[5] Note that Apterygota, Palaeoptera and Exopterygota are possibly paraphyletic groups. Traditional morphology-based or appearance-based systematics has usually given Hexapoda the rank of superclass,[11]:180 and identified four groups within it: insects (Ectognatha), springtails (Collembola), Protura and Diplura, the latter three being grouped together as Entognatha on the basis of internalized mouth parts. Supraordinal relationships have undergone numerous changes with the advent of methods based on evolutionary history and genetic data. A recent theory is that Hexapoda is polyphyletic (where the last common ancestor was not a member of the group), with the entognath classes having separate evolutionary histories from Insecta.[74] Many of the traditional appearance-based taxa have been shown to be paraphyletic, so rather than using ranks like subclass, superorder and infraorder, it has proved better to use monophyletic groupings (in which the last common ancestor is a member of the group). The following represents the best supported monophyletic groupings for the Insecta.
Insects can be divided into two groups historically treated as subclasses: wingless insects, known as Apterygota, and winged insects, known as Pterygota. The Apterygota consist of the primitively wingless order of the silverfish (Thysanura). Archaeognatha make up the Monocondylia based on the shape of their mandibles, while Thysanura and Pterygota are grouped together as Dicondylia. It is possible that the Thysanura themselves are not monophyletic, with the family Lepidotrichidae being a sister group to the Dicondylia (Pterygota and the remaining Thysanura).[75][76]
Paleoptera and Neoptera are the winged orders of insects differentiated by the presence of hardened body parts called sclerites; also, in Neoptera, muscles that allow their wings to fold flatly over the abdomen. Neoptera can further be divided into incomplete metamorphosis-based (Polyneoptera and Paraneoptera) and complete metamorphosis-based groups. It has proved difficult to clarify the relationships between the orders in Polyneoptera because of constant new findings calling for revision of the taxa. For example, Paraneoptera has turned out to be more closely related to Endopterygota than to the rest of the Exopterygota. The recent molecular finding that the traditional louse orders Mallophaga and Anoplura are derived from within Psocoptera has led to the new taxon Psocodea.[77] Phasmatodea and Embiidina have been suggested to form Eukinolabia.[78] Mantodea, Blattodea and Isoptera are thought to form a monophyletic group termed Dictyoptera.[79]
It is likely that Exopterygota is paraphyletic in regard to Endopterygota. Matters that have had a lot of controversy include Strepsiptera and Diptera grouped together as Halteria based on a reduction of one of the wing pairs – a position not well-supported in the entomological community.[80] The Neuropterida are often lumped or split on the whims of the taxonomist. Fleas are now thought to be closely related to boreid mecopterans.[81] Many questions remain to be answered when it comes to basal relationships amongst endopterygote orders, particularly Hymenoptera.
The study of the classification or taxonomy of any insect is called systematic entomology. If one works with a more specific order or even a family, the term may also be made specific to that order or family, for example systematic dipterology.
Ecology
Many insects, especially beetles, are scavengers that feed on dead animals and fallen trees and thereby recycle biological materials into forms found useful by other organisms. Insects are responsible for much of the process by which topsoil is created.[11]:3, 218–228
Defense and predation
Pollination
Mutualism
Parasitism
Other biological interactions
Relationship to humans
As pests

Many insects are considered pests by humans. Insects commonly regarded as pests include those that are parasitic (mosquitoes, lice, bed bugs), transmit diseases (mosquitoes, flies), damage structures (termites), or destroy agricultural goods (locusts, weevils). Many entomologists are involved in various forms of pest control, as in research for companies to produce insecticides, but increasingly relying on methods of biological pest control, or biocontrol. Biocontrol uses one organism to reduce the population density of another organism — the pest — and is considered a key element of integrated pest management.[82][83]
Despite the large amount of effort focused at controlling insects, human attempts to kill pests with insecticides can backfire. If used carelessly the poison can kill all kinds of organisms in the area, including insects' natural predators such as birds, mice, and other insectivores. The effects of DDT's use exemplifies how some insecticides can threaten wildlife beyond intended populations of pest insects.[84][85]
As beneficial
 Because they help flowering plants to cross-pollinate, some insects are critical to agriculture. This European honey bee is gathering nectar while pollen collects on its body.
Because they help flowering plants to cross-pollinate, some insects are critical to agriculture. This European honey bee is gathering nectar while pollen collects on its body.Although pest insects attract the most attention, many insects are beneficial to the environment and to humans. Some insects, like wasps, bees, butterflies, and ants, pollinate flowering plants. Pollination is a mutualistic relationship between plants and insects. As insects gather nectar from different plants of the same species, they also spread pollen from plants on which they have previously fed. This greatly increases plants' ability to cross-pollinate, which maintains and possibly even improves their evolutionary fitness. This ultimately affects humans since ensuring healthy crops is critical to agriculture. A serious environmental problem is the decline of populations of pollinator insects, and a number of species of insects are now cultured primarily for pollination management in order to have sufficient pollinators in the field, orchard or greenhouse at bloom time.[86]:240–243 Insects also produce useful substances such as honey, wax, lacquer and silk. Honey bees have been cultured by humans for thousands of years for honey, although contracting for crop pollination is becoming more significant for beekeepers. The silkworm has greatly affected human history, as silk-driven trade established relationships between China and the rest of the world.
 A robberfly with its prey, a hoverfly. Insectivorous relationships such as these help control insect populations.
A robberfly with its prey, a hoverfly. Insectivorous relationships such as these help control insect populations.Insectivorous insects, or insects which feed on other insects, are beneficial to humans because they eat insects that could cause damage to agriculture and human structures. For example, aphids feed on crops and cause problems for farmers, but ladybugs feed on aphids, and can be used as a means to get significantly reduce pest aphid populations. While birds are perhaps more visible predators of insects, insects themselves account for the vast majority of insect consumption. Without predators to keep them in check, insects can undergo almost unstoppable population explosions.[11]:328–348[11]:400[87][88]
Insects are also used in medicine, for example fly larvae (maggots) were formerly used to treat wounds to prevent or stop gangrene, as they would only consume dead flesh. This treatment is finding modern usage in some hospitals. Recently insects have also gained attention as potential sources of drugs and other medicinal substances.[89] Also adult insects, such as crickets, and insect larvae of various kinds are also commonly used as fishing bait.[90]
In research
 The common fruitfly Drosophila melanogaster is one of the most widely used organisms in biological research.
The common fruitfly Drosophila melanogaster is one of the most widely used organisms in biological research.Insects play important roles in biological research. For example, because of its small size, short generation time and high fecundity, the common fruit fly Drosophila melanogaster is a model organism for studies in the genetics of higher eukaryotes. D. melanogaster has been an essential part of studies into principles like genetic linkage, interactions between genes, chromosomal genetics, development, behavior, and evolution. Because genetic systems are well conserved among eukaryotes, understanding basic cellular processes like DNA replication or transcription in fruit flies can help to understand those processes in other eukaryotes, including humans.[91] The genome of D. melanogaster was sequenced in 2000, reflecting the organism's important role in biological research.[92]
As food
Main article: EntomophagyIn some parts of the world, insects are used for human food, while being a taboo in other places. In some cultures, insects, especially deep-fried cicadas, are considered to be delicacies, while in other places they form part of the normal diet as they have a high protein content for their mass. In most first-world countries, however, entomophagy, or the consumption of insects, is taboo.[93] There are proponents of developing this use to provide a major source of protein in human nutrition.[11]:10–13 Since it is impossible to entirely eliminate pest insects from the human food chain, insects are present in many foods, especially grains. Food safety laws in many countries do not prohibit insect parts in food, but rather limit the quantity. According to cultural materialist anthropologist Marvin Harris, the eating of insects is taboo in cultures that have other protein sources such as fish or livestock.
In culture
Scarab beetles held religious and cultural symbolism in Old Egypt, Greece and some shamanistic Old World cultures. The ancient Chinese regarded cicadas as symbols of rebirth or immortality. In Mesopotamian literature, the epic poem of Gilgamesh has allusions to Odonata which signify the impossibility of immortality. Amongst the Aborigines of Australia of the Arrernte language groups, honey ants and witchety grubs served as personal clan totems. In the case of the 'San' bush-men of the Kalahari, it is the praying mantis which holds much cultural significance including creation and zen-like patience in waiting.[11]:9
See also
- Entomology
- Flying and gliding animals
- Category: Insect-borne diseases
- Prehistoric insects
- Ecological importance of bees
- Insect ecology
- Chemical ecology
- Insect biodiversity
- Defense in insects
Notes
References
- ^ a b Engel, Michael S.; David A. Grimaldi (2004). "New light shed on the oldest insect". Nature 427 (6975): 627–630. doi:10.1038/nature02291. PMID 14961119. http://www.nature.com/nature/journal/v427/n6975/full/nature02291.html.
- ^ a b Chapman, A. D. (2006). Numbers of living species in Australia and the World. Canberra: Australian Biological Resources Study. pp. 60pp. ISBN 978-0-642-56850-2. http://www.deh.gov.au/biodiversity/abrs/publications/other/species-numbers/index.html.
- ^ Wilson, E.O.. "Threats to Global Diversity". http://www.globalchange.umich.edu/globalchange2/current/lectures/biodiversity/biodiversity.html. Retrieved 2009-05-17.
- ^ Vojtech Novotny, Yves Basset, Scott E. Miller, George D. Weiblen, Birgitta Bremer, Lukas Cizek & Pavel Drozd (2002). "Low host specificity of herbivorous insects in a tropical forest". Nature 416 (6883): 841–844. doi:10.1038/416841a. PMID 11976681.
- ^ a b Erwin, Terry L. (1997). Biodiversity at its utmost: Tropical Forest Beetles. pp. 27–40. In: Reaka-Kudla, M. L., D. E. Wilson & E. O. Wilson (eds.). Biodiversity II. Joseph Henry Press, Washington, D.C..
- ^ Erwin, Terry L. (1982). "Tropical forests: their richness in Coleoptera and other arthropod species". Coleopt. Bull. 36: 74–75.
- ^ a b c "insect physiology" McGraw-Hill Encyclopedia of Science and Technology, Ch. 9, p 233, 2007
- ^ http://www.britannica.com/EBchecked/topic/289001/insect
- ^ a b Harpe, Douglas; Dan McCormack (November 2001). "Online Etymological Dictionary". Online Etymological Dictionary. LogoBee.com. p. 1. http://www.etymonline.com/index.php?term=insect&allowed_in_frame=0. Retrieved November 1, 2011.
- ^ "O. Orkin Insect zoo". The University of Nebraska Department of Entomology. http://insectzoo.msstate.edu/Students/basic.structure.html. Retrieved 2009-05-03.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa Gullan, P.J.; P.S. Cranston (2005). The Insects: An Outline of Entomology (3 ed.). Oxford: Blackwell Publishing. ISBN 1-4051-1113-5.
- ^ a b c d Resh, Vincent H.; Ring T. Carde (July 1, 2009). Encyclopedia of Insects (2 ed.). U. S. A.: Academic Press. ISBN 0123741440.
- ^ Barnes, R.S.K., Calow, P., Olive, P., Golding, D., and Spicer, J. (2001). "Invertebrates with Legs: the Arthropods and Similar Groups". The Invertebrates: A Synthesis. Blackwell Publishing. p. 168. ISBN 0-632-04761-5. http://books.google.com/?id=TBMsbe9efPgC&pg=PA168&dq=arthropod+hydraulic. Retrieved 2008-09-25.
- ^ Lowenstam, H.A., and Weiner, S. (1989). On biomineralization. Oxford University Press US. p. 111. ISBN 0-19-504977-2. http://books.google.com/?id=JbAgy0AAopsC&dq=arthropod+biomineralization. Retrieved 2008-09-25.
- ^ Schneiderman, Howard A. (1960). "Discontinuous respiration in insects: role of the spiracles". Biol. Bull. 119 (119): 494–528. doi:10.2307/1539265. JSTOR 1539265. http://www.biolbull.org/cgi/reprint/119/3/494?maxtoshow=&HITS=10&hits=10&RESULTFORMAT=&fulltext=insect+thoracic+spiracle&andorexactfulltext=and&searchid=1&FIRSTINDEX=0&sortspec=relevance&resourcetype=HWCIT.
- ^ Eisemann, WK et al. (1984). "Do insects feel pain? — A biological view". Cellular and Molecular Life Sciences 40: 1420–1423.
- ^ Tracey, J et al.; Wilson, RI; Laurent, G; Benzer, S (18 April 2003). "painless, a Drosophila gene essential for nociception". Cell 113 (2): 261–273. doi:10.1016/S0092-8674(03)00272-1. PMID 12705873.
- ^ Sømme, LS (14 January 2005). "Sentience and pain in invertebrates". Norwegian Scientific Committee for Food Safety. http://google.com/scholar?q=cache:dbrbM20yEQ4J:scholar.google.com/&hl=en. Retrieved September 30, 2009.
- ^ a b c "General Entomology - Digestive and Excritory system". NC state University. http://www.cals.ncsu.edu/course/ent425/library/tutorials/internal_anatomy/digestive.html. Retrieved 2009-05-03.
- ^ Duncan, Carl D. (1939). A Contribution to The Biology of North American Vespine Wasps (1 ed.). Stanford: Stanford University Press. pp. 24–29.
- ^ a b c d Nation, James L. (November 2001). "15". Insect Physiology and Biochemistry (1 ed.). CRC Press. pp. 496pp. ISBN 0-8493-1181-0. http://books.google.com/?id=l3v2tOvz1uQC&pg=PA31&dq=insect+foregut#v=onepage&q=insect%20foregut.
- ^ Duncan, Carl D. (1939). A Contribution to The Biology of North American Vespine Wasps (1 ed.). Stanford: Stanford University Press. pp. 24–29.
- ^ Meyer, John R. (17 February 2006). "Circulatory System". NC State University: Department of Entomology, NC State University. pp. 1. http://www.cals.ncsu.edu/course/ent425/tutorial/circulatory.html. Retrieved 2009-10-11.
- ^ Chown, S.L.; S.W. Nicholson (2004). Insect Physiological Ecology. New York: Oxford University Press. ISBN 0-19-851549-9.
- ^ a b c Richard W. Merritt, Kenneth W. Cummins, and Martin B. Berg (editors) (2007). An Introduction to the Aquatic Insects of North America (4th ed.). Kendall Hunt Publishers. ISBN 978-0-7575-5049-2.
- ^ Merritt, RW, KW Cummins, and MB Berg (2007). An Introduction To The Aquatic Insects Of North America. Kendall Hunt Publishing Company. ISBN 0-7575-4128-3.
- ^ William O. H. Hughes, Benjamin P. Oldroyd, Madeleine Beekman, Francis L. W. Ratnieks (2008-05-30). "Ancestral Monogamy Shows Kin Selection Is Key to the Evolution of Eusociality". Science (American Association for the Advancement of Science) 320 (5880): 1213–1216. Bibcode 2008Sci...320.1213H. doi:10.1126/science.1156108. PMID 18511689. http://www.sciencemag.org/cgi/content/abstract/320/5880/1213. Retrieved 2008-08-04.
- ^ Effect of nitrogen fertilization on Aphis gossypii (Homoptera: Aphididae): variation in size, color, and reproduction, E. Nevo and M. Coll, J. Econ. Entomol. 94: 27–32, 2001.
- ^ Effect of nitrogen fertilizer on the intrinsic rate of increase of the rusty plum aphid, Hysteroneura setariae (Thomas) (Homoptera: Aphididae) on rice (Oryza sativa L.), G. C. Jahn, L. P. Almazan, and J. Pacia, Environmental Entomology 34 (4): 938–943, 2005.
- ^ http://insects.about.com/od/adaptations/p/wintersurvival.htm Where Do Insects Go in Winter?
- ^ Lee, Richard E, Jr. (1989). "Insect Cold-Hardiness: To Freeze or Not to Freeze". BioScience 39 (5): 308–313. doi:10.2307/1311113. JSTOR 1311113. http://www.units.muohio.edu/cryolab/publications/documents/Lee89_BioSci.pdf.
- ^ Ruppert, E.E., Fox, R.S., and Barnes, R.D. (2004). Invertebrate Zoology (7 ed.). Brooks / Cole. pp. 523–524. ISBN 0-03-025982-7.
- ^ "Insects". Alien Life Forms. pp. 4. http://crazydaz.com/insects.pdf. Retrieved 2009-05-17.
- ^ Cator, L. J. et al.; Arthur, BJ; Harrington, LC; Hoy, RR (20 February 2009). "Harmonic Convergence in the Love Songs of the Dengue Vector Mosquito". Science 323 (5917): 1077–1079. Bibcode 2009Sci...323.1077C. doi:10.1126/science.1166541. PMC 2847473. PMID 19131593. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2847473.
- ^ Möller, R. (2002) (in German). A Biorobotics Approach to the Study of Insect Visual Homing Strategies. p. 11. http://www.ti.uni-bielefeld.de/downloads/publications/habil.pdf.
- ^ Pugsley, Chris W. (1983). "Literature review of the New Zealand glowworm Arachnocampa luminosa (Diptera: Keroplatidae) and related cave-dwelling Diptera". New Zealand Entomologist 7 (4): 419–424. http://www.ento.org.nz/nzentomologist/free_issues/NZEnto07_4_1983/Volume%207-4-419-424.pdf.
- ^ Lloyd, James E. (1984). "Occurrence of Aggressive Mimicry in Fireflies". The Florida Entomologist 67 (3): 368–376. doi:10.2307/3494715. JSTOR 3494715.
- ^ Lloyd, James E.; Erin C. Gentry (2003). The Encyclopedia of Insects. Academic Press. pp. 115–120. ISBN 0-12-586990-8.
- ^ Briscoe, AD & L Chittka (2001). "The evolution of color vision in insects". Annu. Rev. Entomol 46: 471–510. doi:10.1146/annurev.ento.46.1.471. PMID 11112177.
- ^ Walker, T.J., ed. 2001. University of Florida Book of Insect Records, 2001. [1]
- ^ Hristov, N.I.; Conner, W.E. (2005). "Sound strategy: acoustic aposematism in the bat–tiger moth arms race". Naturwissenschaften 92 (4): 164–169. Bibcode 2005NW.....92..164H. doi:10.1007/s00114-005-0611-7. PMID 15772807.
- ^ Barber, J. R.; W. E. Conner (2007). "Acoustic mimicry in a predator–prey interaction". Proc. Nat. Acad. Sci. 104 (22): 9331–9334. Bibcode 2007PNAS..104.9331B. doi:10.1073/pnas.0703627104. PMC 1890494. PMID 17517637. http://www.pnas.org/cgi/content/figsonly/104/22/9331.
- ^ Theiss, Joachim (1982). "Generation and radiation of sound by stridulating water insects as exemplified by the corixids". Behavioral Ecology and Sociobiology 10: 225–235. doi:10.1007/BF00299689.
- ^ Virant-Doberlet, M.; Čokl A. (2004). "Vibrational communication in insects". Neotropical Entomology 33 (2): 121–134. doi:10.1590/S1519-566X2004000200001. http://www.scielo.br/pdf/ne/v33n2/a01v33n2.pdf.
- ^ Bennet-Clark, H.C. (1998). "Size and scale effects as constraints in insect sound communication". Phil. Trans. R. Soc. Lond. B 353: 407–419. doi:10.1098/rstb.1998.0219.
- ^ Miklas, Nadège; Nataša Stritih, Andrej Čokl, Meta Virant-Doberlet, Michel Renou (2001). "The Influence of Substrate on Male Responsiveness to the Female Calling Song in Nezara viridula". Journal of Insect Behavior 14 (3): 313–332. doi:10.1023/A:1011115111592.
- ^ DeVries, P. J. (1990). "Enhancement of symbiosis between butterfly caterpillars and ants by vibrational communication". Science 248 (4959): 1104–1106. Bibcode 1990Sci...248.1104D. doi:10.1126/science.248.4959.1104. PMID 17733373.
- ^ Nelson, Margaret C.; Jean Fraser (1980). "Sound production in the cockroach, Gromphadorhina portentosa: evidence for communication by hissing". Behavioral Ecology and Sociobiology 6 (4): 305–314. doi:10.1007/BF00292773.
- ^ Moritz, R. F. A.; W. H. Kirchner and R. M. Crewe (April 1991). "Chemical camouflage of the death's head hawkmoth (Acherontia atropos L.) in honeybee colonies". Naturwissenschaften 78 (4): 179–182. Bibcode 1991NW.....78..179M. doi:10.1007/BF01136209.
- ^ Brewer, Gary. "Social insects". North Dakota State University. Archived from the original on March 21, 2008. http://web.archive.org/web/20080321171246/http://www.ndsu.nodak.edu/entomology/topics/societies.htm. Retrieved 2009-05-06.
- ^ Salt, R.W. (January 1961). "Principles of Insect Cold-Hardiness". Annual Review of Entomology (Lethbirdge, Alberta, Canada: Canada Agriculture Research Station) 6: 55. doi:10.1146/annurev.en.06.010161.000415. http://arjournals.annualreviews.org/doi/abs/10.1146/annurev.en.06.010161.000415.
- ^ "Social Insects". North Dakota State University. Archived from the original on March 21, 2008. http://web.archive.org/web/20080321171246/http://www.ndsu.nodak.edu/entomology/topics/societies.htm. Retrieved 2009-10-12.
- ^ Ferris Jabr (2010). "Fact or Fiction: Can a Squid Fly Out of the Water?". Scientific American. http://www.scientificamerican.com/article.cfm?id=can-squid-fly.
- ^ Jockusch, EL; Ober, KA (September 2004). "Hypothesis testing in evolutionary developmental biology: a case study from insect wings". Journal of Heredity 95 (5): 382–396. doi:10.1093/jhered/esh064. PMID 15388766.
- ^ Dudley, R (1998). "Atmospheric oxygen, giant Paleozoic insects and the evolution of aerial locomotor performance". Journal of Experimental Biology 201 (8): 1043–1050. http://jeb.biologists.org/cgi/reprint/201/8/1043.pdf.
- ^ Diana Yates (2008). Birds migrate together at night in dispersed flocks, new study indicates. University of Illinois at Urbana - Champaign. Retrieved on 2009-04-26.
- ^ Drake, V. A.; R. A. Farrow (1988). "The Influence of Atmospheric Structure and Motions on Insect Migration". Annual Review of Entomology 33: 183–210. doi:10.1146/annurev.en.33.010188.001151.
- ^ Bart Geerts and Dave Leon (2003). P5A.6 Fine-Scale Vertical Structure of a Cold Front As Revealed By Airborne 95 GHZ Radar. University of Wyoming. Retrieved on 2009-04-26.
- ^ a b Biewener, Andrew A (2003). Animal Locomotion. Oxford University Press. ISBN 0-19-850022-X.
- ^ Ikawa, Terumi,; Okabe, Hidehiko; Hoshizaki, Sugihiko; Kamikado, Takahiro; Cheng, Lanna (2004). "Distribution of the oceanic insects Halobates (Hemiptera: Gerridae) off the south coast of Japan". Entomological Science 7 (4): 351–357. doi:10.1111/j.1479-8298.2004.00083.x.
- ^ Mill, P. J.; R. S. Pickard (1975). "Jet-propulsion in anisopteran dragonfly larvae". Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology 97 (4): 329–338. doi:10.1007/BF00631969.
- ^ Linsenmair, K.; Jander R. (1976). "Das "entspannungsschwimmen" von Velia and Stenus". Naturwissenschaften 50: 231. Bibcode 1963NW.....50..231L. doi:10.1007/BF00639292.
- ^ Bush, J. W. M.; David L. Hu (2006). "Walking on Water: Biolocomotion at the Interface". Annu. Rev. Fluid Mech. 38: 339–369. Bibcode 2006AnRFM..38..339B. doi:10.1146/annurev.fluid.38.050304.092157. http://www.cims.nyu.edu/~dhu/Pubweb/Bush_Hu_06.pdf.
- ^ "Palaeos invertebrates:Arthropoda". Palaeos Invertebrates. 2002-05-03. http://www.palaeos.com/Invertebrates/Arthropods/Pancrustacea.html. Retrieved 2009-05-06.
- ^ "Evolution of insect flight". Malcolm W. Browne. 1994-10-25. http://dml.cmnh.org/1994Oct/msg00116.html. Retrieved 2009-05-06.
- ^ "Tree of Life Web Project. Version 1 January 1995 (temporary) of Arthropoda". Tree of Life Web Project. 1995. http://www.tolweb.org/Arthropoda. Retrieved 2009-05-09.
- ^ "Researchers Discover Oldest Fossil Impression of a Flying Insect". Newswise. http://newswise.com/articles/view/545296/. Retrieved 2008-19-20.
- ^ Rice, C. M., Ashcroft, W. A., Batten, D. J., Boyce, A. J., Caulfield, J. B. D., Fallick, A. E., Hole, M. J., Jones, E., Pearson, M. J., Rogers, G., Saxton, J. M., Stuart, F. M., Trewin, N. H. & Turner, G. (1995). "A Devonian auriferous hot spring system, Rhynie, Scotland". Journal of the Geological Society, London 152: 229–250. doi:10.1144/gsjgs.152.2.0229.
- ^ Grimaldi, D. and Engel, M.S. (2005). Evolution of the Insects. Cambridge University Press. ISBN 0-521-82149-5.
- ^ Rasnitsyn, A.P. and Quicke, D.L.J. (2002). History of Insects. Kluwer Academic Publishers. ISBN 1-4020-0026-X.
- ^ a b c J. Stein Carter (2005-03-29). "Coevolution and Pollination". University of Cincinnati. http://biology.clc.uc.edu/courses/bio303/coevolution.htm. Retrieved 2009-05-09.
- ^ Sahney, S., Benton, M.J. & Falcon-Lang, H.J. (2010). "Rainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica" (PDF). Geology 38 (12): 1079–1082. doi:10.1130/G31182.1. http://geology.geoscienceworld.org/cgi/content/abstract/38/12/1079.
- ^ Tree of Life Web Project (2002). "Insecta". http://www.tolweb.org/Insecta/8205. Retrieved 2009-05-12.
- ^ David A. Kendall (2009). "Classification of Insect". http://www.kendall-bioresearch.co.uk/class.htm. Retrieved 2009-05-09.
- ^ Gilliott, Cedric (August 1995). Entomology (2 ed.). Springer-Verlag New York, LLC. pp. 820pp. ISBN 0-306-44967-6. http://books.google.com/?id=DrTKxvZq_IcC&pg=PA96&dq=Insect+classification+based+on+winged+and+wingless#v=onepage&q=Insect%20classification%20based%20on%20winged%20and%20wingless.
- ^ Kapoor, V.C. C. (January 1998). Principles and Practices of Animal Taxonomy. 1 (1 ed.). Science Publishers. p. 48. ISBN 1-57808-024-X.
- ^ Johnson, K. P., Yoshizawa, K. and V. S. Smith. 2004. Multiple origins of parasitism in lice. Proceedings of the Royal Society of London 271: 1771–1776.
- ^ Terry, M. D. and M. F. Whiting. 2005. Mantophasmatodea and phylogeny of the lower neopterous insects. Cladistics 21(3): 240–257
- ^ "Evidence from multiple gene sequences indicates that termites evolved from wood-feeding cockroaches.". E Current Biology 10 (13): 801–804. 2000. doi:10.1016/S0960-9822(00)00561-3.
- ^ Bonneton, F., F. G. Brunet, J. Kathirithamby, V. Laudet. 2006. The rapid divergence of the ecdysone receptor is a synapomorphy for Mecopterida that clarifies the Strepsiptera problem. Insect Molecular Biology 15(3):351–362.
- ^ Whiting, M.F. 2002. Mecoptera is paraphyletic: multiple genes and phylogeny of Mecoptera and Siphonaptera. Zoologica Scripta 31(1): 93–104.
- ^ Bale, JS; van Lenteren, F; Bigler, F. (27 February 2008). "Biological control and sustainable food production". Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 363 (1492): 761–776. doi:10.1098/rstb.2007.2182. PMC 2610108. PMID 1782711. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2610108.
- ^ Davidson, E. (2006). Big Fleas Have Little Fleas: How Discoveries of Invertebrate Diseases Are Advancing Modern Science. Tucson, Ariz.: University of Arizona Press. ISBN 0-8165-2544-7.
- ^ Colborn, T; vom Saal, FS; Soto, AM (October 1993). "Developmental effects of endocrine-disrupting chemicals in wildlife and humans". Environmental Health Perspectives 101 (5): 378–384. doi:10.2307/3431890. JSTOR 3431890. PMC 1519860. PMID 8080506. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1519860.
- ^ Nakamaru, M; Iwasab, Y; Nakanishic, J (October 2003). "Extinction risk to bird populations caused by DDT exposure". Chemosphere 53 (4): 377–387. doi:10.1016/S0045-6535(03)00010-9. PMID 12946395.
- ^ Smith, Deborah T (December 1991). Agriculture and the Environment: The 1991 Yearbook of Agriculture (1991 ed.). United States Government Printing. pp. 191 pp.. ISBN 0-16-034144-2. http://books.google.com/?id=fDTbAAAAMAAJ&q=insects+pollinate&dq=insects+pollinate.
- ^ "Biocontrol Network - Beneficial Insects". Biocontrol Network. http://www.biconet.com/biocontrol.html. Retrieved 2009-05-09.
- ^ Davidson, RH and William F. Lyon (1979). Insect Pests of Farm, Garden, and Orchard. Wiley, John & Sons, Incorporated. p. 38. ISBN 0-471-86314-9.
- ^ Dossey, Aaron T. (December 2010). "Insects and their chemical weaponry: New potential for drug discovery". Natural Product Reports (Royal Society of Chemistry (RSC Publishing)) 27: Pages 1737–1757. http://pubs.rsc.org/en/Content/ArticleLanding/2010/NP/C005319H.
- ^ Sherman, Ronald A.; Pechter, Edward A. (13 December 1987). "Maggot therapy: a review of the therapeutic applications of fly larvae in human medicine, especially for treating osteomyelitis". Medical and Veterinary Entomology (Journal compilation © 2009 The Royal Entomological Society) 2 (3): Pages 225–230. doi:10.1111/j.1365-2915.1988.tb00188.x. http://www3.interscience.wiley.com/journal/119457073/abstract.
- ^ Pierce, BA (2006). Genetics: A Conceptual Approach (2nd ed.). New York: W.H. Freeman and Company. p. 87. ISBN 0-7167-8881-0.
- ^ Adams, MD et al.; Celniker, SE; Holt, RA; Evans, CA; Gocayne, JD; Amanatides, PG; Scherer, SE; Li, PW et al. (24 March 2000). "The genome sequence of Drosophila melanogaster". Science 287 (5461): 2185–2195. Bibcode 2000Sci...287.2185.. doi:10.1126/science.287.5461.2185. PMID 10731132.
- ^ Michels, John (1880). John Michels. ed. Science. 1. American Association for the Advance of Science. 229 Broadway ave., N.Y.: American Association for the Advance of Science. pp. 2090pp. ISBN 1-930775-36-9. http://books.google.com/?id=aDkLAAAAYAAJ&pg=PA69&dq=Insects+also+produce+useful+substances+honey+silk#v=onepage&q=Insects%20also%20produce%20useful%20substances%20honey%20silk.
External links
- Insects of North America
- Overview of Orders of Insects
- Manual for Entomology and Arachnology
- Tree of Life Project – Insecta, Insecta Movies
- Insects of Brazil
- Insect Morphology Overview of insect external and internal anatomy
- Fossil Insect Database International Palaeoentological Society
- UF Book of Insect Records
- InsectImages.org 24,000 high resolution insect photographs
- BBC Nature: Insect news, and video clips from BBC programmes past and present.
- The Nature Explorers Many insect video clips.
Orders of Insects (Wikispecies) Extant Archaeognatha • Blattodea (Cockroaches) • Coleoptera (Beetles) • Dermaptera (Earwigs) • Diptera (Flies) • Embioptera (Webspinners) • Ephemeroptera (Mayflies) • Hemiptera • Hymenoptera • Isoptera (Termites) • Lepidoptera • Mantodea (Mantises) • Mecoptera • Megaloptera • Neuroptera • Notoptera • Odonata • Orthoptera • Phasmatodea • Phthiraptera (Lice) • Plecoptera • Psocoptera • Raphidioptera (Snakeflies) • Siphonaptera (Fleas) • Strepsiptera • Thysanoptera (Thrips) • Trichoptera (Caddisflies) • Zoraptera • Zygentoma (Thysanura)
Extinct Extant arthropod classes by subphylum Kingdom Animalia · Subkingdom Eumetazoa · (unranked) Bilateria · (unranked) Protostomia · Superphylum Ecdysozoa Chelicerata Arachnida (Araneae · Scorpiones · Opiliones · Acari · Pseudoscorpionida · Amblypygi · Thelyphonida · Solifugae · Palpigradi · Ricinulei · Schizomida) · Xiphosura · PycnogonidaMyriapoda Hexapoda Crustacea Branchiopoda · Remipedia · Cephalocarida · Maxillopoda (Cirripedia, Copepoda) · Ostracoda · Malacostraca (Decapoda, Amphipoda, Isopoda)Categories:- Insects
- Arthropods
- Entomology










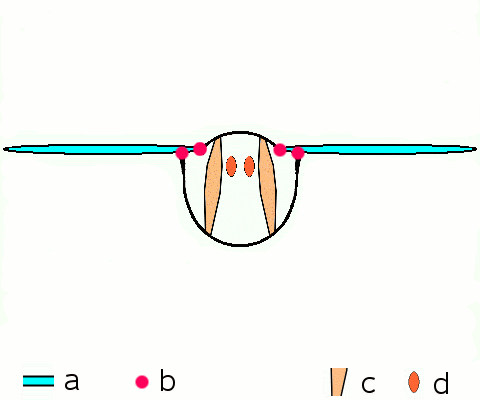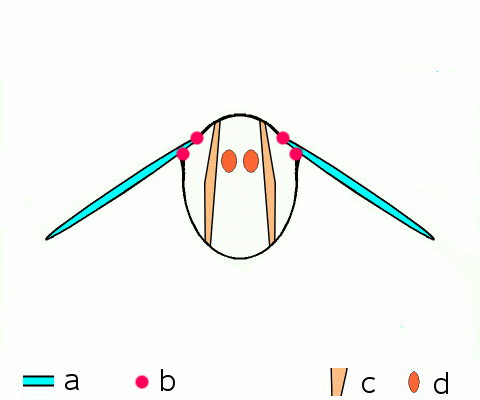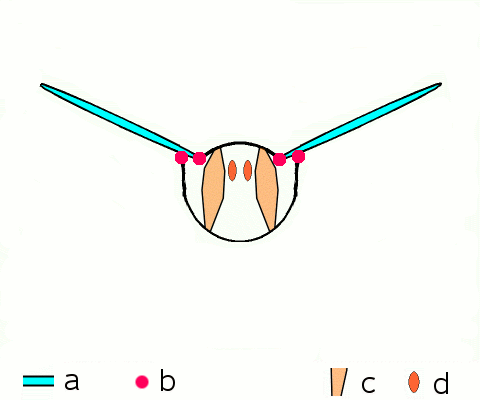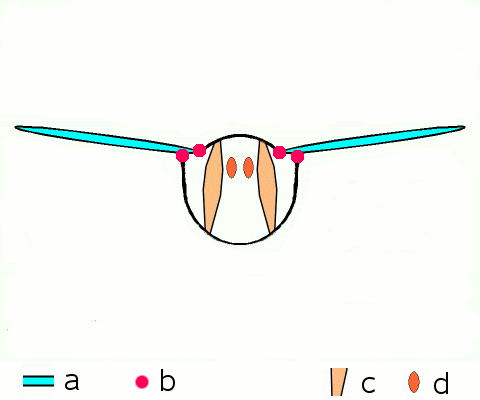






Wikimedia Foundation. 2010.