- Epistasis
-
See Epistaxis for the condition known colloquially as a nosebleed.
See Epitasis for the main action of a classical drama.
See Epitaxy for the method of depositing a monocrystalline film on a monocrystalline substrate.See also: Quantitative Trait LocusIn genetics, epistasis is the phenomenon where the effects of one gene are modified by one or several other genes, which are sometimes called modifier genes. The gene whose phenotype is expressed is called epistatic, while the phenotype altered or suppressed is called hypostatic. Epistasis can be contrasted with dominance, which is an interaction between alleles at the same gene locus. Epistasis is often studied in relation to Quantitative Trait Loci (QTL) and polygenic inheritance.
In general, the fitness increment of any one allele depends in a complicated way on many other alleles; but, because of the way that the science of population genetics was developed, evolutionary scientists tend to think of epistasis as the exception to the rule. In the first models of natural selection devised in the early 20th century, each gene was considered to make its own characteristic contribution to fitness, against an average background of other genes. Some introductory college courses still teach population genetics this way.
Epistasis and genetic interaction refer to different aspects of the same phenomenon. The term epistasis is widely used in population genetics and refers especially to the statistical properties of the phenomenon, and does not necessarily imply biochemical interaction between gene products. However, in general epistasis is used to denote the departure from 'independence' of the effects of different genetic loci. Confusion often arises due to the varied interpretation of 'independence' between different branches of biology. For further discussion of the definitions of epistasis, and the history of these definitions, see.[1]
Examples of tightly linked genes having epistatic effects on fitness are found in supergenes and the human major histocompatibility complex genes. The effect can occur directly at the genomic level, where one gene could code for a protein preventing transcription of the other gene. Alternatively, the effect can occur at the phenotypic level. For example, the gene causing albinism would hide the gene controlling color of a person's hair. In another example, a gene coding for a widow's peak would be hidden by a gene causing baldness. Fitness epistasis (where the affected trait is fitness) is one cause of linkage disequilibrium.
Studying genetic interactions can reveal gene function, the nature of the mutations, functional redundancy, and protein interactions. Because protein complexes are responsible for most biological functions, genetic interactions are a powerful tool.
Contents
Classification by fitness or trait value
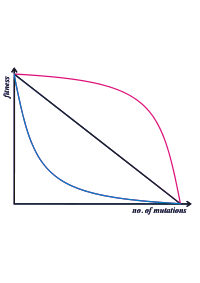 Diagram illustrating different relationships between numbers of mutations and fitness. Synergistic epistasis is the blue line - each mutation has a disproportionately large effect on the organism's fitness. Antagonistic epistasis is the red line. See Evolution of Sex
Diagram illustrating different relationships between numbers of mutations and fitness. Synergistic epistasis is the blue line - each mutation has a disproportionately large effect on the organism's fitness. Antagonistic epistasis is the red line. See Evolution of Sex
Two-locus epistatic interactions can be either synergistic (enhancing the effectiveness) or antagonistic (reducing the activity).[2][3] In the example of a haploid organism with genotypes (at two loci) AB, Ab, aB or ab, we can think of the following trait values where higher values suggest greater expression of the characteristic (the exact values are simply given as examples):
AB Ab aB ab No epistasis (additive across loci) 2 1 1 0 Synergistic epistasis 3 1 1 0 Antagonistic epistasis 1 1 1 0 Hence, we can classify thus:
Trait values Type of epistasis AB = Ab + aB − ab No epistasis, additive inheritance AB > Ab + aB − ab Synergistic epistasis AB < Ab + aB − ab Antagonistic epistasis Understanding whether the majority of genetic interactions are synergistic or antagonistic will help solve such problems as the evolution of sex.
Epistasis and sex
Negative epistasis and sex are thought to be intimately correlated. Experimentally, this idea has been tested in using digital simulations of asexual and sexual populations. Over time, sexual populations move towards more negative epistasis, or the lowering of fitness by two interacting alleles. It is thought that negative epistasis allows individuals carrying the interacting deleterious mutations to be removed from the populations efficiently. This removes those alleles from the population, resulting in an overall more fit population. This hypothesis was proposed by Alexey Kondrashov, and is sometimes known as the deterministic mutation hypothesis[4] and has also been tested using artificial gene networks.[2]
However, the evidence for this hypothesis has not always been straightforward and the model proposed by Kondrashov has been criticized for assuming mutation parameters far from real world observations. For example, see MacCarthy and Bergman.[5] In addition, in those tests which used artificial gene networks, negative epistasis is only found in more densely connected networks,[2] whereas empirical evidence indicates that natural gene networks are sparsely connected,[6] and theory shows that selection for robustness will favor more sparsely connected and minimally complex networks.[6]
Functional or mechanistic classification
- Genetic suppression - the double mutant has a less severe phenotype than either single mutant. [This term can also apply to a case where the double mutant has a phenotype intermediate between those of the single mutants, in which case the more severe single mutant phenotype is "suppressed" by the other mutation or genetic condition. For example, in a diploid organism, a hypomorphic (or partial loss-of-function) mutant phenotype can be suppressed by knocking out one copy of a gene that acts oppositely in the same pathway. In this case, the second gene is described as a "dominant suppressor" of the hypomorphic mutant; "dominant" because the effect is seen when one wild-type copy of the suppressor gene is present. For most genes, the phenotype of the heterozygous suppressor mutation by itself would be wild type (because most genes are not haplo-insufficient), so that the double mutant (suppressed) phenotype is intermediate between those of the single mutants.]
- Genetic enhancement - the double mutant has a more severe phenotype than one predicted by the additive effects of the single mutants.
- Synthetic lethality or unlinked non-complementation - two mutations fail to complement and yet do not map to the same locus.
- Intragenic complementation, allelic complementation, or interallelic complementation - two mutations map to the same locus, yet the two alleles complement in the heteroallelic diploid. Causes of intragenic complementation include:
- homology effects such as transvection, where, for example, an enhancer from one allele acts in trans to activate transcription from the promoter of the second allele.
- trans-splicing of two mutant RNA molecules to produce a functional RNA.
- At the protein level, another possibility involves proteins that normally function as dimers. In a heteroallelic diploid, two different abnormal proteins could form a functional dimer if each can compensate for the lack of function in the other.
See also
- Co-adaptation
- Epistasis and functional genomics
- Mutation
- Quantitative trait locus
References
- ^ Cordell, Heather J. (2002). "Epistasis: what it means, what it doesn't mean, and statistical methods to detect it in humans". Human Molecular Genetics 11 (20): 2463–8. doi:10.1093/hmg/11.20.2463. PMID 12351582.
- ^ a b c Azevedo R, Lohaus R, Srinivasan S, Dang K, Burch C (2006). "Sexual reproduction selects for robustness and negative epistasis in artificial gene networks". Nature 440 (7080): 87–90. doi:10.1038/nature04488. PMID 16511495.
- ^ Bonhoeffer S, Chappey C, Parkin NT, Whitcomb JM, Petropoulos CJ (2004). "Evidence for positive epistasis in HIV-1". Science 306 (5701): 1547–50. doi:10.1126/science.1101786. PMID 15567861.
- ^ A. S. Kondrashov (1988). "Deleterious mutations and the evolution of sexual reproduction". Nature 336 (6198): 435–440. doi:10.1038/336435a0. PMID 3057385.
- ^ MacCarthy T, Bergman A. (July 2007). "Coevolution of robustness, epistasis, and recombination favors asexual reproduction". Proc Natl Acad Sci U S A 104 (31): 12801–6. doi:10.1073/pnas.0705455104. PMC 1931480. PMID 17646644. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1931480.
- ^ a b Leclerc R. (August 2008). "Survival of the sparsest: robust gene networks are parsimonious". Mol Syst Biol. 4 (213): 213. doi:10.1038/msb.2008.52. PMC 2538912. PMID 18682703. http://www.nature.com/msb/journal/v4/n1/full/msb200852.html.
External links
- Science Aid: Epistasis High school (GCSE, Alevel) resource.
- GeneticInteractions.org
- Epistasis.org
The development of phenotype Key concepts Genetic architecture Dominance relationship · Epistasis · Polygenic inheritance · Pleiotropy · Plasticity · Canalisation · Fitness landscape · Transgressive phenotypeNon-genetic influences Developmental architecture Evolution of genetic systems Influential figures Debates List of evolutionary biology topics Categories:

Wikimedia Foundation. 2010.