- Dominance (genetics)
-
For other uses, see Dominance.
 A pedigree chart shows how genes are inherited.
A pedigree chart shows how genes are inherited.
Dominance in genetics is a relationship between two variant forms (alleles) of a single gene, in which one allele masks the effect of the other in influencing some trait. In the simplest case, if a gene exists in two allelic forms (A & a), three combinations of alleles (genotypes) are possible: AA, Aa, and aa. If Aa individuals (heterozygotes) show the same form of the trait (phenotype) as AA individuals (homozygotes), and aa homozygotes show a different phenotype, allele A is said to dominate or be dominant to allele a, and a is said to be recessive to A.
By one convention, dominant alleles are written in uppercase letters, and recessive alleles in lowercase letters.
Contents
Background
Diploid / haploid
Most familiar plants, like peas, and familiar animals, like fruit flies and humans, have paired chromosomes, and are described as diploid. One chromosome of each pair is contributed by each parent: one by the female parent in her ovum, and one by the male parent in his sperm, which are joined at fertilization. The ovum and sperm cells have only one copy of each chromosome and are described as haploid. Production of haploid gametes occurs through a process called meiosis.
Chromosomes, genes, and alleles
Each chromosome of a matching pair is structurally similar to the other, and each member of a homologous pair has the same genetic material arranged in the same order and physical locations (loci, singular locus). The genetic material in each chromosome comprises a series of discrete genes that influence various traits. Thus, each gene also has a corresponding homologue, which may exist in different forms: the variant forms are called alleles. The alleles at the same locus on the two homologous chromosomes may be identical or different.
In popular usage, "gene" and "allele" are often used interchangeably. This produces misunderstandings. Properly, "gene" refers to a hereditary unit, ordinarily at a fixed position on a chromosome, that influences a particular trait. Genes are now understood to comprise DNA. "Allele" refers to any of the many particular forms of a gene that may be present in a population of individuals from a particular species, at a particular locus. E.g., it is inaccurate to say "This pea plant has a pair of wrinkled genes", and it is more accurate to say, "This plant has two 'w' alleles for the 'Seed Shape' gene, and will produce wrinkled peas." Consider also the example of blood type in humans. Near the long arm of chromosome nine appears a gene that determines whether an individual will be blood type, A, B, or O. There are three different alleles that could be present at this locus, but only two can be present in any individual, one inherited from their mother and one from their father.[1]
Homozygous, heterozygous
If two alleles of a given gene are identical, the organism is called a homozygote and is homozygous with respect to that gene; if instead the two alleles are different, the organism is a heterozygote and is heterozygous. The genetic makeup of an organism, either at a single locus or over all its genes collectively, is called the genotype. The genotype of an organism directly or indirectly affects its molecular, physical,and other traits, which individually or collectively are called the phenotype. At heterozygous gene loci, the two alleles interact to produce the phenotype. The simplest form of allele interaction is the one described by Mendel, now called Mendelian, in which the appearance/phenotype caused by one allele is apparent, called dominant, and the appearance/phenotype caused by the other allele is not apparent, called recessive.
In the simplest case, the phenotypic effect of one allele completely masks the other in heterozygous combination; that is, the phenotype produced by the two alleles in heterozygous combination is identical to that produced by one of the two homozygous genotypes. The allele that masks the other is said to be dominant to the latter, and the alternative allele is said to be recessive to the former.[2]
Which trait is dominant?
The terms dominant and recessive refer to the interaction of alleles in producing the phenotype of the heterozygote. If there are two alternative phenotypes, by definition the phenotype exhibited by the heterozygote is called "dominant" and the "hidden" phenotype is called "recessive". The key concept of dominance is that the heterozygote is phenotypically identical to one of the two homozygotes. That trait corresponding to the dominant allele may then be called the "dominant" trait.
Dominance is a genotypic relationship between alleles, as manifested in the phenotype. It is unrelated to the nature of the phenotype itself, e.g., whether it is regarded as normal or abnormal, standard or nonstandard, healthy or diseased, stronger or weaker, or more or less extreme. It is also important to distinguish between the "round" gene locus, the "round" allele at that locus, and the "round" phenotype it produces. It is inaccurate to say that "the round gene dominates the wrinkled gene" or that "round peas dominate wrinkled peas."
Nomenclature
In genetics, the common convention is that dominant alleles are written as capital letters and recessive alleles as lower-case letters. In the pea example, once the dominance relationships of the two alleles are known, it is possible to designate the dominant allele that produces a round shape by a capital-letter symbol R, and the alternative recessive allele that produces a wrinkled shape by a lower-case symbol r. The homozygous dominant, heterozygous, and homozygous recessive genotypes are then written RR, Rr, and rr, respectively. It would also be possible to designate the two alleles as W and w, and the three genotypes WW, Ww, and ww, the first two of which produced round peas and the third wrinkled peas. Note that the choice or "R" or "W" as the symbol for the dominant allele does not pre-judge whether the allele causing the "round" or "wrinkled" phenotype when homozygous is the dominant one.
Another system of notation designates the gene involved in seed shape as the "Shp" gene, which exists in two allelic forms, ShpR and Shpw, the dominance relationships of the two being indicated by the case of the superscripts. This system is the standard system in Drosophila genetics.
Relationship to other genetic concepts
The concept of dominance is involved with a number of other genetic concepts.
Multiple alleles
Although any individual has at most two different alleles, most genes exist in a large number of allelic forms in the population as a whole. In some cases, the alleles have different effects on the phenotype, and their dominance interactions with each other can be described as a series. For example, the best known human blood groups, the ABO system,[3] comprises three sets of alleles at the I locus, IA, IB, and IO. The first two are dominant to the latter: that is, the AA and AO genotypes produce indistinguishable blood group phenotypes, called "Type A", as do BB and BO, which produce "Type B" blood. In another example, coat color in siamese cats[4] and related breeds is determined by a series of alleles at the albino gene locus (c) that produce different levels of pigment and hence different levels of color dilution. Four of these are c+, cb, cs, and ca (standard, Burmese, siamese, and albino, respectively), where the first allele is completely dominant to the last three, and the last is completely recessive to the first three.
Incomplete and semi-dominance
Complete dominance occurs when the phenotype of the heterozygote is completely indistinguishable from that of the dominant homozygote. This is frequently not the case. Incomplete dominance occurs when the phenotype of the heterozygous genotype is an intermediate of the phenotypes of the homozygous genotypes. For example, the snapdragon flower color is either homozygous for red or white. When the red homozygous flower is paired with the white homozygous flower, the result yields a pink snapdragon flower. The pink snapdragon is the result of incomplete dominance.
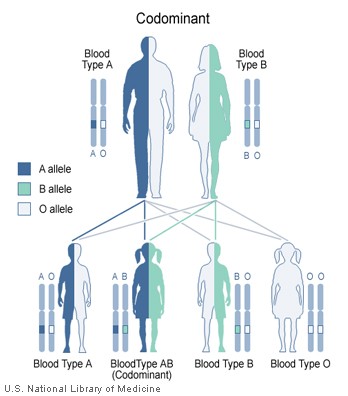
Co-dominance
 Blood types in humans show co-dominance.Main article: Codominance
Blood types in humans show co-dominance.Main article: CodominanceCo-dominance occurs when the contributions of both alleles are visible in the phenotype. In the ABO example, the IA and IB alleles are co-dominant in producing the AB blood group phenotype, in which both A-type and B-type antigens are made. Another example occurs at the locus for the Beta-globin component of hemoglobin, where the three molecular phenotypes of HbA/HbA, HbA/HbS, and HbS/HbS are all equally detectable by protein electrophoresis. (The medical condition produced by the heterozygous genotype is called an incomplete dominant, see above). For most gene loci at the molecular level, both alleles are expressed co-dominantly, because both are transcribed into RNA.
Co-dominance is different from incomplete or semi-dominance. For example, pink flowers might be the product of two alleles that produce red and white pigments that become mixed (co-dominance on the pigment level, no dominance on the color level), or the result of one allele that produces the usual amount of red pigment and another non-functional allele that produces no pigment, so as to produce a dilute, intermediate pink color (no dominance at either level).
Autosomal versus sex-linked dominance
In humans and other mammal species, sex is determined by two sex chromosomes called the X chromosome and the Y chromosome. Human females are typically XX; males are typically XY. The remaining pairs of chromosome are found in both sexes and are called autosomes; genetic traits due to loci on these chromosomes are described as autosomal, and may be dominant or recessive. Genetic traits on the X and Y chromosomes are called sex linked, because they tend to be characteristic of one sex or the other. In practice, the term almost always refers to X-linked traits. Females have two copies of every gene locus found on the X chromosome, just as for the autosomes, and the same dominance relationships apply. Males however have only one copy of each X chromosome gene locus, and are described as hemizygous for these genes. The Y chromosome is much smaller than the X, and contains a much smaller set of genes that influence 'maleness', such as the SRY gene for testis determining factor. Dominance rules for sex-linked gene loci are determined by their behavior in the female: because the male has only one allele, that allele is always expressed regardless of whether it is dominant or recessive. An example of sex-linked is: Hair loss is a trait caused by a gene located on the X chromosomes.[5]
Epistasis
Main article: EpistasisEpistasis ["epi + stasis = to sit on top"] is an interaction between genotypes at two different gene loci, which sometimes resembles a dominance interaction at a single locus. Epistasis modifies the characteristic 9:3:3:1 ratio expected for two non-epistatic genes. Most genetic systems involve complex epistatic interactions among multiple gene loci. For two loci, 14 classes of epistatic interactions are recognized. As an example of recessive epistasis, one gene locus may determine whether a flower pigment is yellow (AA or Aa) or green (aa), while another locus determines whether the pigment is produced (BB or Bb) or not (bb). In a bb plant, the flowers will be white, irrespective of the genotype of the other locus as AA, Aa, or aa. The b allele is not dominant to the A allele: the B locus shows recessive epistasis to the A locus, because the B locus when homozygous for the recessive allele (bb) suppresses phenotypic expression of the A locus. In a cross between two AaBb plants, this produces a characteristic 9:3:4 ratio, in this case of yellow : green : white flowers.
In dominant epistasis, one gene locus may determine yellow and green pigment as in the previous example: AA and Aa are yellow, and aa are green. A second locus determines whether a pigment precursor is produced (dd) or not (DD or Dd). Here, in a D- plant, the flowers will be colorless irrespective of the genotype at the A locus, because of the epistatic effect of the dominant D allele. Thus, in a cross between two AaDd plants, 3/4 of the plants will be colorless, and the yellow and green phenotypes are expressed only in dd plants. This produces a characteristic 12:3:1 ratio of white : yellow : green plants.
Supplementary epistasis occurs when two loci affect the same phenotype. For example, if pigment color is produced by CC or Cc but not cc, and by DD or Dd but not dd, then pigment is produced only in C-D- genotypes, and not in any genotype combination with cc or dd. That is, both loci must have at least one dominant allele to produce the phenotype. This produces a characteristic ratio 9:7 ratio of pigmented to unpigmented plants.[5]
Molecular mechanisms
The molecular basis of dominance was unknown to Mendel. It is now understood that a gene locus includes a long series (hundreds to thousands) of bases or nucleotides of deoxyribonucleic acid (DNA) at a particular point on a chromosome. The central dogma of molecular biology states that "DNA makes RNA makes protein", that is, that DNA is transcribed to make an RNA copy, and RNA is translated to make a protein. In this process, different alleles at a locus may or may not be transcribed, and if transcribed may be translated to slightly different forms of the same protein (called isoforms). Proteins often function as enzymes that catalyze chemical reactions in the cell, which directly or indirectly produce phenotypes. In any diploid organism, the DNA sequences of the two alleles present at any gene locus may be identical (homozygous) or different (heterozygous). Even if the gene locus is heterozygous at the level of the DNA sequence, the proteins made by each allele may be identical. In the absence of any difference between the protein products, neither allele can be said to be dominant (see co-dominance, above). Even if the two protein products are slightly different (allozymes), it is likely that they produce the same phenotype with respect to enzyme action, and again neither allele can be said to be dominant.
Dominance typically occurs when one of the two alleles is non-functional at the molecular level, that is, it is not transcribed or else does not produce a protein product. This can be the result of a mutation that alters the DNA sequence of the allele. An organism homozygous for the non-functional allele will generally show a distinctive phenotype, due to the absence of the protein product. For example, in humans and other organisms, the unpigmented skin of the albino phenotype [6] results when an individual is homozygous for an allele that prevents synthesis of the skin pigment protein melanin. It is important to understand that it is not the lack of function that allows the allele to be described as recessive: this is the interaction with the alternative allele in the heterozygote. Three general types of interaction are possible:
- In the typical case, the single functional allele makes sufficient protein to produce a phenotype identical to that of the homozygote: this is called haplosufficiency. For example, suppose the standard amount of enzyme produced in the functional homozygote is 100%, with the two functional alleles contributing 50% each. The single functional allele in the heterozygote produces 50% of the standard amount of enzyme, which is sufficient to produce the standard phenotype. If the heterozygote and the functional-allele homozygote have identical phenotypes, the functional allele is dominant to the non-functional allele. This occurs at the albino gene locus: the heterozygote produces sufficient enzyme to convert the pigment precursor to melanin, and the individual has standard pigmentation.
- Alternatively, a single functional allele in the heterozygote may produce insufficient gene product for proper function, and the phenotype resembles that of the homozygote for the non-functional allele. This haploinsufficiency is much less common: usually the deficiency of gene product results in incomplete dominance (below).
- The intermediate interaction occurs where the heterozygous genotype produces a phenotype intermediate between the two homozygotes. Depending on which of the two homozygotes the heterozygote most resembles, one allele is said to show incomplete dominance over the other. For example, in humans the Hb gene locus is responsible for the Beta-chain protein (HBB) that is one of the two globin proteins that make up the blood pigment hemoglobin[6] Many people are homozygous for an allele called HbA; some persons carry an alternative allele called HbS, either as homozygotes or heterozygotes. The hemoglobin molecules of HbS/HbS homozygotes undergo a change in shape that distorts the morphology of the red blood cells, and causes a severe, life-threatening form of anemia called sickle-cell anemia. Persons heterozygous HbA/HbS for this allele have a much less severe form of anemia called sickle-cell trait. Because the disease phenotype of HbA/HbS heterozygotes is more similar to but not identical to the HbA/HbA homozygote, the HbA allele is said to be incompletely dominant to the HbS allele.
In some cases, dominance of a non-standard allele results when that allele produces a defective protein that interferes with the proper function of the protein produced by the standard allele. The presence of the defective protein "dominates" the standard protein, and the disease phenotype of the heterozygote more closely resembles that of the homozygote for two variant alleles. This phenomenon occurs in a number of trinucleotide repeat diseases: for an example and more details see Huntington Disease.[7]
Dominant and recessive genetic diseases in humans
In humans, many genetic traits or diseases are classified simply as "dominant" or "recessive." Especially with respect to so-called recessive diseases, this can oversimplify the underlying molecular basis and lead to misunderstanding of the nature of dominance. For example, the genetic disease phenylketonuria (PKU)[8] results from any of a large number (>60) of alleles at the gene locus for the enzyme phenylalanine hydroxylase (PAH).[3] Many of these alleles produce little or no PAH, as a result of which the substrate phenylalanine and its metabolic byproducts accumulate in the central nervous system and can cause severe mental retardation if untreated.
The genotypes and phenotypic consequences of interactions among three alleles are shown in the following table:[9]
Genotype PAH activity [phe] conc PKU ? AA 100% 60 uM No AB 30% 120 uM No CC 5% 200 ~ 300 uM Hyperphenylalanemia BB 0.3% 600 ~ 2400 uM Yes In unaffected persons homozygous for a standard functional allele (AA), PAH activity is standard (100%), and the concentration of phenylalanine in the blood [phe] is about 60 uM. In untreated persons homozygous for one of the PKU alleles (BB), PAH activity is close to zero, [phe] ten to forty times standard, and the individual manifests PKU.
In the AB heterozygote, PAH activity is only 30% (not 50%) of standard, blood [phe] is elevated two-fold, and the person does not manifest PKU. Thus, the A allele is dominant to the B allele with respect to PKU, but the B allele is incompletely dominant to the A allele with respect to its molecular effect, determination of PAH activity level (0.3% < 30% << 100%). Finally, the A allele is an incomplete dominant to B with respect to [phe], as 60 uM < 120 uM << 600 uM. Note once more that it is irrelevant to the question of dominance that the recessive allele produces a more extreme [phe] phenotype.
For a third allele C, a CC homozygote produces a very small amount of PAH enzyme, which results in a somewhat elevated level of [phe] in the blood, a condition called hyperphenylalanemia, which does not result in mental retardation.
That is, the dominance relationships of any two alleles may vary according to which aspect of the phenotype is under consideration. It is typically more useful to talk about the phenotypic consequences of the allelic interactions involved in any genotype, rather than to try to force them into dominant and recessive categories.
History
The concept of dominance was first described by the "Father of Genetics," Gregor Mendel, in the 1860s. Mendel observed that, for a variety of traits of garden peas having to do with the appearance of seeds, seed pods, and plant appearance, there occurred two discrete phenotypes: round vs wrinkled, or yellow vs green seeds, red vs white flowers, tall vs short plants, and so on. When bred separately, the plants always produced the same phenotypes, generation after generation. However, when lines with different phenotypes were crossed (interbred), one and only one of the parental phenotypes showed up in the offspring: green, or round, or red, or tall, and so on. However, when these hybrid plants were crossed, the offspring plants showed the two original phenotypes, in a characteristic 3:1 ratio, with the more common type having the phenotype of the parental hybrid plants. Mendel reasoned that each of the parents in the first cross were homozygotes for different alleles (AA and aa), that each contributed one allele to the offspring, such that all of these hybrids were heterozygotes (Aa), and that one of the two alleles in the hybrid cross dominated expression of the other: A masked a. The final cross between two heterozygotes (Aa X Aa) would produce AA, Aa, and aa offspring in a 1:2:1 genotype ratio with the first three classes showing the "A" phenotype, and the last showing the "a" phenotype, thereby producing the 3:1 phenotype ratio.
Mendel did not use the terms gene, allele, phenotype, genotype, homozygote, and heterozygote, all of which were introduced afterward. He did introduce the notation of capital and lowercase letters for dominant and recessive alleles, respectively, still in use today.
See also
References
- ^ Ridley, Matt (1999). "Disease". Genome. Harper Collins. pp. 136–146. http://books.google.com/books?id=h2zcDWshkEkC&pg=PA136#v=onepage&f=false.
- ^ King et al., RC (2006). A Dictionary of Genetics (7th ed.). Oxford University Press. p. 129. ISBN 978-0195307610. http://books.google.com/books?id=ykp-7oJ5pREC&pg=PA129#v=onepage&f=false. "Dominance [refers] to alleles that fully manifest their phenotype when present in the heterozygous ... state."
- ^ a b "OMIM Entry - *612349 - PHENYLALANINE HYDROXYLASE; PAH". OMIM.org.
- ^ "Cat Coat Color". Vgl.ucdavis.edu. doi:10.1111/j.1365-2052.2009. http://www.vgl.ucdavis.edu/service/cat/coatcolor.html. Retrieved 2011-11-02.
- ^ a b Carr, Steven M. "Extensions to Mendelian Analysis". Mun.ca. Memorial University of Newfoundland. http://www.mun.ca/biology/scarr/2250_Extensions_to_Mendelian_Analysis.html. Retrieved 2011-11-02.
- ^ a b "Albinism, oculocutaneous, type IA, 203100". OMIM.org.
- ^ "Huntington disease 143100". OMIM.org.
- ^ "Hyperphenylalaninemia, non-PKU mild - 261600". OMIM.org.
- ^ Carr, Steven M.. "One Gene, One Enzyme". Mun.ca. Memorial University of Newfoundland. http://www.mun.ca/biology/scarr/2250_One_Gene_One_Enzyme.html. Retrieved 2011-11-02.
- "On-line notes for Biology 2250 – Principles of Genetics". Memorial University of Newfoundland.
- "Sickle-Cell Anemia". NIH.gov.
- "ABO blood groups". NIH.gov.
External links
- "Online Mendelian Inheritance in Man (OMIM)"
- "Autosomal dominance of Huntington's Disease". Huntington's Disease Outreach Project for Education at Stanford
Categories:

Wikimedia Foundation. 2010.