- ABO blood group system
-
"ABO" redirects here. For other uses, see ABO (disambiguation)."H substance" redirects here. For use in inflammation, see histamine.
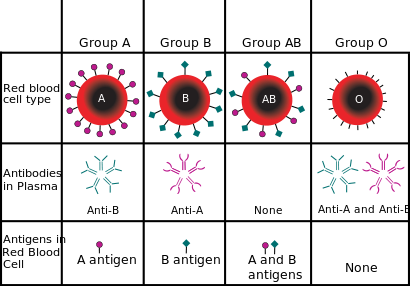
The ABO blood group system is the most important blood type system (or blood group system) in human blood transfusion. The associated anti-A antibodies and anti-B antibodies are usually IgM antibodies, which are usually produced in the first years of life by sensitization to environmental substances such as food, bacteria, and viruses. ABO blood types are also present in some other animals, for example apes such as chimpanzees, bonobos, and gorillas.[1]
History of discoveries
The ABO blood group system is widely credited to have been discovered by the Austrian scientist Karl Landsteiner, who found three different blood types in 1900;[2] he was awarded the Nobel Prize in Physiology or Medicine in 1930 for his work. Due to inadequate communication at the time it was subsequently found that Czech serologist Jan Janský had independently pioneered the classification of human blood into four groups,[3] but Landsteiner's independent discovery had been accepted by the scientific world while Janský remained in relative obscurity. Janský's classification is however still used in Russia and states of former USSR (see below). In America, Moss published his own (very similar) work in 1910.[4]
Landsteiner described A, B, and O; Alfred von Decastello and Adriano Sturli discovered the fourth type, AB, in 1902.[5] Ludwik Hirszfeld and E. von Dungern discovered the heritability of ABO blood groups in 1910–11, with Felix Bernstein demonstrating the correct blood group inheritance pattern of multiple alleles at one locus in 1924.[6] Watkins and Morgan, in England, discovered that the ABO epitopes were conferred by sugars, to be specific, N-acetylgalactosamine for the A-type and galactose for the B-type.[7][8][9] After much published literature claiming that the ABH substances were all attached to glycosphingolipids, Laine's group (1988) found that the band 3 protein expressed a long polylactosamine chain[10] that contains the major portion of the ABH substances attached.[11] Later, Yamamoto's group showed the precise glycosyl transferase set that confers the A, B and O epitopes.[12]
ABO antigens
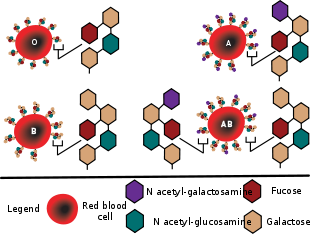 Diagram showing the carbohydrate chains that determine the ABO blood group
Diagram showing the carbohydrate chains that determine the ABO blood group
The H antigen is an essential precursor to the ABO blood group antigens. The H locus is located on chromosome 19. It contains 3 exons that span more than 5 kb of genomic DNA, and it encodes a fucosyltransferase that produces the H antigen on RBCs. The H antigen is a carbohydrate sequence with carbohydrates linked mainly to protein (with a minor fraction attached to ceramide moiety). It consists of a chain of β-D-galactose, β-D-N-acetylglucosamine, β-D-galactose, and 2-linked, α-L-fucose, the chain being attached to the protein or ceramide.
The ABO locus is located on chromosome 9. It contains 7 exons that span more than 18 kb of genomic DNA. Exon 7 is the largest and contains most of the coding sequence. The ABO locus has three main alleleic forms: A, B, and O. The A allele encodes a glycosyltransferase that bonds α-N-acetylgalactosamine to D-galactose end of H antigen, producing the A antigen. The B allele encodes a glycosyltransferase that joins α-D-galactose bonded to D-galactose end of H antigen, creating the B antigen.
In case of O allele, the exon 6 contains a deletion that results in a loss of enzymatic activity. The O allele differs from the A allele by deletion of only one nucleotide – guanine at position 261. The deletion causes a frameshift, and results in premature termination of translation, and thus, degradation of the mRNA. This results in H antigen remaining unchanged in case of O groups.
The majority of the ABO antigens are expressed on the ends of long polylactosamine chains attached mainly to band 3 protein, the anion exchange protein of the RBC membrane, and a minority of the epitopes are expressed on neutral [[glycosphingolipid].
Serology
Anti-A and anti-B antibodies (called isohaemagglutinins), which are not present in the newborn, appear in the first years of life. They are isoantibodies, that is, they are produced by an individual against antigens produced by members of the same species (isoantigens). Anti-A and anti-B antibodies are usually IgM type, which are not able to pass through the placenta to the fetal blood circulation. O-type individuals can produce IgG-type ABO antibodies.
Origin theories
It is possible that food and environmental antigens (bacterial, viral, or plant antigens) have epitopes similar enough to A and B glycoprotein antigens. The antibodies created against these environmental antigens in the first years of life can cross-react with ABO-incompatible red blood cells that it comes in contact with during blood transfusion later in life. Anti-A antibodies are hypothesized to originate from immune response towards influenza virus, whose epitopes are similar enough to the α-D-N-galactosamine on the A glycoprotein to be able to elicit a cross-reaction. Anti-B antibodies are hypothesized to originate from antibodies produced against Gram-negative bacteria, such as E. coli, cross-reacting with the α-D-galactose on the B glycoprotein.[13]
The "Light in the Dark theory" (DelNagro, 1998) suggests that, when budding viruses acquire host cell membranes from one human patient (in particular, from the lung and mucosal epithelium where they are highly expressed), they also take along ABO blood antigens from those membranes, and may carry them into secondary recipients where these antigens can elicit a host immune response against these non-self foreign blood antigens. These viral-carried human blood antigens may be responsible for priming newborns into producing neutralizing antibodies against foreign blood antigens. Support for this theory has come to light in recent experiments with HIV. HIV can be neutralized in in vitro experiments using antibodies against blood group antigens specifically expressed on the HIV-producing cell lines.[14][15]
The "Light in the Dark theory" suggests a novel evolutionary hypothesis: there is true communal immunity, which has developed to reduce the inter-transmissibility of viruses within a population. It suggests that individuals in a population supply and make a diversity of unique antigenic moieties so as to keep the population as a whole more resistant to infection. A system set up ideally to work with variable recessive alleles.[citation needed]
However, it is more likely that the force driving evolution of allele diversity is simply negative frequency-dependent selection; cells with rare variants of membrane antigens are more easily distinguished by the immune system from pathogens carrying antigens from other hosts. Thus, individuals possessing rare types are better equipped to detect pathogens. The high within-population diversity observed in human populations would, then, be a consequence of natural selection on individuals[16]
Nonantigen biology
The carbohydrate molecules on the surfaces of red blood cells have roles in cell membrane integrity, cell adhesion, membrane transportation of molecules, and acting as receptors for extracellular ligands, and enzymes. ABO antigens are found having similar roles on epithelial cells as well as red blood cells.[17][18]
Transfusion reactions
Due to the presence of isoantibodies against non-self blood group antigens, individuals of type A blood group immediately raise anti-B antibodies against B-blood group RBCs if transfused with blood from B group. The anti-B antibodies bind to B antigens on RBCs and cause complement-mediated lysis of the RBCs. The same happens for B and O groups (which raises both anti-A and anti-B antibodies). However, only blood group AB does not have anti-A and anti-B isoantibodies. This is because both A and B-antigens are present on the RBCs and are both self-antigens, hence they can receive blood from all groups and are universal recipient.
As far as transfusion compatibility is concerned, it is not strictly as simple as matching A, B, and O groups. In other words, no individual will ever receive a blood transfusion based on the ABO system alone. The rhesus factor must also be considered. Together, the rhesus factor and ABO grouping are the two most important compatibility factors to consider. An individual may be Rh+ or Rh-. In simpler terms, if an individual is blood type A and positive for the rhesus factor, then he or she is deemed "A+".
ABO and Rh blood type donation showing matches between donor and recipient types Recipients Donors O+ A+ B+ AB+ O- ** A- B- AB- O+ • • A+ • • • • B+ • • • • AB+ * • • • • • • • • O- • A- • • B- • • AB- • • • • - * Type AB+ is the universal recipient: Although those with AB blood type may be referred to as universal recipients, in actuality, type AB+ blood is that of the universal recipient, whereas type AB- is not. This is an important distinction to make.
- ** Because A-, A+, B-, B+, AB-, AB+, O- and O+ individuals can all receive blood from donors of type O- blood, an individual with type O- blood is deemed universal donor. In similar manner, O+ is not the universal donor blood type.
One caveat to this axiom of 'universal donor' is that this applies to packed RBCs, and not to whole blood products. Using the first table, type O carries anti-A and anti-B antibodies in the serum. To transfuse a type A, B, or AB recipient with type O whole blood would produce a hemolytic transfusion reaction due to the antibodies found in the serum of whole blood.
No antibodies are formed against the H antigen, except in those individuals with the Bombay phenotype.
In ABH secretors, ABH antigens are secreted by most mucus-producing cells of the body interfacing with the environment, including lung, skin, liver, pancreas, stomach, intestines, ovaries, and prostate.[19]
ABO hemolytic disease of the newborn
Main article: Hemolytic disease of the newborn (ABO)ABO blood group incompatibilities between the mother and child does not usually cause hemolytic disease of the newborn (HDN) because antibodies to the ABO blood groups are usually of the IgM type, which do not cross the placenta; however, in an O-type mother, IgG ABO antibodies are produced and the baby can develop ABO hemolytic disease of the newborn.
Inheritance
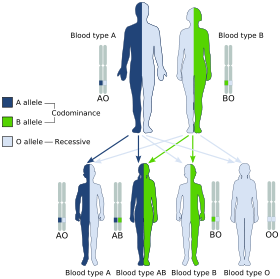 A and B are codominant, giving the AB phenotype.
A and B are codominant, giving the AB phenotype.Blood groups are inherited from both parents. The ABO blood type is controlled by a single gene (the ABO gene) with three alleles: i, IA, and IB. The gene encodes a glycosyltransferase—that is, an enzyme that modifies the carbohydrate content of the red blood cell antigens. The gene is located on the long arm of the ninth chromosome (9q34).
The IA allele gives type A, IB gives type B, and i gives type O. As both IA and IB are dominant over i, only ii people have type O blood. Individuals with IAIA or IAi have type A blood, and individuals with IBIB or IBi have type B. IAIB people have both phenotypes, because A and B express a special dominance relationship: codominance, which means that type A and B parents can have an AB child. A type A and a type B couple can also have a type O child if they are both heterozygous (IBi,IAi) The cis-AB phenotype has a single enzyme that creates both A and B antigens. The resulting red blood cells do not usually express A or B antigen at the same level that would be expected on common group A1 or B red blood cells, which can help solve the problem of an apparently genetically impossible blood group.[20]
Distribution and evolutionary history
The distribution of the blood groups A, B, O and AB varies across the world according to the population. There are also variations in blood type distribution within human subpopulations.
In the UK, the distribution of blood type frequencies through the population still shows some correlation to the distribution of placenames and to the successive invasions and migrations including Vikings, Danes, Saxons, Celts, and Normans who contributed the morphemes to the placenames and the genes to the population.[21]
There are six common alleles in white individuals of the ABO gene that produce one's blood type:[22][23]
- A
- A101 (A1)
- A201 (A2)
- B
- B101 (B1)
- O
- O01 (O1)
- O02 (O1v)
- O03 (O2)
Many rare variants of these alleles have been found in human populations around the world.
Some evolutionary biologists theorize that the IA allele evolved earliest, followed by O (by the deletion of a single nucleotide, shifting the reading frame) and then IB.[citation needed] This chronology accounts for the percentage of people worldwide with each blood type. It is consistent with the accepted patterns of early population movements and varying prevalent blood types in different parts of the world: for instance, B is very common in populations of Asian descent, but rare in ones of Western European descent. Another theory states that there are four main lineages of the ABO gene and that mutations creating type O have occurred at least three times in humans.[24] From oldest to youngest, these lineages comprise the following alleles: A101/A201/O09, B101, O02 and O01. The continued presence of the O alleles is hypothesized to be the result of balancing selection.[24] Both theories contradict the previously held theory that type O blood evolved earliest.
ABO and Rh distribution by country
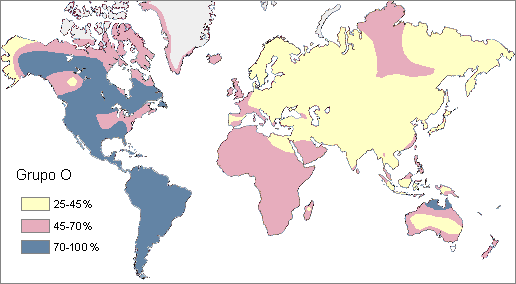
 Frequency of O group in indigenous populations around the world
Frequency of O group in indigenous populations around the worldABO and Rh blood type distribution by country (population averages) Country Population[25] O+ A+ B+ AB+ O- A- B- AB- Australia[26] 21,262,641 40.0% 31.0% 8.0% 2.0% 9.0% 7.0% 2.0% 1.0% Austria[27] 8,210,281 30.0% 33.0% 12.0% 6.0% 7.0% 8.0% 3.0% 1.0% Belgium[28] 10,414,336 38.0% 34.0% 8.5% 4.1% 7.0% 6.0% 1.5% 0.8% Brazil[29] 198,739,269 36.0% 34.0% 8.0% 2.5% 9.0% 8.0% 2.0% 0.5% Canada[30] 33,487,208 39.0% 36.0% 7.6% 2.5% 7.0% 6.0% 1.4% 0.5% Czech Republic[31] 10,532,770 27.0% 36.0% 15.0% 7.0% 5.0% 6.0% 3.0% 1.0% Denmark[32] 5,500,510 35.0% 37.0% 8.0% 4.0% 6.0% 7.0% 2.0% 1.0% Estonia[33] 1,299,371 30.0% 31.0% 20.0% 6.0% 4.5% 4.5% 3.0% 1.0% Finland[34] 5,250,275 27.0% 38.0% 15.0% 7.0% 4.0% 6.0% 2.0% 1.0% France[35] 62,150,775 36.0% 37.0% 9.0% 3.0% 6.0% 7.0% 1.0% 1.0% Germany[36] 82,329,758 35.0% 37.0% 9.0% 4.0% 6.0% 6.0% 2.0% 1.0% Hong Kong SAR[37] 7,055,071 40.0% 26.0% 27.0% 7.0% 0.3% 0.2% 0.1% 0.1% Iceland[38] 306,694 47.6% 26.4% 9.3% 1.6% 8.4% 4.6% 1.7% 0.4% India[39] 1,166,079,217 36.5% 22.1% 30.9% 6.4% 2.0% 0.8% 1.1% 0.2% Ireland[40] 4,203,200 47.0% 26.0% 9.0% 2.0% 8.0% 5.0% 2.0% 1.0% Israel[41] 7,233,701 32.0% 34.0% 17.0% 7.0% 3.0% 4.0% 2.0% 1.0% Netherlands[42] 16,715,999 39.5% 35.0% 6.7% 2.5% 7.5% 7.0% 1.3% 0.5% New Zealand[43] 4,213,418 38.0% 32.0% 9.0% 3.0% 9.0% 6.0% 2.0% 1.0% Norway[44] 4,660,539 34.0% 42.5% 6.8% 3.4% 6.0% 7.5% 1.2% 0.6% Poland[45] 38,482,919 31.0% 32.0% 15.0% 7.0% 6.0% 6.0% 2.0% 1.0% Portugal[46] 10,707,924 36.2% 39.8% 6.6% 2.9% 6.0% 6.6% 1.1% 0.5% Saudi Arabia[47] 28,686,633 48.0% 24.0% 17.0% 4.0% 4.0% 2.0% 1.0% 0.3% South Africa[48] 49,320,000 39.0% 32.0% 12.0% 3.0% 7.0% 5.0% 2.0% 1.0% Spain[49] 40,525,002 36.0% 34.0% 8.0% 2.5% 9.0% 8.0% 2.0% 0.5% Sweden[50] 9,059,651 32.0% 37.0% 10.0% 5.0% 6.0% 7.0% 2.0% 1.0% Turkey[51] 76,805,524 29.8% 37.8% 14.2% 7.2% 3.9% 4.7% 1.6% 0.8% United Kingdom[52] 61,113,205 37.0% 35.0% 8.0% 3.0% 7.0% 7.0% 2.0% 1.0% United States[53] 307,212,123 37.4% 35.7% 8.5% 3.4% 6.6% 6.3% 1.5% 0.6% Weighted mean 2,261,025,244 36.4% 28.3% 20.6% 5.1% 4.3% 3.5% 1.4% 0.5% Racial & Ethnic Distribution of ABO (without Rh) Blood Types[54]
(This table has more entries than the table above but does not distinguish between Rh types.)PEOPLE GROUP O (%) A (%) B (%) AB (%) Aborigines 61 39 0 0 Abyssinians 43 27 25 5 Ainu (Japan) 17 32 32 18 Albanians 38 43 13 6 Grand Andamanese 9 60 23 9 Arabs 34 31 29 6 Armenians 31 50 13 6 Asian (in USA - General) 40 28 27 5 Austrians 36 44 13 6 Bantus 46 30 19 5 Basques 51 44 4 1 Belgians 47 42 8 3 Blackfoot (N. Am. Indian) 17 82 0 1 Bororo (Brazil) 100 0 0 0 Brazilians 47 41 9 3 Bulgarians 32 44 15 8 Burmese 36 24 33 7 Buryats (Siberia) 33 21 38 8 Bushmen 56 34 9 2 Chinese-Canton 46 23 25 6 Chinese-Peking 29 27 32 13 Chuvash 30 29 33 7 Czechs 30 44 18 9 Danes 41 44 11 4 Dutch 45 43 9 3 Egyptians 33 36 24 8 English 47 42 9 3 Eskimos (Alaska) 38 44 13 5 Eskimos (Greenland) 54 36 23 8 Estonians 34 36 23 8 Fijians 44 34 17 6 Finns 34 41 18 7 French 43 47 7 3 Georgians 46 37 12 4 Germans 41 43 11 5 Greeks 40 42 14 5 Gypsies (Hungary) 29 27 35 10 Hawaiians 37 61 2 1 Hindus (Bombay) 32 29 28 11 Hungarians 36 43 16 5 Icelanders 56 32 10 3 Indians (India - General) 37 22 33 7 Indians (USA - General) 79 16 4 1 Irish 52 35 10 3 Italians (Milan) 46 41 11 3 Japanese 30 38 22 10 Jews (Germany) 42 41 12 5 Jews (Poland) 33 41 18 8 Kalmuks 26 23 41 11 Kikuyu (Kenya) 60 19 20 1 Koreans 28 32 31 10 Lapps 29 63 4 4 Latvians 32 37 24 7 Lithuanians 40 34 20 6 Malaysians 62 18 20 0 Maori 46 54 1 0 Mayas 98 1 1 1 Moros 64 16 20 0 Navajo (N. Am. Indian) 73 27 0 0 Nicobarese (Nicobars) 74 9 15 1 Norwegians 39 50 8 4 Papuas (New Guinea) 41 27 23 9 Persians 38 33 22 7 Peru (Indians) 100 0 0 0 Filipinos 45 22 27 6 Poles 33 39 20 9 Portuguese 35 53 8 4 Romanians 34 41 19 6 Russians 33 36 23 8 Sardinians 50 26 19 5 Scots 51 34 12 3 Serbians 38 42 16 5 Shompen (Nicobars) 100 0 0 0 Slovaks 42 37 16 5 South Africans 45 40 11 4 Spanish 38 47 10 5 Sudanese 62 16 21 0 Swedes 38 47 10 5 Swiss 40 50 7 3 Tartars 28 30 29 13 Thais 37 22 33 8 Turks 43 34 18 6 Ukrainians 37 40 18 6 USA (US blacks) 49 27 20 4 USA (US whites) 45 40 11 4 Vietnamese 42 22 30 5 Mean 43.91 34.80 16.55 5.14 Standard deviation 16.87 13.80 9.97 3.41 Blood group B has its highest frequency in Northern India and neighboring Central Asia, and its incidence diminishes both towards the west and the east, falling to single digit percentages in Spain.[55][56] It is believed to have been entirely absent from Native American and Australian Aboriginal populations prior to the arrival of Europeans in those areas.[56][57]
Blood group A is associated with high frequencies in Europe, especially in Scandinavia and Central Europe, although its highest frequencies occur in some Australian Aborigine populations and the Blackfoot Indians of Montana.[58][59]
Association with von Willebrand factor
The ABO antigen is also expressed on the von Willebrand factor (vWF) glycoprotein,[60] which participates in hemostasis (control of bleeding). In fact, having type O blood predisposes to bleeding,[61] as 30% of the total genetic variation observed in plasma vWF is explained by the effect of the ABO blood group,[62] and individuals with group O blood normally have significantly lower plasma levels of vWF (and Factor VIII) than do non-O individuals.[63][64] In addition, vWF is degraded more rapidly due to the higher prevalence of blood group O with the Cys1584 variant of vWF (an amino acid polymorphism in VWF):[65] the gene for ADAMTS13 (vWF-cleaving protease) maps to the ninth chromosome (9q34), the same locus as ABO blood type. Higher levels of vWF are more common amongst people who have had ischaemic stroke (from blood clotting) for the first time.[66] The results of this study found that the occurrence was not affected by ADAMTS13 polymorphism, and the only significant genetic factor was the person's blood group.
Disease association
Compared to non-O group (A, AB, and B) individuals, O group individuals have a 14% reduced risk of squamous cell carcinoma and 4% reduced risk of basal cell carcinoma.[67] It is also associated with a reduced risk of pancreatic cancer.[68][69] The B antigen links with increased risk of ovarian cancer.[70] Gastric cancer has reported to be more common in blood group A and least in group O.[71]
Subgroups
A1 and A2
The A blood type contains about twenty subgroups, of which A1 and A2 are the most common (over 99%). A1 makes up about 80% of all A-type blood, with A2 making up the rest.[72] These two subgroups are interchangeable as far as transfusion is concerned, but complications can sometimes arise in rare cases when typing the blood.[72]
Bombay phenotype
Main article: Hh antigen systemIndividuals with the rare Bombay phenotype (hh) do not express antigen H on their red blood cells. As H antigen serves as precursor for producing A and B antigens, the absence of H antigen means the individuals do not have A or B antigens as well (similar to O blood group). However, unlike O group, the H antigen is absent, hence the individuals produce isoantibodies to antigen H as well as to both A and B antigens. In case they receive blood from O blood group, the anti-H antibodies will bind to H antigen on RBC of donor blood and destroy the RBCs by complement-mediated lysis. Therefore Bombay phenotype can receive blood only from other hh donors (although they can donate as though they were type O).
Nomenclature in Europe and former USSR
 Ukraine uniform imprint B+
Ukraine uniform imprint B+In parts of Europe, the "O" in ABO blood type is substituted with "0" (zero), signifying the lack of A or B antigen. In the former USSR blood types are referenced using numbers and Roman numerals instead of letters. This is Janský's original classification of blood types. It designates the blood types of humans as I, II, III, and IV, which are elsewhere designated, respectively, as O, A, B, and AB.[73] The designation A and B with reference to blood groups was proposed by Ludwik Hirszfeld.
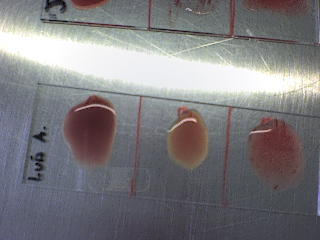
Examples of ABO and Rhesus D slide testing method
-
Blood group O positive: neither anti-A nor anti-B have agglutinated, but anti-Rh has
-

Result: Blood group A positive: anti-A and anti-Rh have agglutinated but anti-B has not.
In the slide testing method shown above, three drops of blood are placed on a glass slide with liquid reagents. Agglutination indicates the presence of blood group antigens in the blood.
Universal blood created from other types, and artificial blood
In April 2007, an international team of researchers announced in the journal Nature Biotechnology an inexpensive and efficient way to convert types A, B, and AB blood into type O.[74] This is done by using glycosidase enzymes from specific bacteria to strip the blood group antigens from red blood cells. The removal of A and B antigens still does not address the problem of the Rhesus blood group antigen on the blood cells of Rhesus positive individuals, and so blood from Rhesus negative donors must be used. Patient trials will be conducted before the method can be relied on in live situations.
Another approach to the blood antigen problem is the manufacture of artificial blood, which could act as a substitute in emergencies. BBC.
Conjectures
There are numerous popular conjectures surrounding ABO blood groups. These beliefs have existed since the ABO blood groups were identified and can be found in different cultures throughout the world. For example, during the 1930s, connecting blood groups to personality types became popular in Japan and other areas of the world.[75]
Additional myths include the idea that group A causes severe hangovers, group O is associated with perfect teeth, and those with blood group A2 have the highest IQs. Scientific evidence in support of these concepts is nonexistent.[76]
See also
References
- ^ Maton, Anthea; Jean Hopkins, Charles William McLaughlin, Susan Johnson, Maryanna Quon Warner, David LaHart, Jill D. Wright (1993). Human Biology and Health. Englewood Cliffs, New Jersey, USA: Prentice Hall. ISBN 0-13-981176-1.
- ^ Landsteiner K (1900). "Zur Kenntnis der antifermentativen, lytischen und agglutinierenden Wirkungen des Blutserums und der Lymphe". Zentralblatt Bakteriologie 27: 357–62.
- ^ Janský J (1907). "(Haematologick studie u. psychotiku" (in Czech). Sborn. Klinick 8: 85–139.
- ^ Moss WL (1910). "Studies on isoagglutinins and isohemolysins". Bulletin Johns Hopkins Hospital 21: 63–70.
- ^ von Decastello A, Sturli A (1902). "Ueber die Isoagglutinine im Serum gesunder und kranker Menschen". Mfinch med Wschr 49: 1090–5.
- ^ Crow J (1993). "Felix Bernstein and the first human marker locus". Genetics 133 (1): 4–7. PMC 1205297. PMID 8417988. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1205297. Full text at PMC: 1205297
- ^ Morgan, W. T. J. & Watkins, W. M. Br. Med. Bull. 25, 30–34 (1969)
- ^ Watkins, W. M. Advances in Human Genetics Vol. 10 (eds Harris, H. & Hirschhorn, K.) 1–136 (Plenum, New York, 1980)
- ^ Watkins, W. M. & Morgan, W. T. J. Vox Sang. 4, 97−119 (1959).
- ^ Jarnefelt, Rush, Li, Laine, J. Biol. Chem. 253: 8006–8009(1978)
- ^ Laine and Rush in Molecular Immunology of Complex Carbohydrates (A. Wu, E. Kabat, Eds.) Plenum Publishing Corporation, N.Y. NY (1988)
- ^ Yamamoto, et al., Molecular genetic basis of the histo-blood group ABO system, Nature 345: 229–233 (1990)
- ^ Letter to the Editor: “Natural” Versus Regular Antibodies Journal The Protein Journal Publisher Springer Netherlands ISSN 1572-3887 (Print) 1573-4943 (Online) Issue Volume 23, Number 6 / August, 2004 Category Letter to the Editor DOI 10.1023/B:JOPC.0000039625.56296.6e Page 357 Subject Group Chemistry and Materials Science Online Date Friday, January 07, 2005
- ^ Arendrup, M; Hansen JE, Clausen H, Nielsen C, Mathiesen LR, Nielsen JO (April 1991). "Antibody to histo-blood group A antigen neutralizes HIV produced by lymphocytes from blood group A donors but not from blood group B or O donors". AIDS 5 (4): 441–4. doi:10.1097/00002030-199104000-00014. PMID 1711864.
- ^ Neil, SJ; McKnight A, Gustafsson K, Weiss RA (2005-06-15). "HIV-1 incorporates ABO histo-blood group antigens that sensitize virions to complement-mediated inactivation". Blood 105 (12): 4693–9. doi:10.1182/blood-2004-11-4267. PMID 15728127. http://bloodjournal.hematologylibrary.org/cgi/content/full/105/12/4693.
- ^ Seymour RM, Allan MJ, Pomiankowski A, and Gustafsson K (2004) Evolution of the Human ABO Polymorphism by Two Complementary Selective Pressures. Proceedings: Biological Sciences 271:1065-1072.
- ^ Reid, ME; Mohandas, N (2004). "Red blood cell blood group antigens: structure and function.". Seminars in hematology 41 (2): 93–117. doi:10.1053/j.seminhematol.2004.01.001. PMID 15071789.
- ^ Mohandas, N; Narla, A (2005). "Blood group antigens in health and disease.". Current opinion in hematology 12 (2): 135–40. doi:10.1097/01.moh.0000153000.09585.79. PMID 15725904.
- ^ Laine, R.A. and Rush, J.S. (1988) "Chemistry of Human Erythrocyte Polylactosamine Glycopeptides (Erythroglycans) as Related to ABH Blood Group antigenic Determinants: Evidence that Band 3 Carbohydrate on Human Erythrocytes Carries the Majority of ABH Blood Group Substance" in Molecular Immunology of Complex Carbohydrates (A. Wu, E. Kabat, Eds.) Plenum Publishing Corporation, N.Y. NY.
- ^ Yazer M, Olsson M, Palcic M (2006). "The cis-AB blood group phenotype: fundamental lessons in glycobiology". Transfus Med Rev 20 (3): 207–17. doi:10.1016/j.tmrv.2006.03.002. PMID 16787828.
- ^ Potts, WTW (1979). "History and Blood Groups in the British Isles". In Sawyer PH. English Medieval Settlement. St. Martin's Press. ISBN 0-7131-6257-0.
- ^ Seltsam A, Hallensleben M, Kollmann A, Blasczyk R (2003). "The nature of diversity and diversification at the ABO locus". Blood 102 (8): 3035–42. doi:10.1182/blood-2003-03-0955. PMID 12829588.
- ^ Ogasawara K; Bannai M; Saitou N et al. (1996). "Extensive polymorphism of ABO blood group gene: three major lineages of the alleles for the common ABO phenotypes". Human Genetics 97 (6): 777–83. doi:10.1007/BF02346189. PMID 8641696.
- ^ a b Calafell, Francesc; et al. (September 2008). "Evolutionary dynamics of the human ABO gene". Human Genetics 124 (2): 123–135. doi:10.1007/s00439-008-0530-8. PMID 18629539. http://www.springerlink.com/content/yv4072vu67mv1166/fulltext.html. Retrieved 2008-09-24.
- ^ CIA World Factbook
- ^ Blood Types - What Are They?, Australian Red Cross[dead link]
- ^ "Austrian Red Cross - Blood Donor Information". Old.roteskreuz.at. 2006-03-21. http://old.roteskreuz.at/47_body.html#blutgruppen. Retrieved 2010-11-19.
- ^ "Rode Kruis Wielsbeke - Blood Donor information material". Rodekruiswielsbeke.be. http://www.rodekruiswielsbeke.be/infobloed.html. Retrieved 2010-11-19.
- ^ Tipos Sanguíneos
- ^ Canadian Blood Services - Société canadienne du sang. "Types & Rh System, Canadian Blood Services". http://www.bloodservices.ca/CentreApps/Internet/UW_V502_MainEngine.nsf/page/Blood%20Types%20and%20Rh%20System?OpenDocument. Retrieved 2010-11-19.
- ^ Czech Red Cross. "Podíl krevních skupin v populaci České republiky". http://www.cckpraha1.cz/krev.htm. Retrieved 2011-03-18.
- ^ Frequency of major blood groups in the Danish population.[dead link]
- ^ "Veregruppide esinemissagedus Eestis" (in Estonian). http://www.kliinikum.ee/verekeskus/index.php?menu=9&mod=page&id=5. Retrieved 2010-11-19.
- ^ "Suomalaisten veriryhmäjakauma" (in Finnish). 2009-08-21. http://www.veripalvelu.fi/asp/system/empty.asp?P=1275&VID=default&SID=908029945449597&S=1&C=24395. Retrieved 2010-11-19.
- ^ "Les groupes sanguins (système ABO)" (in French). Centre Hospitalier Princesse GRACE - Monaco. C.H.P.G. MONACO. 2005. http://www.chpg.mc/go/article.php3?id_article=111. Retrieved 2008-07-15.
- ^ de:Blutgruppe#Häufigkeit der Blutgruppen
- ^ Blood Donation, Hong Kong Red Cross[dead link]
- ^ "Blóðflokkar" (in Icelandic). .landspitali.is. http://www4.landspitali.is/lsh_ytri.nsf/pages/bb_blodflokkarnir. Retrieved 2010-11-19.
- ^ "Indian Journal for the Practising Doctor". Indmedica.com. http://www.indmedica.com/journals.php?journalid=3&issueid=75&articleid=965&action=article. Retrieved 2010-11-19.
- ^ "Irish Blood Transfusion Service - Irish Blood Group Type Frequency Distribution". Irish Blood Transfusion Service. http://www.ibts.ie/All_About_Blood/Blood_Group_Basics/. Retrieved 2009-11-07.
- ^ "The national rescue service in Israel" (in Hebrew). Mdais.org. http://www.mdais.org/362/. Retrieved 2010-11-19.
- ^ "Voorraad Erytrocytenconcentraten Bij Sanquin" (in Dutch). http://www.sanquin.nl/Sanquin-nl/erygrafiek.nsf/All/Voorraad-Erytrocytenconcentraten-Bij-Sanquin.html. Retrieved 2009-03-27.
- ^ "What are Blood Groups?". NZ Blood. http://www.nzblood.co.nz/?t=31. Retrieved 2010-11-19.
- ^ Norwegian Blood Donor Organization
- ^ "Regionalne Centrum Krwiodawstwa i Krwiolecznictwa we Wroclawiu" (in Polish). Rckik.wroclaw.pl. 2010-09-02. http://rckik.wroclaw.pl/?id=5&go=0. Retrieved 2010-11-19.
- ^ "Portuguese Blood Institute" (in Portuguese). http://www.ipsangue.org/maxcontent-documento-107.html. (assuming Rh and AB antigens are independent)
- ^ "Frequency of ABO blood groups in the eastern region of Saudi Arabia". Cat.inist.fr. http://cat.inist.fr/?aModele=afficheN&cpsidt=14126617. Retrieved 2010-11-19.
- ^ "South African National Blood Service - What's Your Type?". Sanbs.org.za. http://www.sanbs.org.za/donors_whattype.htm. Retrieved 2010-11-19.
- ^ "Federación Nacional de Donantes de Sangre/La sangre/Grupos". Donantesdesangre.net. http://www.donantesdesangre.net/menu.htm. Retrieved 2010-11-19.
- ^ "Frequency of major blood groups in the Swedish population". Geblod.nu. 2007-10-02. http://www.geblod.nu/general.aspx?PageId=10. Retrieved 2010-11-19.
- ^ "Turkey Blood Group Site". Kangrubu.com. http://www.kangrubu.com/default.asp?sayfa=kan_gruplari. Retrieved 2010-11-19.
- ^ "Frequency of major blood groups in the UK". Blood.co.uk. http://www.blood.co.uk/pages/all_about.html. Retrieved 2010-11-19.
- ^ "Blood Types in the U.S". Bloodcenter.stanford.edu. 2008-06-20. http://bloodcenter.stanford.edu/about_blood/blood_types.html. Retrieved 2010-11-19.
- ^ RACIAL & ETHNIC DISTRIBUTION of ABO BLOOD TYPES, BLOODBOOK.COM
- ^ Blood Transfusion Division, United States Army Medical Research Laboratory (1971). Selected contributions to the literature of blood groups and immunology. 1971 v. 4. United States Army Medical Research Laboratory, Fort Knox, Kentucky. http://books.google.com/?id=ALilcA7Acd0C. "... In northern India, in Southern and Central China and in the neighboring Central Asiatic areas, we find the highest known frequencies of B. If we leave this center, the frequency of the B gene decreases almost everywhere ..."
- ^ a b Encyclopaedia Britannica (2002). The New Encyclopaedia Britannica. Encyclopaedia Britannica, Inc.. ISBN 0-85229-787-4. http://books.google.com/?id=fpdUAAAAMAAJ. "... The maximum frequency of the B gene occurs in Central Asia and northern India. The B gene was probably absent from American Indians and Australian Aborigines before racial admixture occurred with the coming of the white man ..."
- ^ Carol R. Ember, Melvin Ember (1973). Anthropology. Appleton-Century-Crofts. http://books.google.com/?id=fvpFAAAAMAAJ. "... Blood type B is completely absent in most North and South American Indians ..."
- ^ Laura Dean, MD (2005). Blood Groups an Red Cell Antigens. National Center for Biotechnology Information, United States Government. ISBN 1-932811-05-2. http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=rbcantigen. "... Type A is common in Central and Eastern Europe. In countries such as Austria, Denmark, Norway, and Switzerland, about 45-50% of the population have this blood type, whereas about 40% of Poles and Ukrainians do so. The highest frequencies are found in small, unrelated populations. For example, about 80% of the Blackfoot Indians of Montana have blood type A ..."
- ^ (PDF) Technical Monograph No. 2: The ABO Blood Group System and ABO Subgroups. Biotec. March 2005. http://www.biotec.com/pdf/Technical%20Monograph%20No.%202%20-%20ABO%20system%20and%20subgroups.pdf. "... The frequency of blood group A is quite high (25-55%) in Europe, especially in Scandinavia and parts of central Europe. High group A frequency is also found in the Aborigines of South Australia (up to 45%) and in certain American Indian tribes where the frequency reaches 35% ..."
- ^ Sarode, R; Goldstein J, Sussman II, Nagel RL, Tsai HM (June 2000). "Role of A and B blood group antigens in the expression of adhesive activity of von Willebrand factor". Br J Haematol. 109 (4): 857–64. doi:10.1046/j.1365-2141.2000.02113.x. PMID 10929042.
- ^ O'Donnell, J; Laffan MA (August 2001). "The relationship between ABO histo-blood group, factor VIII and von Willebrand factor". Transfus Med. 11 (4): 343–51. doi:10.1046/j.1365-3148.2001.00315.x. PMID 11532189.
- ^ O'Donnell, J; Boulton FE, Manning RA, Laffan MA (2002-02-01). "Amount of H antigen expressed on circulating von Willebrand factor is modifiedby ABO blood group genotype and is a major determinant of plasma von Willebrand factor antigen levels". Arterioscler Thromb Vasc Biol. (American Heart Association, Inc.) 22 (2): 335–41. doi:10.1161/hq0202.103997. PMID 11834538. http://atvb.ahajournals.org/cgi/content/full/22/2/335.
- ^ Gill, JC; Endres-Brooks J, Bauer PJ, Marks WJ, Montgomery RR (June 1987). "The effect of ABO blood group on the diagnosis of von Willebrand disease" (abstract). Blood 69 (6): 1691–5. PMID 3495304. http://www.bloodjournal.org/cgi/content/abstract/69/6/1691.
- ^ Shima, M; Fujimura Y (1995). "ABO blood group genotype and plasma von Willebrand factor in normal individuals". Vox Sang 68 (4): 236–40. doi:10.1111/j.1423-0410.1995.tb02579.x. PMID 7660643.
- ^ Bowen, DJ; Collins PW (March 2005). "The prevalence of the cysteine1584 variant of von Willebrand factor is increased in type 1 von Willebrand disease: co-segregation with increased susceptibility to ADAMTS13 proteolysis but not clinical phenotype". Br J Haematol (Blackwell Synergy) 128 (6): 830–6. doi:10.1111/j.1365-2141.2005.05375.x. PMID 15755288.
- ^ Bongers T; de Maat M; van Goor M et al. (2006). "High von Willebrand factor levels increase the risk of first ischemic stroke: influence of ADAMTS13, inflammation, and genetic variability". Stroke 37 (11): 2672–7. doi:10.1161/01.STR.0000244767.39962.f7. PMID 16990571.
- ^ Xie J, Qureshi AA, Li Y, Han J, (2010). ABO Blood Group and Incidence of Skin Cancer. PLoS ONE 5(8): e11972. doi|10.1371/journal.pone.0011972
- ^ Wolpin, BM; Kraft, P; Gross, M; Helzlsouer, K; Bueno-De-Mesquita, HB; Steplowski, E; Stolzenberg-Solomon, RZ; Arslan, AA et al. (2010). "Pancreatic cancer risk and ABO blood group alleles: results from the pancreatic cancer cohort consortium". Cancer research 70 (3): 1015–23. doi:10.1158/0008-5472.CAN-09-2993. PMC 2943735. PMID 20103627. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2943735.
- ^ Amundadottir, L; Kraft, P; Stolzenberg-Solomon, RZ; Fuchs, CS; Petersen, GM; Arslan, AA; Bueno-De-Mesquita, HB; Gross, M et al. (2009). "Genome-wide association study identifies variants in the ABO locus associated with susceptibility to pancreatic cancer". Nature genetics 41 (9): 986–90. doi:10.1038/ng.429. PMC 2839871. PMID 19648918. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2839871.
- ^ Gates, MA; Wolpin, BM; Cramer, DW; Hankinson, SE; Tworoger, SS (2010). "ABO blood group and incidence of epithelial ovarian cancer". International journal of cancer. Journal international du cancer 128 (2): 482–486. doi:10.1002/ijc.25339. PMC 2946962. PMID 20309936. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2946962.
- ^ Aird, I; Bentall, HH; Roberts, JA (1953). "A relationship between cancer of stomach and the ABO blood groups". British medical journal 1 (4814): 799–801. doi:10.1136/bmj.1.4814.799. PMC 2015995. PMID 13032504. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2015995.
- ^ a b Blood Group A Suptypes, The Owen Foundation. Retrieved 2008-07-01.
- ^ Erb IH (1 May 1940). "Blood Group Classifications, a Plea for Uniformity". Canadian Medical Association Journal 42 (5): 418–21. PMC 537907. PMID 20321693. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=537907.
- ^ Liu, QP; Sulzenbacher G, Yuan H, Bennett EP, Pietz G, Saunders K, Spence J, Nudelman E, Levery SB, White T, Neveu JM, Lane WS, Bourne Y, Olsson ML, Henrissat B, Clausen H (April 2007). "Bacterial glycosidases for the production of universal red blood cells". Nat Biotechnol 25 (4): 454–64. doi:10.1038/nbt1298. PMID 17401360. http://www.nature.com/nbt/journal/v25/n4/abs/nbt1298.html.
- ^ American Red Cross, Southern California Blood Services Region (n.d.). "Answers to Commonly Asked Questions About Blood and Blood Banking" (PDF). Blood: the Basics: 4. http://www.socalredcross.org/pdf/BloodThe.pdf. Retrieved 2007-11-16.
- ^ Klein, Harvey G (March 7, 2005). "Why Do People Have Different Blood Types?". Scientific American. http://www.sciam.com/article.cfm?id=why-do-people-have-differ. Retrieved 2007-11-16.
Further reading
- Dean L (2005). "Chapter 5: The ABO blood group.". Blood Groups and Red Cell Antigens. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=rbcantigen.chapter.ch05ABO. Retrieved 2007-03-24.
- Farr A (1 April 1979). "Blood group serology--the first four decades (1900--1939)". Med Hist 23 (2): 215–26. PMC 1082436. PMID 381816. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1082436.
External links
- ABO at BGMUT Blood Group Antigen Gene Mutation Database at NCBI, NIH
- Encyclopaedia Britannica, ABO blood group system
- National Blood Transfusion Service
- Molecular Genetic Basis of ABO
Transfusion medicine General concepts Blood group systems/
blood typesBlood products/
blood donationCategories:- Blood antigen systems





Wikimedia Foundation. 2010.