- Clade
-
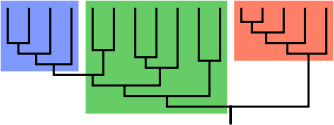 Cladogram (family tree) of a biological group. The red and blue boxes represent clades (i.e., complete branches). The green box is not a clade, but rather represents an evolutionary grade, an incomplete group, because the blue clade is descended from it, but is excluded.
Cladogram (family tree) of a biological group. The red and blue boxes represent clades (i.e., complete branches). The green box is not a clade, but rather represents an evolutionary grade, an incomplete group, because the blue clade is descended from it, but is excluded.
A clade[note 1] is a group consisting of a species (extinct or extant) and all its descendants. In the terms of biological systematics, a clade is a single "branch" on the "tree of life".[1] The idea that such a "natural group" of organisms should be grouped together and given a taxonomic name is central to biological classification. In cladistics (which takes its name from the term), clades are the only acceptable units.
The term was coined in 1958 by English biologist Julian Huxley.[1]
Contents
Definitions

Clade and ancestor
A clade is termed monophyletic, meaning it contains one ancestor (which can be an organism, a population, or a species) and all its descendants.[note 2][2][3] The term clade refers to the grouping of the ancestor and its living and/or deceased descendants together. The ancestor can be a theoretical or actual species.
Clade definition
Three methods of defining clades are featured in phylogenetic nomenclature: node-, stem-, and apomorphy-based:
- In node-based definition, clade name A refers to the least inclusive clade containing taxa (or specimens) X, Y, etc., and their common ancestor. The ancestor is the branch point, or node.
- In stem-based definition, A refers to the most inclusive clade containing X, Y, etc., and their common ancestor, down to where Z branches off below A. Taxa are included between the node of A and down to (but not including) the branching point to Z; that is, the stem of A.
- In apomorphy-based definition, A refers to the clade identified by an apomorphy (a trait) found in X, Y, etc., and their common ancestor.
In Linnaean taxonomy, clades are defined by a set of traits (apomorphies) unique to the group. This system is basically similar to the apomorphy-based clades of phylogenetic nomenclature. The difference is one of weight: While phylogenetic nomenclature bases the group on an ancestor with a certain trait, Linnaean taxonomy uses the traits themselves to define the group.
Clades as constructs
In cladistics, the clade is a hypothetical construct based on experimental data. Clades are found using multiple (sometimes hundreds) of traits from a number of species (or specimens) and analysing them statistically to find the most likely phylogenetic tree for the group.[4] Although similar in some ways to a biological classification of species, the method is statistical and thus directly open to scrutiny and reinterpretation. With changing phylogenetic analysis, the actual content of any defined clade involved may change as well.[5] Although taxonomists use clades as a tool in classification where feasible, the taxonomic "tree of life" is not the same as the cladistic. The traditional genus, family, etc. names are not necessarily clades; though they will often be.
Clade names
In Linnaean systematics, the various groups are ordered into a series of taxonomic ranks (the familiar order, family etc.). These ranks will by convention dictate the ending to names for some groups. Clades do not by their nature fit this scheme, and no such restriction exists as to their names in cladistics. There is however a convention for naming more or less inclusive groups, which are given prefixes like crown- or pan-, see Crown group.
Taxonomy and systematics
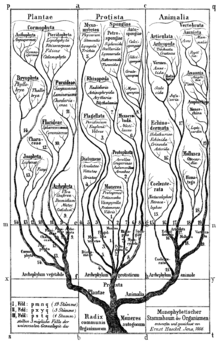
The idea of a "clade" did not exist in pre-Darwinian Linnaean taxonomy, which was based by necessity only on internal or external morphological similarities between organisms – although as it happens, many of the better known animal groups in Linnaeus' original Systema Naturae (notably among the vertebrate groups) do represent clades. The phenomenon of convergent evolution is however responsible for many cases where there are misleading similarities in the morphology of groups that evolved from different lineages.
With the publication of Darwin's theory of evolution in 1859, taxonomy gained a theoretical basis, and the idea was born that groups used in a system of classification should represent branches on the evolutionary tree of life. In the century and a half since then, taxonomists have worked to make the taxonomic system reflect evolution. However, partly because the Tree of Life branches rather unevenly, the hierarchy of the Linnaean system does not always lend itself well to representing clades. The result is that when it comes to naming, cladistics and Linnaean taxonomy are not always compatible. In particular, higher level taxa in Linnaean taxonomy often represent evolutionary grades rather than clades, resulting in groups made up of clades where one or two sub-branches have been excluded. Typical examples include bony fishes, which include the ancestor of tetrapods, and (within the tetrapods) reptiles, which include the ancestors of both birds and mammals.[note 3]
In phylogenetic nomenclature, clades can be nested at any level, and do not have to be slotted into a small number of ranks in an overall hierarchy. In contrast, the Linnaean units of "order", "class" etc. must be used when naming a new taxon. As there are only seven formal levels to the Linnaean system (species being the lowest), only a finite number of sub- and super-units can be created. In order to be able to use the full complexity of taxonomic trees (cladograms) in an area with which they are very familiar, some researchers have opted to dispense with ranks all together, instead using clade names without Linnaean ranks. The reason for preferring one system over the other is partly one of application: cladistic trees give details, suitable for specialists; the Linnaean system gives a well ordered overview, at the expense of details of the phylogenetic tree.
In a few instances, the Linnaean system has actually impeded our understanding of the phylogeny and broad evolutionary patterns. The best known example is the interpretation of the strange fossils of the Burgess Shale and the subsequent idea of a "Cambrian Explosion" [6] With the application of cladistics, and the rejection of any significance of the concept of phyla, the confusion of the late 20th century over the fossils of the Burgess Shale has been resolved. It appears there never was an "explosion" of major bauplans with subsequent extinctions.[7] The seemingly weird Burgess Shale animals have been found to be representatives of a group known as the Lobopodia, that includes arthropods, water bears and velvet worms.[8]
In most instances the two systems are not at odds, however. The cladistic statement, that the clade Lobopodia contains (among others) the Arthropoda, Tardigrada and Onychophora, is factually identical to the Linnaean evolutionary statement that the group Lobopodia is ancestral to the phyla Arthropoda, Tardigrada and Onychophora. The difference is one of semantics rather than phylogeny.
See also
- Cladistics
- Phylogeny
- Paraphyly
- Polyphyly
- Phylogenetic nomenclature
- Binomial nomenclature
- Biological classification
- Crown group
Notes
- ^ (from Ancient Greek κλάδος, klados, "branch")
- ^ A semantic case has been made that the name should be "holophyletic," but this term has not acquired widespread use. For more information, see holophyly
- ^ The term "reptile" is here to be understood as traditionally defined, e.g. Romer & Parson (1985): The Vertebrate Body. (6th ed.) Saunders, Philadelphia. There are other (cladistic) definitions of "reptile" that exclude the first amniotes and the synapsid line, see Sauropsida.
References
- ^ a b Dupuis, Claude (1984). "Willi Hennig's impact on taxonomic thought". Annual Review of Ecology and Systematics 15: 1–24. ISSN 0066-4162.
- ^ "The PhyloCode, Chapter 1". International Society for Phylogenetic Nomenclature. 2009. http://www.ohio.edu/phylocode/art1-3.html#chapter1. Retrieved 23 January 2010.
- ^ Envall, Mats (2008). "On the difference between mono-, holo-, and paraphyletic groups: a consistent distinction of process and pattern". Biological Journal of the Linnaean Society 94: 217. doi:10.1111/j.1095-8312.2008.00984.x.
- ^ Hyvönen, Jaako (2005). "Monophyly, consensus, compromise" (pdf). University of Helsinki. http://www.helsinki.fi/~jhyvonen/ec05/05_11.11.pdf. Retrieved 15 January 2010.
- ^ Nixon, Kevin C.; Carpenter, James M. (2000). "On the Other "Phylogenetic Systematics"". Cladistics 16 (3): 298–318. doi:10.1111/j.1096-0031.2000.tb00285.x.
- ^ Budd, G.E.; Jensen, S. (2000). "A critical reappraisal of the fossil record of the bilaterian phyla". Biological Reviews 75 (2): 253–295. doi:10.1017/S000632310000548X. PMID 10881389. http://journals.cambridge.org/production/action/cjoGetFulltext?fulltextid=624.
- ^ Erwin, D.H. (2007). "Disparity: Morphological Pattern And Developmental Context". Palaeontology 50: 57. doi:10.1111/j.1475-4983.2006.00614.x.
- ^ Whittle, R. J.; Gabbott, S. E.; Aldridge, R. J.; Theron, J. (2009). "An Ordovician Lobopodian from the Soom Shale Lagerstätte, South Africa". Palaeontology 52 (3): 561–567. doi:10.1111/j.1475-4983.2009.00860.x.
External links
- Evolving Thoughts: Clade
- DM Hillis, D Zwickl & R Gutell. "Tree of life". An unrooted cladogram depicting around 3000 species.
- Phylogenetic systematics, an introductory slide-show on evolutionary trees University of California, Berkeley
Topics in phylogenetics Relevant fields Basic concepts Inference methods Maximum parsimony · Maximum likelihood · Neighbor-joining · UPGMA · Bayesian inference · Least squares · Three-taxon analysisCurrent topics -morphy -phyly List of evolutionary biology topics Basic topics in evolutionary biology Evidence of common descent Processes of evolution Population genetic mechanisms Evolutionary developmental
biology (Evo-devo) conceptsEvolution of organs
and biological processesAging · Avian flight · Cellular · DNA · Eye · Flagella · Hair · Human intelligence · Mammalian auditory ossicles · Mosaic evolution · Multicellular · Nervous Systems · SexTaxa evolution Modes of speciation History of evolutionary thought Charles Darwin · On the Origin of Species · Modern evolutionary synthesis · Gene-centered view of evolution · Life (classification trees)Other subfields List of evolutionary biology topics · Timeline of evolution Categories:

Wikimedia Foundation. 2010.