- Evolution of cetaceans
-
 The approximately 80 modern species in the order Cetacea.
The approximately 80 modern species in the order Cetacea.
The cetaceans (whales, dolphins and porpoises) are marine mammal descendants of land mammals. Their terrestrial origins are indicated by:
- Their need to breathe air from the surface;
- The bones of their fins, which resemble the limbs of land mammals
- The vertical movement of their spines, characteristic more of a running mammal than of the horizontal movement of fish.
The question of how land animals evolved into ocean-going leviathans was a mystery until discoveries starting in the late 1970s in Pakistan revealed several stages in the transition of cetaceans from land to sea.
Contents
Earliest ancestors
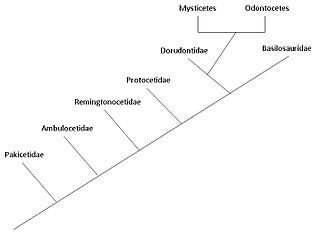
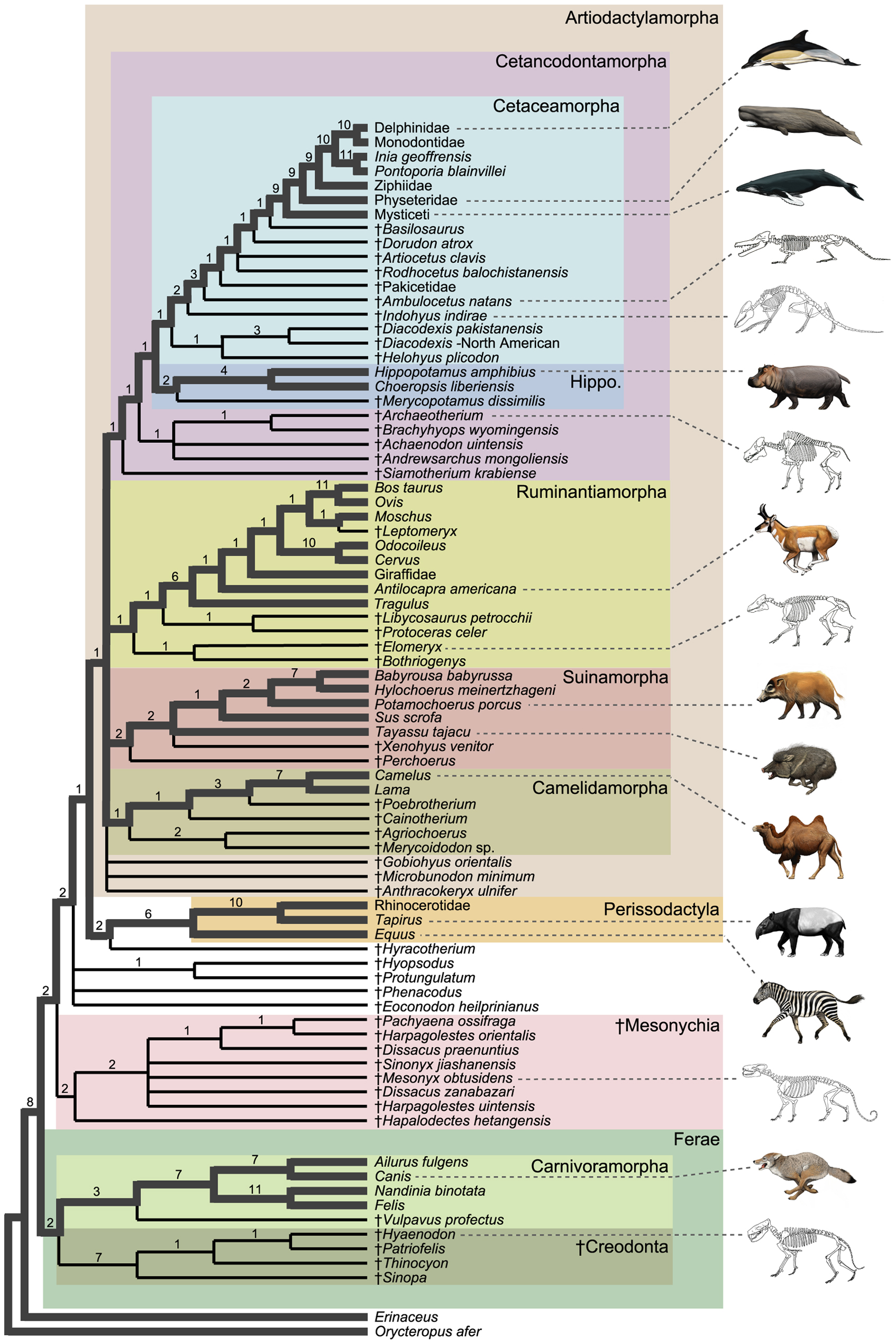
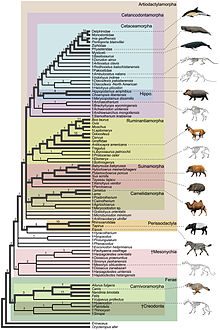 Cladogram showing the position of cetacea within Artiodactylamorpha
Cladogram showing the position of cetacea within ArtiodactylamorphaThe traditional theory of cetacean evolution was that whales were related to the mesonychids, an extinct order of carnivorous ungulates (hoofed animals), which resembled wolves with hooves and were a sister group of artiodactyls (even-toed ungulates). These animals had unusual triangular teeth similar to those of whales. This is why scientists long believed that whales evolved from a form of mesonychid. But more recent molecular phylogeny data suggest that whales are more closely related to the artiodactyls, specifically the hippopotamus.[2] The strong evidence for a clade combining cetaceans and artiodactyls is further discussed in the article Cetartiodactyla. However, the anthracothere ancestors of hippos do not appear in the fossil record until millions of years after Pakicetus, the first known whale ancestor.
The molecular data is supported by the recent discovery of Pakicetus, the earliest proto-whale (see below). The skeletons of Pakicetus show that whales did not derive directly from mesonychids. Instead, they are artiodactyls that began to take to the water soon after artiodactyls split from mesonychids. Proto-whales retained aspects of their mesonychid ancestry (such as the triangular teeth) which modern artiodactyls have lost. An interesting implication is that the earliest ancestors of all hoofed mammals were probably at least partly carnivorous or scavengers, and today's artiodactyls and perissodactyls became herbivores later in their evolution. By contrast, whales retained their carnivorous diet, because prey was more available and they needed higher caloric content in order to live as marine endotherms. Mesonychids also became specialized carnivores, but this was likely a disadvantage because large prey was not yet common. This is why they were out-competed by better-adapted animals like the creodonts and later Carnivora which filled the gaps left by the dinosaurs.
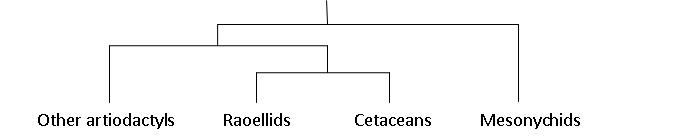
Indohyus
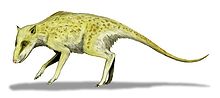
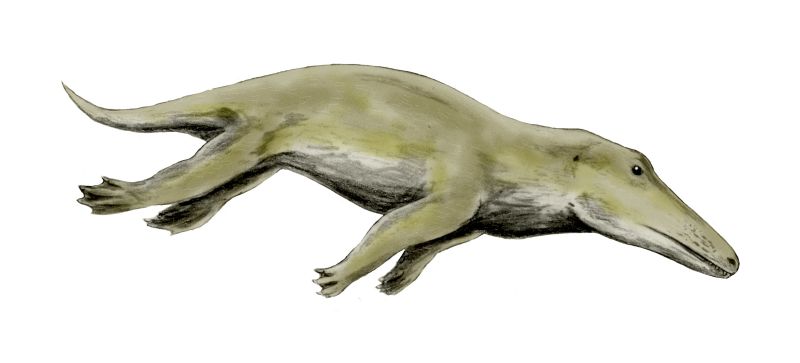
Indohyus is a small deer-like creature, which lived about 48 million years ago in Kashmir.[4] It belongs to the artiodactyls family Raoellidae, and is believed to be the closest sister group of Cetacea.[1] About the size of a raccoon or domestic cat, this herbivorous creature shared some of the traits of whales, most notably the involucrum, which is the diagnostic characteristic of any cetacean, and is not found in any other species.[1] Besides, it also showed signs of adaptations to aquatic life, including a thick and heavy outer coating which is similar to the bones of modern creatures such as the hippopotamus,[2][5] and reduces buoyancy so that they can stay underwater. This suggests a similar survival strategy to the African mousedeer or water chevrotain which, when threatened by a bird of prey, dives into water and hides beneath the surface for up to four minutes.[6][7][8]
Pakicetidae

General
The pakicetids are hoofed mammals that are the earliest whales, with Indohyus from family Raoellidae being the closest sister group.[4] [9] They lived in the early Eocene, around 53 million years ago. Their fossils were first discovered in North Pakistan in 1979, located at a river not far from the shores of former Tethys Sea.[10] After the initial discovery, more fossils were found, mainly in the late-early Eocene fluvial deposits in northern Pakistan and northwestern India.[1] Based on this discovery, pakicetids most likely lived in an arid environment with ephemeral streams and moderately developed floodplains millions of years ago.[1] By using stable oxygen isotopes analysis, they were shown to drink fresh water.[11] Their diet probably included land animals that approached water for drinking or some freshwater aquatic organisms that lived in the river.[1]
Skull morphology
Pakicetids were classified as cetaceans mainly based on the structure of the auditory bulla, which is formed from the ectotympanic bone only. The shape of the ear region in pakicetids is highly unusual and the skull is cetacean-like, although a blowhole is still absent at this stage. The jawbone of pakicetids also lacks the enlarged space (mandibular foramen) that is filled with fat or oil, which is used in receiving underwater sound in modern whales.[12] They have dorsal orbits (eye sockets facing up), which are similar to crocodiles. This eye placement helps submerged predators observe potential prey above the water.[11] According to Thewissen et al., the teeth of pakicetids also resemble the teeth of fossil whales, being less like a dog's incisors, with a serrated triangular shape, similar to a shark's tooth, which is another link to more modern whales.[13] It was initially thought that the ears of pakicetids were adapted for underwater hearing, but, as would be expected from the anatomy of the rest of this creature, the ears of pakicetids are specialized for hearing on land.[14] However, pakicetids were able to listen underwater, by using enhanced bone conduction, rather than depending on tympanic membrane like general land mammals. This method of hearing does not give directional hearing underwater.[12]
Postcranial morphology
Pakicetids have long thin legs, with relatively short hands and feet which suggest that they were poor swimmers.[1] To compensate for that, their bones are unusually thick (osteosclerotic), which is probably an adaptation to make the animal heavier to counteract the buoyancy of the water.[15] According to a morphological analysis by Thewissen et al., pakicetids display no aquatic skeletons adaptation; instead they display adaptations for running and jumping.[16] Hence pakicetids were most likely an aquatic wader.
Ambulocetidae


General
In 1994, Ambulocetus natans, which lived about 49 million years ago, was discovered in Pakistan. It was probably amphibious, and resembled the crocodile in its physical appearance.[16] In the Eocene, ambulocetids inhabited the bays and estuaries of the Tethys Ocean in northern Pakistan.[1] The fossils of ambulocetids are always found in near-shore shallow marine deposits associated with abundant marine plant fossils and littoral molluscs.[1] Although they are found only in marine deposits, their oxygen isotope values indicate that they consumed a range of water with different degree of salinity, with some specimens having no evidence of sea water consumption and others did not ingest fresh water at the time when their teeth are fossilized. It is clear that ambulocetids tolerated a wide range of salt concentrations.[11] Hence, ambulocetids represent the transition phase of cetacean ancestors between fresh water and marine habitat.
Skull morphology
The mandibular foramen in ambulocetids had increased in size, which indicates that a fat pad was likely to be housed in the lower jaw. In modern whales, this fat pad in the mandibular foramen extends posteriorly to the middle ear. This allows sounds to be received in the lower jaw, and then transmitted through the fat pad to the middle ear.[11] Similar to pakicetids, the orbits of ambulocetids are on the dorsal side of the skull, but they face more laterally than in pakicetids.[11]
Postcranial morphology
Ambulocetids had relatively long limbs with particular strong hind legs, and they retained a tail with no sign of fluke (the horizontal tail fin of modern cetaceans).[10] Although they could walk on land, as well as swim, it is clear that they were not fast on either terrain.[17] It has been speculated that Ambulocetids hunted like crocodiles, lurking in the shallows to snatch unsuspecting riparian prey and fish.[16] They probably swam by pelvic paddling (a way of swimming which mainly utilizes their hind limbs to generate propulsion in water) and caudal undulation (a way of swimming which uses the undulations of the vertebral column to generate force for movements), as otters, seals and whales do. This is an intermediate stage in the evolution of cetacean locomotion, as modern whales swim by caudal oscillation (a way of swimming similar to caudal undulation, but uses energy more efficiently).[11]
Remingtonocetidae

General
Remingtonocetids lived in middle-Eocene South Asia, about 49 to 43 million years ago.[18] Compared to family Pakicetidae and Ambulocetidae, Remingtonocetidae was a diverse family found in north and central Pakistan, and also western India.[1] Remingtonocetids were also found in shallow marine deposits, but they are obviously more aquatic than ambulocetids. This can be seen from the recovery of their fossils from a variety of coastal marine environments, including near-shore and lagoonal deposits.[1] It is shown that most remingtonocetids did not ingest fresh water, and had hence lost their dependency on fresh water relatively soon after their origin.[11]
Skull morphology
The orbits of remingtonocetids face laterally and are small. This suggests that vision is not an important sense for them.[11] The nasal opening, which will later evolve to become blowhole in modern cetaceans, is located near the tip of the long snout. The position of the nasal opening had remained unchanged since pakicetids.[11] One of the notable features in remingtonocetids is that the semicircular canals, which are important for balancing in land mammals, had decreased in size.[19] This reduction in size had closely accompanied the cetacean invasion of marine environments. According to Spoor et al., this modification of the semicircular canal system may represents a crucial ‘point of no return’ event in early cetacean evolution, which excluded a prolonged semi-aquatic phase.[19]
Postcranial morphology
Compared to ambulocetids, remingtonocetids had relatively short fore and hind limbs.[11] Based on their skeletal remains, remingtonocetids were probably amphibious whales that are well adapted to swimming, and likely to swim by caudal undulation only.[1]
Protocetidae

General
The protocetids form a diverse and heterogeneous group known from Asia, Europe, Africa, and North America. They lived in the Eocene, approximately 48 to 35 million years ago.[11] The fossil remains of protocetids were uncovered from coastal and lagoonal facies in South Asia; but unlike previous cetacean families, their fossils uncovered from Africa and North America also include open marine forms.[1] Hence they were probably amphibious, but more aquatic compared to remingtonocetids.[18] Protocetids were the first whales to leave the Indian subcontinent and disperse to all shallow subtropical oceans of the world.[11] There were many genera among the family Protocetidae , and some of these are very well known (e.g., Rodhocetus). Great variations in aquatic adaptations exist among them, with some probably able to support their weight on land, whereas others could not.[1] Their supposed amphibious nature is supported by the discovery of a pregnant Maiacetus,[20] in which the fossilised fetus was positioned for a head-first delivery, suggesting that Maiacetus gave birth on land.
Skull morphology

Unlike remingtonocetids and ambulocetids, protocetids have large orbits and are oriented laterally. Increasingly lateral facing eyes might be used to observe prey that live underwater, which are similar to modern cetacean.[11] Furthermore, the nasal openings are large and are now halfway up the snout.[11] The great variety of teeth suggests diverse feeding modes in protocetids.[18] In both remingtonocetids and protocetids, the size of mandibular foramen had increased.[11] The large mandibular foramen indicates that the mandibular fat pad was present. However air-filled sinuses that are present in modern cetaceans, which function to isolate the ear acoustically to enable better underwater hearing, is still not present.[12] The external auditory meatus (ear canal) which is absent in modern cetaceans is also present. Hence, the method of sound transmission present in them combines aspects of pakicetids and modern odontocetes.[12] At this intermediate stage of hearing development, the transmission of airborne sound was poor due to the modifications of ear for underwater hearing; while directional underwater hearing was also poor compared to modern cetaceans.[12]
Postcranial morphology
Some protocetids had short, large fore- and hindlimbs that are likely to be used in swimming, but the limbs give a slow and cumbersome locomotion on land.[11][17] It is possible that some protocetids had flukes.[17] However, it is clear that they are adapted even further to an aquatic life-style. In Rodhocetus, for example, the sacrum (a bone that in land-mammals is a fusion of five vertebrae that connects the pelvis with the rest of the vertebral column) was divided into loose vertebrae. However, the pelvis was still connected to one of the sacral vertebrae. The ungulate ancestry of these early whales is still underlined by characteristics like the presence of hooves at the ends of the toes in Rodhocetus.
Basilosauridae and Dorudontinae

General
Basilosaurids were discovered in 1840 and initially mistaken for a reptile, hence its name. Together with dorudontids, they lived in the late Eocene around 41 to 35 million years ago, and are the oldest known obligate aquatic cetaceans.[1][14] They were fully recognizable whales which lived entirely in the ocean. This is supported by their fossils usually found in deposits indicative of fully marine environments, lacking any freshwater infux.[1] They were probably distributed throughout the tropical and subtropical seas of the world.[1] Basilosaurids are commonly found in association with dorudontids. In fact, they are closely related to one another.[11] The fossilised stomach contents in one basilosaurid indicates that it ate fish.[1]
Skull morphology
Although they look very much like modern whales, basilosaurids and dorudontids lacked the 'melon organ' that allows their descendants to use echolocation as effectively as modern whales. They had small brains; this suggests they were solitary and did not have the complex social structure of some modern cetaceans. The mandibular foramen of basilosaurids and dorudontids now cover the entire depth of the lower jaw as in modern cetaceans.[11] Their orbits face laterally, and the nasal opening had moved even higher up the snout, closer to the position of blowhole in modern cetaceans.[11] Furthermore, their ear structures are functionally modern, with the major innovation being the insertion of air-filled sinuses between ear and skull.[12] Unlike modern cetaceans, basilosaurids retain a large external auditory meatus.[12]

Postcranial morphology
Both basilosaurids and dorudontids have skeletons that are immediately recognizable as cetaceans. A basilosaurid was as big as the larger modern whales, up to 18 m (60 ft) long; dorudontids were smaller, about 5 m (16 ft) long. The large size of basilosaurids is due to the extreme elongation of their lumbar vertebrae. They had a tail fluke, but their body proportions suggest that it swam by caudal undulation and that the fluke was not the propulsive organ.[1] In contrast, dorudontids had a shorter but powerful vertebral column. They too had a fluke, and unlike basilosaurids, they probably swam similarly to modern cetaceans, by using caudal oscillations.[1][11] The forelimbs of basilosaurids and dorudontids were probably flipper-shaped, and the external hind limbs were tiny and are certainly not involved in locomotion.[1] Their fingers, on the other hand, still retain the mobile joints of their ambulocetid relatives. The two tiny but well-formed hind legs of basilosaurids which were probably used as claspers when mating; they are a small reminder of the lives of their ancestors. Interestingly, the pelvic bones associated with these hind limbs was now no longer connected to the vertebral column as it was in protocetids. Essentially, any sacral vertebrae can no longer be clearly distinguished from the other vertebrae.
Ancestry of modern Cetacea
Both basilosaurids and dorudontids are relatively closely related to modern cetacean, which belong to order Odontoceti and Mysticeti. However, according to Fordyce and Barnes, the large size and elongated vertebral body of basilosaurid preclude it from being the ancestor of the two modern orders. As for dorudontids, there are some species within the family that do not have elongated vertebral bodies, which might be the immediate ancestors of Odontoceti and Mysticeti.[18]
Early echolocation

Toothed whales (Odontocetes) echolocate by creating a series of clicks emitted at various frequencies. Sound pulses are emitted through their melon-shaped foreheads, reflected off objects, and retrieved through the lower jaw. Skulls of Squalodon show evidence for the first hypothesized appearance of echolocation.[citation needed] Squalodon lived from the early to middle Oligocene to the middle Miocene, around 33-14 million years ago. Squalodon featured several commonalities with modern Odontocetes. The cranium was well compressed, the rostrum telescoped outward (a characteristic of the modern suborder Odontoceti), giving Squalodon an appearance similar to that of modern toothed whales. However, it is thought unlikely that squalodontids are direct ancestors of living dolphins.
Early baleen whales

All modern mysticetes are large filter-feeding or baleen whales, though the exact means by which baleen is used differs among species (gulp-feeding with balaenopterids, skim-feeding with balaenids, and bottom ploughing with eschrichtiids). The first members of some modern groups appeared during the middle Miocene. These changes may have been a result of worldwide environmental change and physical changes in the oceans. A large scale change in ocean current and temperature could have initiated the radiation of modern mysticetes, leading to the demise of the archaic forms. Generally it is speculated the four modern mysticete families have separate origins among the cetotheres. Modern baleen whales, Balaenopteridae (rorquals and humpback whale, Megaptera novaengliae), Balaenidae (right whales), Eschrichtiidae (gray whale, Eschrictius robustus), and Neobalaenidae (pygmy right whale, Caperea marginata) all have derived characteristics presently unknown in any cetothere.
Early dolphins
 Skeleton of Xiphiacetus sp.
Skeleton of Xiphiacetus sp.During the early Miocene (about 20 Ma), echolocation developed in its modern form. Various extinct dolphin-like families flourished. Early dolphins include Kentriodon and Hadrodelphis. These belong to Kentriodontidae, which were small to medium-sized toothed cetaceans with largely symmetrical skulls, and thought likely to include ancestors of some modern species. Kentriodontids date to the late Oligocene to late Miocene. Kentriodontines ate small fish and other nectonic organisms; they are thought to have been active echolocators, and might have formed schools. Diversity, morphology and distribution of fossils appear parallel to some modern species.
Skeletal evolution
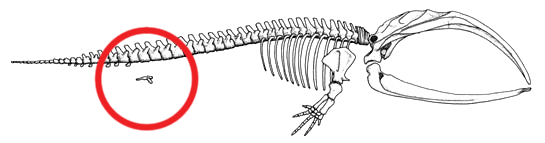
 The skeleton of a Baleen whale with the hind limb and pelvic bone structure circled in red. This bone structure stays internal during the entire life of the species.
The skeleton of a Baleen whale with the hind limb and pelvic bone structure circled in red. This bone structure stays internal during the entire life of the species.Today, the whale hind parts are internal and reduced. Occasionally, the genes that code for longer extremities cause a modern whale to develop miniature legs (known as atavism).
Whereas early cetaceans such as the Pakicetus had the nasal openings at the end of the snout, in later species such as the Rodhocetus, the openings had begun to drift toward the top of the skull. This is known as nasal drift.
The nostrils of modern whales have become modified into blowholes that allow them to break to the surface, inhale, and submerge with convenience. The ears began to move inward as well, and, in the case of Basilosaurus, the middle ears began to receive vibrations from the lower jaw. Today's modern toothed whales use the 'melon organ', a pad of fat, for echolocation.
See also
- Dolphin, Evolution and anatomy
- Archaeoceti
- Aquatic adaptation
- Indohyus
- Transitional form
- Evolution of sirenians
- Evolution of mammals
- List of extinct cetaceans
References
- ^ a b c d e f g h i j k l m n o p q r s t u v Thewissen, J. G. M.; Williams, E. M. (1 November 2002). "THE EARLY RADIATIONS OF CETACEA (MAMMALIA): Evolutionary Pattern and Developmental Correlations". Annual Review of Ecology and Systematics 33 (1): 73–90. doi:10.1146/annurev.ecolsys.33.020602.095426.
- ^ a b University Of California, Berkeley (2005, February 7). "UC Berkeley, French Scientists Find Missing Link Between The Whale And Its Closest Relative, The Hippo". ScienceDaily. http://www.sciencedaily.com/releases/2005/02/050205103109.htm. Retrieved 2007-12-21.
- ^ Thewissen, J. G. M.; Cooper, Lisa Noelle, Clementz, Mark T., Bajpai, Sunil, Tiwari, B. N. (20 December 2007). "Whales originated from aquatic artiodactyls in the Eocene epoch of India". Nature 450 (7173): 1190–1194. Bibcode 2007Natur.450.1190T. doi:10.1038/nature06343. PMID 18097400.
- ^ a b Northeastern Ohio Universities Colleges of Medicine and Pharmacy (2007, December 21). "Whales Descended From Tiny Deer-like Ancestors". ScienceDaily. http://www.sciencedaily.com/releases/2007/12/071220220241.htm. Retrieved 2007-12-21.
- ^ University Of Michigan (2001, September 20). "New Fossils Suggest Whales And Hippos Are Close Kin". ScienceDaily. http://www.sciencedaily.com/releases/2001/09/010920072245.htm. Retrieved 2007-12-21.
- ^ Carl Zimmer (2007-12-19). "The Loom : Whales: From So Humble A Beginning..". ScienceBlogs. http://scienceblogs.com/loom/2007/12/19/whales_from_so_humble_a_beginn.php. Retrieved 2007-12-21.
- ^ Ian Sample (2007-12-19). "Whales may be descended from a small deer-like animal". Guardian Unlimited. http://www.guardian.co.uk/science/2007/dec/19/whale.deer?gusrc=rss&feed=networkfront. Retrieved 2007-12-21.
- ^ PZ Myers (2007-12-19). "Pharyngula: Indohyus". Pharyngula. ScienceBlogs. http://scienceblogs.com/pharyngula/2007/12/indohyus.php. Retrieved 2007-12-21.
- ^ Philip D. Gingerich, D. E. Russell (1981). "Pakicetus inachus, a new archaeocete (Mammalia, Cetacea) from the early-middle Eocene Kuldana Formation of Kohat (Pakistan)". Univ. Mich. Contr. Mus. Paleont 25: 235–246.
- ^ a b Castro, E. Huber, Peter, Michael (2003). Marine Biology (4 ed). McGraw-Hill.
- ^ a b c d e f g h i j k l m n o p q r s t THEWISSEN, J. G. M.; BAJPAI, SUNIL (1 January 2001). "Whale Origins as a Poster Child for Macroevolution". BioScience 51 (12): 1037. doi:10.1641/0006-3568(2001)051[1037:WOAAPC]2.0.CO;2. ISSN 0006-3568.
- ^ a b c d e f g Nummela, Sirpa; Thewissen, J. G. M., Bajpai, Sunil, Hussain, S. Taseer, Kumar, Kishor (11 August 2004). "Eocene evolution of whale hearing". Nature 430 (7001): 776–778. Bibcode 2004Natur.430..776N. doi:10.1038/nature02720. PMID 15306808.
- ^ Whale Origins
- ^ a b J. G. M. Thewissen, E. M. Williams, L. J. Roe and S. T. Hussain (2001). "Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls". Nature 413 (6853): 277–281. doi:10.1038/35095005. PMID 11565023.
- ^ Thewissen, J. G. M.; Cooper, Lisa Noelle, Clementz, Mark T., Bajpai, Sunil, Tiwari, B. N. (20 December 2007). "Whales originated from aquatic artiodactyls in the Eocene epoch of India". Nature 450 (7173): 1190–1194. Bibcode 2007Natur.450.1190T. doi:10.1038/nature06343. PMID 18097400.
- ^ a b c Thewissen, J. G. M.; Williams, E. M., Roe, L. J., Hussain, S. T. (19 September 2001). "Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls". Nature 413 (6853): 277–281. doi:10.1038/35095005. PMID 11565023.
- ^ a b c "The Thewissen Lab". http://www.neoucom.edu/DEPTS/ANAT/Thewissen/whale_origins/index.html. Retrieved 12 Feb 2011.
- ^ a b c d Fordyce, R E; Barnes, L G (30 April 1994). "The Evolutionary History of Whales and Dolphins". Annual Review of Earth and Planetary Sciences 22 (1): 419–455. Bibcode 1994AREPS..22..419F. doi:10.1146/annurev.ea.22.050194.002223.
- ^ a b Spoor, F.; Bajpai, S., Hussain, S. T., Kumar, K., Thewissen, J. G. M. (8 May 2002). "Vestibular evidence for the evolution of aquatic behaviour in early cetaceans". Nature 417 (6885): 163–166. doi:10.1038/417163a. PMID 12000957.
- ^ Gingerich PD, ul-Haq M, von Koenigswald W, Sanders WJ, Smith BH, et al.. "New Protocetid Whale from the Middle Eocene of Pakistan: Birth on Land, Precocial Development, and Sexual Dimorphism". PLoS one. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0004366. Retrieved 2009-02-04.
External links
For a review of whale evolution, see Uhen, M. D. (2010). "The Origin(s) of Whales". Annual Review of Earth and Planetary Sciences 38: 189–219. Bibcode 2010AREPS..38..189U. doi:10.1146/annurev-earth-040809-152453.
- Timeline of Whale Evolution - Smithsonian Ocean Portal
- Cetacean Paleobiology – University of Bristol
- BBC: Whale's evolution
- BBC: Whale Evolution – The Fossil Evidence
- Hooking Leviathan by Its Past by Stephen Jay Gould
- Whale Origins, Thewissen Lab, Northeastern Ohio Universities College of Medicine
- Digital Library of Dolphin Development, Thewissen Lab
- Research on the Origin and Early Evolution of Whales (Cetacea), Gingerich, P.D., University of Michigan
- Pakicetus inachus, a new archaeocete (Mammalia, Cetacea) from the early-middle Eocene Kuldana Formation of Kohat (Pakistan). Gingerich, P.D., 1981, Museum of Paleontology, The University of Michigan
- Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls, Nature 413, 277–281 (20 September 2001), J. G. M. Thewissen, E. M. Williams, L. J. Roe and S. T. Hussain
- Evolution of Whales segment from the Whales Tohorā Exhibition Minisite of the Museum of New Zealand Te Papa Tongarewa
Basic topics in evolutionary biology Evidence of common descent Processes of evolution Population genetic mechanisms Evolutionary developmental
biology (Evo-devo) conceptsEvolution of organs
and biological processesAging · Avian flight · Cellular · DNA · Eye · Flagella · Hair · Human intelligence · Mammalian auditory ossicles · Mosaic evolution · Multicellular · Nervous Systems · SexTaxa evolution Modes of speciation History of evolutionary thought Charles Darwin · On the Origin of Species · Modern evolutionary synthesis · Gene-centered view of evolution · Life (classification trees)Other subfields List of evolutionary biology topics · Timeline of evolution Categories:- Cetaceans
- Evolution by taxon










Wikimedia Foundation. 2010.