- Speciation
-
Part of a series on Evolutionary Biology  Key TopicsHistory of TheoryNatural HistoryProcesses & OutcomesSocial ImplicationsFields & ApplicationsApplications of evolution
Key TopicsHistory of TheoryNatural HistoryProcesses & OutcomesSocial ImplicationsFields & ApplicationsApplications of evolution
Artificial selection
Bioinformatics
Evolutionary anthropology
Evolutionary computation
Evolutionary dynamics
Experimental evolution
Ecological genetics
Evolutionary neuroscience
Evolutionary physiology
Evolutionary psychology
Phylogenetics
SystematicsEvolutionary Biology Portal
Category • Related topics • BookSpeciation is the evolutionary process by which new biological species arise. The biologist Orator F. Cook seems to have been the first to coin the term 'speciation' for the splitting of lineages or 'cladogenesis,' as opposed to 'anagenesis' or 'phyletic evolution' occurring within lineages.[1][2] Whether genetic drift is a minor or major contributor to speciation is the subject matter of much ongoing discussion.
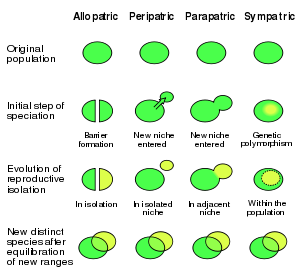
There are four geographic modes of speciation in nature, based on the extent to which speciating populations are geographically isolated from one another: allopatric, peripatric, parapatric, and sympatric. Speciation may also be induced artificially, through animal husbandry or laboratory experiments. Observed examples of each kind of speciation are provided throughout.[3]
Contents
Natural speciation
All forms of natural speciation have taken place over the course of evolution; however it still remains a subject of debate as to the relative importance of each mechanism in driving biodiversity.[4]
One example of natural speciation is the diversity of the three-spined stickleback, a marine fish that, after the last ice age, has undergone speciation into new freshwater colonies in isolated lakes and streams. Over an estimated 10,000 generations, the sticklebacks show structural differences that are greater than those seen between different genera of fish including variations in fins, changes in the number or size of their bony plates, variable jaw structure, and color differences.[5]
Speciation Rate
There is debate as to the rate at which speciation events occur over geologic time. While some evolutionary biologists claim that speciation events have remained relatively constant over time, some palaeontologists such as Niles Eldredge and Stephen Jay Gould have argued that species usually remain unchanged over long stretches of time, and that speciation occurs only over relatively brief intervals, a view known as punctuated equilibrium.
Allopatric
Main article: allopatric speciationDuring allopatric (from the ancient Greek allos, "other" + Greek patrā, "fatherland") speciation, a population splits into two geographically isolated populations (for example, by habitat fragmentation due to geographical change such as mountain building). The isolated populations then undergo genotypic and/or phenotypic divergence as: (a) they become subjected to dissimilar selective pressures; (b) they independently undergo genetic drift; (c) different mutations arise in the two populations. When the populations come back into contact, they have evolved such that they are reproductively isolated and are no longer capable of exchanging genes.
- Observed instances
Island genetics, the tendency of small, isolated genetic pools to produce unusual traits, has been observed in many circumstances, including insular dwarfism and the radical changes among certain famous island chains, for example on Komodo. The Galápagos islands are particularly famous for their influence on Charles Darwin. During his five weeks there he heard that Galápagos tortoises could be identified by island, and noticed that Mockingbirds differed from one island to another, but it was only nine months later that he reflected that such facts could show that species were changeable. When he returned to England, his speculation on evolution deepened after experts informed him that these were separate species, not just varieties, and famously that other differing Galápagos birds were all species of finches. Though the finches were less important for Darwin, more recent research has shown the birds now known as Darwin's finches to be a classic case of adaptive evolutionary radiation.[6]
Peripatric
Main article: Peripatric speciationIn peripatric speciation, a subform of allopatric speciation, new species are formed in isolated, smaller peripheral populations that are prevented from exchanging genes with the main population. It is related to the concept of a founder effect, since small populations often undergo bottlenecks. Genetic drift is often proposed to play a significant role in peripatric speciation.
- Observed instances
- Mayr bird fauna[citation needed]
- The Australian bird Petroica multicolor
- Reproductive isolation occurs in populations of Drosophila subject to population bottlenecking
Parapatric
Main article: Parapatric speciationIn parapatric speciation, there is only partial separation of the zones of two diverging populations afforded by geography; individuals of each species may come in contact or cross habitats from time to time, but reduced fitness of the heterozygote leads to selection for behaviours or mechanisms that prevent their inter-breeding. Parapatric speciation is modelled on continuous variation within a 'single', connected habitat acting as a source of natural selection rather than the effects of isolation of habitats produced in peripatric and allopatric speciation.
Ecologists refer to parapatric and peripatric speciation in terms of ecological niches. A niche must be available in order for a new species to be successful.
- Observed instances
- Ring species
- The Larus gulls form a ring species around the North Pole.
- The Ensatina salamanders, which form a ring round the Central Valley in California.
- The Greenish Warbler (Phylloscopus trochiloides), around the Himalayas.
- the grass Anthoxanthum has been known to undergo parapatric speciation in such cases as mine contamination of an area.
Sympatric
Main article: Sympatric speciationSympatric speciation refers to the formation of two or more descendant species from a single ancestral species all occupying the same geographic location.
In sympatric speciation, species diverge while inhabiting the same place. Often-cited examples of sympatric speciation are found in insects that become dependent on different host plants in the same area.[7][8] However, the existence of sympatric speciation as a mechanism of speciation is still hotly contested. People have argued that the evidences of sympatric speciation are in fact examples of micro-allopatric, or heteropatric speciation. The most widely accepted example of sympatric speciation is that of the cichlids of Lake Nabugabo in East Africa, which is thought to be due to sexual selection.
Until recently, there has a been a dearth of strong evidence that supports this form of speciation, with a general feeling that interbreeding would soon eliminate any genetic differences that might appear. But there has been at least one recent study that suggests that sympatric speciation has occurred in Tennessee cave salamanders.[9]
Sympatric speciation driven by ecological factors may also account for the extraordinary diversity of crustaceans living in the depths of Siberia's Lake Baikal.
Example of three-spined sticklebacks
 The three-spined stickleback (Gasterosteus aculeatus)
The three-spined stickleback (Gasterosteus aculeatus)
The three-spined sticklebacks, freshwater fishes, that have been studied by Dolph Schluter (who received his Ph.D. for his work on Darwin's finches with Peter J. Grant) and his current colleagues in British Columbia, were once thought to provide an intriguing example best explained by sympatric speciation. Schluter and colleagues found:
- Two different species of three-spined sticklebacks in each of five different lakes
- DNA analysis indicates that each lake was colonized independently, presumably by a marine ancestor, after the last ice age
- DNA analysis also shows that the two species in each lake are more closely related to each other than they are to any of the species in the other lakes
- The two species in each lake are reproductively isolated; neither mates with the other.
- However, aquarium tests showed:
- the benthic species from one lake will spawn with the benthic species from the other lakes and
- likewise the limnetic species from the different lakes will spawn with each other.
- These benthic and limnetic species even display their mating preferences when presented with sticklebacks from Japanese lakes; that is, a Canadian benthic prefers a Japanese benthic over its close limnetic cousin from its own lake.
- Their conclusion: in each lake, what began as a single population faced such competition for limited resources that:
- disruptive selection — competition favoring fishes at either extreme of body size and mouth size over those nearer the mean — coupled with:
- assortative mating — each size preferred mates like it — favored a divergence into two subpopulations exploiting different food in different parts of the lake.
- The fact that this pattern of speciation occurred the same way on three separate occasions suggests strongly that ecological factors in a sympatric population can cause speciation.
However, the DNA evidence cited above is from mitochondrial DNA (mtDNA), which can often move easily between closely related species ("introgression") when they hybridize. A more recent study,[10] using genetic markers from the nuclear genome, shows that limnetic forms in different lakes are more closely related to each other (and to marine lineages) than to benthic forms in the same lake. The three-spine stickleback is now usually considered an example of "double invasion" (a form of allopatric speciation) in which repeated invasions of marine forms have subsequently differentiated into benthic and limnetic forms. The three-spine stickleback provides an example of how molecular biogeographic studies that rely solely on mtDNA can be misleading, and that consideration of the genealogical history of alleles from multiple unlinked markers (i.e. nuclear genes) is necessary to infer speciation histories.
Speciation via polyploidization
Polyploidy is a mechanism that has caused many rapid speciation events in sympatry because offspring of, for example, tetraploid x diploid matings often result in triploid sterile progeny.[11] However, not all polyploids are reproductively isolated from their parental plants, and gene flow may still occur for example through triploid hybrid x diploid matings that produce tetraploids, or matings between meiotically unreduced gametes from diploids and gametes from tetraploids (see also hybrid speciation).
It has been suggested that many of the existing plant and most animal species have undergone an event of polyploidization in their evolutionary history.[12][13] Reproduction of successful polyploid species is sometimes asexual, by parthenogenesis or apomixis, as for unknown reasons many asexual organisms are polyploid. Rare instances of polyploid mammals are known, but most often result in prenatal death.
Hawthorn fly
One example of evolution at work is the case of the hawthorn fly, Rhagoletis pomonella, also known as the apple maggot fly, which appears to be undergoing sympatric speciation.[14] Different populations of hawthorn fly feed on different fruits. A distinct population emerged in North America in the 19th century some time after apples, a non-native species, were introduced. This apple-feeding population normally feeds only on apples and not on the historically preferred fruit of hawthorns. The current hawthorn feeding population does not normally feed on apples. Some evidence, such as the fact that six out of thirteen allozyme loci are different, that hawthorn flies mature later in the season and take longer to mature than apple flies; and that there is little evidence of interbreeding (researchers have documented a 4-6% hybridization rate) suggests that sympatric speciation is occurring. The emergence of the new hawthorn fly is an example of evolution in progress.[15]
Speciation via hybrid formation
See Hybrid speciation section under the Genetics heading beneath.
Reinforcement
Reinforcement, also called Wallace effect, is the process by which natural selection increases reproductive isolation.[16] It may occur after two populations of the same species are separated and then come back into contact. If their reproductive isolation was complete, then they will have already developed into two separate incompatible species. If their reproductive isolation is incomplete, then further mating between the populations will produce hybrids, which may or may not be fertile. If the hybrids are infertile, or fertile but less fit than their ancestors, then there will be further reproductive isolation and speciation has essentially occurred (e.g., as in horses and donkeys.)
The reasoning behind this is that if the parents of the hybrid offspring each have naturally selected traits for their own certain environments, the hybrid offspring will bear traits from both, therefore would not fit either ecological niche as well as either parent. The low fitness of the hybrids would cause selection to favor assortative mating, which would control hybridization. This is sometimes called the Wallace effect after the evolutionary biologist Alfred Russel Wallace who suggested in the late 19th century that it might be an important factor in speciation.[17]
Conversely, if the hybrid offspring are more fit than their ancestors, then the populations will merge back into the same species within the area they are in contact.Reinforcement favoring reproductive isolation is required for both parapatric and sympatric speciation. Without reinforcement, the geographic area of contact between different forms of the same species, called their "hybrid zone," will not develop into a boundary between the different species. Hybrid zones are regions where diverged populations meet and interbreed. Hybrid offspring are very common in these regions, which are usually created by diverged species coming into secondary contact. Without reinforcement, the two species would have uncontrollable inbreeding. Reinforcement may be induced in artificial selection experiments as described below.
Artificial speciation
New species have been created by domesticated animal husbandry, but the initial dates and methods of the initiation of such species are not clear. For example, domestic sheep were created by hybridisation, and no longer produce viable offspring with Ovis orientalis, one species from which they are descended.[18] Domestic cattle, on the other hand, can be considered the same species as several varieties of wild ox, gaur, yak, etc., as they readily produce fertile offspring with them.[19]
The best-documented creations of new species in the laboratory were performed in the late 1980s. William Rice and G.W. Salt bred fruit flies, Drosophila melanogaster, using a maze with three different choices of habitat such as light/dark and wet/dry. Each generation was placed into the maze, and the groups of flies that came out of two of the eight exits were set apart to breed with each other in their respective groups. After thirty-five generations, the two groups and their offspring were isolated reproductively because of their strong habitat preferences: they mated only within the areas they preferred, and so did not mate with flies that preferred the other areas.[20] The history of such attempts is described in Rice and Hostert (1993).[21]
Diane Dodd was also able to show how reproductive isolation can develop from mating preferences in Drosophila pseudoobscura fruit flies after only eight generations using different food types, starch and maltose.[22]

Dodd's experiment has been easy for many others to replicate, including with other kinds of fruit flies and foods.[23]
Genetics
Few speciation genes have been found. They usually involve the reinforcement process of late stages of speciation. In 2008 a speciation gene causing reproductive isolation was reported.[24] It causes hybrid sterility between related subspecies.
Hybrid speciation
Main article: Hybrid speciationHybridization between two different species sometimes leads to a distinct phenotype. This phenotype can also be fitter than the parental lineage and as such natural selection may then favor these individuals. Eventually, if reproductive isolation is achieved, it may lead to a separate species. However, reproductive isolation between hybrids and their parents is particularly difficult to achieve and thus hybrid speciation is considered an extremely rare event. The Mariana Mallard is known to have arisen from hybrid speciation.
Hybridisation is an important means of speciation in plants, since polyploidy (having more than two copies of each chromosome) is tolerated in plants more readily than in animals.[25][26] Polyploidy is important in hybrids as it allows reproduction, with the two different sets of chromosomes each being able to pair with an identical partner during meiosis.[27] Polyploids also have more genetic diversity, which allows them to avoid inbreeding depression in small populations.[28]
Hybridization without change in chromosome number is called homoploid hybrid speciation. It is considered very rare but has been shown in Heliconius butterflies [29] and sunflowers. Polyploid speciation, which involves changes in chromosome number, is a more common phenomenon, especially in plant species.
Gene transposition as a cause
Theodosius Dobzhansky, who studied fruit flies in the early days of genetic research in 1930s, speculated that parts of chromosomes that switch from one location to another might cause a species to split into two different species. He mapped out how it might be possible for sections of chromosomes to relocate themselves in a genome. Those mobile sections can cause sterility in inter-species hybrids, which can act as a speciation pressure. In theory, his idea was sound, but scientists long debated whether it actually happened in nature. Eventually a competing theory involving the gradual accumulation of mutations was shown to occur in nature so often that geneticists largely dismissed the moving gene hypothesis.[30]
However, 2006 research shows that jumping of a gene from one chromosome to another can contribute to the birth of new species.[31] This validates the reproductive isolation mechanism, a key component of speciation.[32]
Interspersed repeats
Main article: Interspersed repeatInterspersed repetitive DNA sequences function as isolating mechanisms. These repeats protect newly evolving gene sequences from being overwritten by gene conversion, due to the creation of non-homologies between otherwise homologous DNA sequences. The non-homologies create barriers to gene conversion. This barrier allows nascent novel genes to evolve without being overwritten by the progenitors of these genes. This uncoupling allows the evolution of new genes, both within gene families and also allelic forms of a gene. The importance is that this allows the splitting of a gene pool without requiring physical isolation of the organisms harboring those gene sequences.
In 2011, it has been proposed that rapid amplification of repetitive DNA by genetic drift in small subpopulations can help them to "drift apart" which may lead to the origin of new species.[33]
Human speciation
Humans have genetic similarities with chimpanzees and bonobos, their closest relatives, suggesting common ancestors. Analysis of genetic drift and recombination using a Markov model suggests humans and chimpanzees speciated apart 4.1 million years ago.[34]
See also
- Extinction
- Species problem
- Heteropatry
- Chronospecies
- Koinophilia
- Assortative mating
- Court Jester Hypothesis
References
- ^ Cook O. F. (1906). "Factors of species-formation". Science 23 (587): 506–507. doi:10.1126/science.23.587.506. PMID 17789700.
- ^ Cook O. F. (1908). "Evolution without isolation". American Naturalist 42: 727–731. doi:10.1086/279001.
- ^ Observed Instances of Speciation by Joseph Boxhorn. Retrieved 8 June 2009.
- ^ J.M. Baker (2005). "Adaptive speciation: The role of natural selection in mechanisms of geographic and non-geographic speciation". Studies in History and Philosophy of Biological and Biomedical Sciences 36 (2): 303–326. doi:10.1016/j.shpsc.2005.03.005. PMID 19260194.
- ^ Kingsley, D.M. (January 2009) "From Atoms to Traits," Scientific American, p. 57
- ^ Frank J. Sulloway (1982). "The Beagle collections of Darwin's finches (Geospizinae)". Bulletin of the British Museum (Natural History) Zoology Series 43 (2): 49–58. available online
- ^ Feder, J. L., X. Xie, J. Rull, S. Velez, A. Forbes, B. Leung, H. Dambroski, K. E. Filchak, and M. Aluja. 2005. Mayr, Dobzhansky, and Bush and the complexities of sympatric speciation in Rhagoletis. Proceedings of the National Academy of Sciences, USA 1902:6573-6580
- ^ Berlocher, S. H., and J. L. Feder. 2002. Sympatric speciation in phytophagous insects: moving beyond controversy? Annual Review of Entomology 47:773-815
- ^ MATTHEW L. NIEMILLER, BENJAMIN M. FITZPATRICK, BRIAN T. MILLER (2008). "Recent divergence with gene flow in Tennessee cave salamanders (Plethodontidae: Gyrinophilus) inferred from gene genealogies". Molecular Ecology 17 (9): 2258–2275. doi:10.1111/j.1365-294X.2008.03750.x. PMID 18410292.
- ^ E.B. TAYLOR, J.D. McPHAIL (2000). "Historical contingency and determinism interact to prime speciation in sticklebacks". Proceedings of the Royal Society of London Series B 267 (1460): 2375–2384. doi:10.1098/rspb.2000.1294. PMC 1690834. PMID 11133026. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1690834.[1] available online
- ^ Ramsey, J., and D. W. Schemske. 1998. Pathways, mechanisms, and rates of polyploid formation in flowering plants. Annual Review of Ecology and Systematics 29:467-501
- ^ Otto S.P., Whitton J. (2000). "Polyploidy: incidence and evolution". Annual Review of Genetics 34: 401–437. doi:10.1146/annurev.genet.34.1.401. PMID 11092833.
- ^ Comai L (2005). "The advantages and disadvantages of being polyploid". Nature Reviews Genetics 6 (11): 836–846. doi:10.1038/nrg1711. PMID 16304599.
- ^ Feder JL, Roethele JB, Filchak K, Niedbalski J, Romero-Severson J (1 March 2003). "Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella". Genetics 163 (3): 939–53. PMC 1462491. PMID 12663534. http://www.genetics.org/cgi/pmidlookup?view=long&pmid=12663534.
- ^ Berlocher SH, Bush GL (1982). "An electrophoretic analysis of Rhagoletis (Diptera: Tephritidae) phylogeny". Systematic Zoology 31 (2): 136–55. doi:10.2307/2413033. JSTOR 2413033.
- ^ Ridley, M. (2003) "Speciation — What is the role of reinforcement in speciation?" adapted from Evolution 3rd edition (Boston: Blackwell Science) tutorial online
- ^ Ollerton, J. "Flowering time and the Wallace Effect" (PDF). Heredity, August 2005. http://oldweb.northampton.ac.uk/aps/env/lbrg/journals/papers/OllertonHeredityCommentary2005.pdf. Retrieved 2007-05-22.
- ^ Hiendleder, S.; Kaupe, B.; Wassmuth, R.; Janke, A. (2002). "et al. (2002) "Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies". Proceedings of the Royal Society B: Biological Sciences 269 (1494): 893–904. doi:10.1098/rspb.2002.1975. PMC 1690972. PMID 12028771. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1690972.
- ^ Nowak, R. (1999) Walker's Mammals of the World 6th ed. (Baltimore: Johns Hopkins University Press)
- ^ Rice, W.R. and G.W. Salt (1988). "Speciation via disruptive selection on habitat preference: experimental evidence". The American Naturalist 131: 911–917. doi:10.1086/284831.
- ^ W.R. Rice and E.E. Hostert (1993). "Laboratory experiments on speciation: What have we learned in forty years?". Evolution 47 (6): 1637–1653. doi:10.2307/2410209. JSTOR 2410209.
- ^ Dodd, D.M.B. (1989). "Reproductive isolation as a consequence of adaptive divergence in Drosophila pseudoobscura". Evolution 43 (6): 1308–1311. doi:10.2307/2409365. JSTOR 2409365.
- ^ Kirkpatrick, M. and V. Ravigné (2002) "Speciation by Natural and Sexual Selection: Models and Experiments" The American Naturalist 159:S22–S35 DOI
- ^ http://www.sciencemag.org/cgi/content/short/323/5912/376
- ^ Wendel J (2000). "Genome evolution in polyploids". Plant Mol. Biol. 42 (1): 225–49. doi:10.1023/A:1006392424384. PMID 10688139.
- ^ Sémon M, Wolfe KH (2007). "Consequences of genome duplication". Curr Opin Genet Dev 17 (6): 505–12. doi:10.1016/j.gde.2007.09.007. PMID 18006297.
- ^ Comai L (2005). "The advantages and disadvantages of being polyploid". Nat. Rev. Genet. 6 (11): 836–46. doi:10.1038/nrg1711. PMID 16304599.
- ^ Soltis P, Soltis D (2000). "The role of genetic and genomic attributes in the success of polyploids". Proc. Natl. Acad. Sci. U.S.A. 97 (13): 7051–7. doi:10.1073/pnas.97.13.7051. PMC 34383. PMID 10860970. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=34383.
- ^ Mavarez, J.; Salazar, C.A., Bermingham, E., Salcedo, C., Jiggins, C.D. , Linares, M. (2006). "Speciation by hybridization in Heliconius butterflies". Nature 441 (7095): 868–71. doi:10.1038/nature04738. PMID 16778888.
- ^ University of Rochester Press Releases
- ^ Masly, John P., Corbin D. Jones, Mohamed A. F. Noor, John Locke, and H. Allen Orr (September 2006). "Gene Transposition as a Cause of Hybrid Sterility in Drosophila". Science 313 (5792): 1448–1450. doi:10.1126/science.1128721. PMID 16960009. http://www.sciencemag.org/cgi/content/short/313/5792/1448. Retrieved 2007-03-18.
- ^ Minkel, J.R. (September 8, 2006) "Wandering Fly Gene Supports New Model of Speciation" Science News
- ^ Jurka, Jerzy, Weidong Bao, Kenji K. Kojima (September 2011). "Families of transposable elements, population structure and the origin of species". Biology Direct 6: 44. PMC 3183009. PMID 21929767. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3183009.
- ^ Hobolth A, Christensen OF, Mailund T, Schierup MH (2007) "Genomic Relationships and Speciation Times of Human, Chimpanzee, and Gorilla Inferred from a Coalescent Hidden Markov Model." PLoS Genet 3(2): e7 (doi:10.1371/journal.pgen.0030007)
Further reading
- Coyne, J. A. & Orr, H. A. (2004). Speciation. Sunderlands, Massachusetts: Sinauer Associates, Inc. ISBN 0-87893-089-2.
- Grant, V. (1981). Plant Speciation (2nd Edit. ed.). New York: Columbia University Press. ISBN 0-231-05113-1.
- Mayr, E. (1963). Animal Species and Evolution. Harvard University Press. ISBN 0-674-03750-2
- Marko, P. B. (2008). Allopatry. Oxford: Elsevier. ISBN 978-0-444-52033-3.
- White, M. J. D. (1978). Modes of Speciation. San Francisco, California: W. H. Freeman and Company. ISBN 0-716-70284-3.
- Dedicated issue of Philosophical Transactions B on Speciation in microorganisms is freely available.
External links
- Observed Instances of Speciation from the Talk.Origins Frequently Asked Questions
- Speciation, and
- Evidence for Speciation from Understanding Evolution by the University of California Museum of Paleontology
- Speciation from John Hawks' Anthropology Weblog - paleoanthropology, genetics, and evolution
Speciation Basic concepts Modes of speciation Auxiliary mechanisms Intermediate stages Basic topics in evolutionary biology Processes of evolution Population genetic mechanisms Evolutionary developmental
biology (Evo-devo) conceptsEvolution of organs
and biological processesAging · Avian flight · Cellular · DNA · Eye · Flagella · Hair · Human intelligence · Mammalian auditory ossicles · Mosaic evolution · Multicellular · Nervous Systems · SexTaxa evolution Modes of speciation History of evolutionary thought Charles Darwin · On the Origin of Species · Modern evolutionary synthesis · Gene-centered view of evolution · Life (classification trees)Other subfields Categories:




Wikimedia Foundation. 2010.