- Galápagos tortoise
-
Galápagos tortoise 
Conservation status Scientific classification Kingdom: Animalia Phylum: Chordata Class: Reptilia Order: Testudines Suborder: Cryptodira Family: Testudinidae Genus: Chelonoidis Species: C. nigra Binomial name Chelonoidis nigra[1]
(Quoy & Gaimard, 1824b[3])Subspecies Subspecies[2]See: Subspecies of Galápagos tortoise
- C. n. abingdoni (Extinct in the wild)
Günther, 1877[4] - C. n. becki
Rothschild, 1901[5] - C. n. chathamensis
Van Denburgh, 1907[6] - C. n. darwini
Van Denburgh, 1907[6] - C. n. duncanensis
Garman, 1917[7] - C. n. hoodensis
Van Denburgh, 1907[6] - C. n. nigra (Extinct)
Quoy & Gaimard, 1824b[3] - C. n. phantastica (Extinct)
Van Denburgh, 1907[6] - C. n. porteri
Rothschild, 1903[8] - C. n. vicina
Günther, 1875[9]
Synonyms Species synonymy[10]- Testudo californiana
Quoy & Gaimard, 1824a[11] (nomen oblitum) - Testudo nigra
Quoy & Gaimard, 1824b[3] (nomen novum) - Testudo elephantopus
Harlan, 1827[12] (nomen dubium) - Testudo nigrita
Duméril and Bibron, 1834[13] (nomen dubium) - Testudo planiceps
Gray, 1853[14] (nomen dubium) - Testudo clivosa
Garman, 1917[7] (nomen dubium) - Testudo typica
Garman, 1917[7] (nomen dubium) - Testudo (Chelonoidis) elephantopus
Williams, 1952[15] - Geochelone (Chelonoidis) elephantopus
Pritchard, 1967[16] - Chelonoidis elephantopus
Bour, 1980[17]
Subspecies synonymy[1]C. n. nigra (nominate subspecies)
- Testudo californiana
Quoy & Gaimard, 1824a[11] (nomen oblitum) - Testudo nigra
Quoy & Gaimard, 1824b[3] (nomen novum) - Testudo galapagoensis
Baur 1889[18]
C. n. abingdoni
- Testudo ephippium
Günther, 1875[9] (partim, misidentified type specimen once erroneously attributed to what is now C. n. duncanensis) - Testudo abingdoni
Günther, 1877[4]
C. n. becki
- Testudo becki
Rothschild, 1901[5]
C. n. chathamensis
- Testudo wallacei
Rothschild 1902[19] (partim, nomen dubium) - Testudo chathamensis
Van Denburgh, 1907[6]
C. n. darwini
- Testudo wallacei
Rothschild 1902[19] (partim, nomen dubium) - Testudo darwini
Van Denburgh, 1907[6]
C. n. duncanensis
- Testudo ephippium
Günther, 1875[9] (partim, misidentified type) - Geochelone nigra duncanensis
Garman, 1917[7] in Pritchard, 1996[10](nomen nudum)
C. n. hoodensis
- Testudo hoodensis
Van Denburgh, 1907[6]
C. n. phantastica
- Testudo phantasticus
Van Denburgh, 1907[6]
C. n. porteri
- Testudo nigrita
Duméril and Bibron, 1834[13] (nomen dubium) - Testudo porteri
Rothschild, 1903[8]
C. n. vicina
The Galápagos tortoise or Galápagos giant tortoise (Chelonoidis nigra) is the largest living species of tortoise, reaching weights of over 400 kg (880 lb) and lengths of over 1.8 meters (5.9 ft). With life spans in the wild of over 100 years, it is one of the longest-lived vertebrates. A captive individual lived at least 170 years.
The tortoise is native to seven of the Galápagos Islands, a volcanic archipelago about 1,000 km (620 mi) west of the Ecuadorian mainland. Spanish explorers who discovered the islands in the 16th century named them after the Spanish galápago, meaning tortoise.
Shell size and shape vary between populations. On islands with humid highlands, the tortoises are larger, with domed shells and short necks. On islands with dry lowlands, the tortoises are smaller, with "saddleback" shells and long necks. These island-to-island differences played a role in the inception of Charles Darwin's theory of evolution.
Tortoise numbers declined from over 250,000 in the 16th century to a low of around 3,000 in the 1970s. The decline was caused by hunting for tortoise meat and oil, habitat clearance for agriculture, and introduction of non-native animals such as rats, goats, and pigs. Ten subspecies of the original fifteen survive in the wild. An eleventh subspecies (C. n. abingdoni) has only a single living individual, in captivity, nicknamed Lonesome George. Conservation efforts beginning in the 20th century have resulted in thousands of captive-bred juveniles being released onto their home islands, and it is estimated that numbers exceeded 19,000 at the start of the 21st century. Despite this rebound, the species as a whole is classified as "Vulnerable" by the International Union for Conservation of Nature (IUCN).
Contents
Taxonomy
- Early classification
The Galápagos Islands were discovered in 1535, but first appeared on maps, of Gerardus Mercator and Abraham Ortelius, in about 1570.[21] The islands were named "Insulae de los Galopegos" (Islands of the Tortoises) in reference to the giant tortoises found there.[22][23][nb 1]
Initially, the giant tortoises of the Indian Ocean and those from the Galápagos were considered to be the same species. Naturalists thought that sailors had transported the tortoises.[24] In 1676, the pre-Linnaean authority Claude Perrault referred to both species as Tortue des Indes.[25] In 1783, Johann Gottlob Schneider classified all giant tortoises as Testudo indica ("Indian tortoise").[26] In 1812, August Friedrich Schweigger named them Testudo gigantea ("gigantic tortoise").[27] In 1834, André Marie Constant Duméril and Gabriel Bibron classified the Galápagos tortoises as a separate species, which they named Testudo nigrita ("black tortoise").[13]
 Walter Rothschild, cataloger of two Galápagos tortoise subspecies
Walter Rothschild, cataloger of two Galápagos tortoise subspecies
- Recognition of subpopulations
The first systematic survey of giant tortoises was by Albert Günther of the British Museum, in 1875.[9] Günther identified at least five distinct populations from the Galápagos, and three from the Indian Ocean islands. He expanded the list in 1877 to six from the Galápagos, four from the Seychelles, and four from the Mascarenes. Günther theorised that all the giant tortoises descended from a single ancestral population which spread by sunken land bridges.[28] This theory was later disproven by the understanding that the Galápagos, Seychelles and Mascarene islands are all of recent volcanic origin and therefore could not have been linked by land bridges. It is now thought that the Galápagos tortoises descended from an ancestor from South America.[29] The Indian Ocean tortoises derived from Madagascar.[30][31]
At the end of the 19th century, Georg Baur[18] and Walter Rothschild[5][8][19] recognised five more populations of Galápagos tortoise. In 1906, the Academy of Sciences collected specimens and gave them to John Van Denburgh for study. He identified four additional populations,[6] and proposed the existence of 15 species.[32] His list still guides the taxonomy of the Galápagos tortoise, though now ten populations are thought to have existed.[1]
- Current species and genus names
The current species designation of nigra ("black" – Quoy & Gaimard, 1824b[3]) was resurrected in 1984[33] after it was discovered to be the senior synonym (an older taxonomic synonym taking historical precedence) for the then commonly used species name of elephantopus ("elephant footed" – Harlan, 1827[12]). The use of nigra is explained by Quoy and Gaimard's Latin description: "Testudo toto corpore nigro", meaning "tortoise with completely black body". Quoy and Gairmard described nigra from a living specimen, but there is no evidence that they knew of its accurate provenance within the Galápagos – the locality was in fact given as California. Garman proposed the linking of nigra with the extinct Floreana subspecies.[7] Later, Pritchard deemed it was convenient to accept this designation despite its tenuousness because this decision allowed minimal disruption to the already-confused nomenclature of the species. The even more senior species synonym of californiana ("californian" – Quoy & Gaimard, 1824a[11]) is considered a nomen oblitum ("forgotten name").[34]
Previously, the Galápagos tortoise was considered to belong to the genus Geochelone, known as typical tortoises or terrestrial turtles. Subsequently, subgenus Chelonoidis was elevated to generic status based on phylogenetic evidence which grouped the South American members of Geochelone into an independent clade (branch of the tree of life).[35] This nomenclature has been adopted by several authorities.[2][1][36][37]
- Subspecies
Main article: Subspecies of Galápagos tortoiseThe exact number of subspecies of Chelonoidis nigra that have existed in history is still under debate among scientists, but some recognize up to 15.[38] Only ten subspecies now exist in the wild, one on Santiago, Santa Cruz, San Cristóbal, Pinzón and Española, and five on Isabela, although this number is controversial and may be fewer. An eleventh surviving subspecies, abingdoni from Pinta Island, is considered extinct in the wild and is represented by a single living specimen, "Lonesome George". The subspecies inhabiting Floreana island (C. n. nigra) is thought to have been hunted to extinction by 1850,[39][40] only years after Charles Darwin's landmark visit of 1835 in which he saw shells but no live tortoises on the island.[41] The phantastica subspecies of Fernandina is of disputed existence as it was described from a single specimen which may have been an artificial introduction to the island.[42][43][44]
Prior to widespread knowledge of the differences between the populations (sometimes called races) from different islands and volcanoes, captive collections in zoos were indiscriminately mixed. Fertile offspring resulted from pairings of animals from different races, confirming that they are subspecies and not distinct species. However, captive crosses between tortoises from different races have lower fertility and higher mortality than those between tortoises of the same race[45][46] and captives in mixed herds normally direct courtship only toward members of the same race.[46]
The valid scientific names of each of the individual populations are not universally accepted,[10][43][47][48] and some researchers consider each subspecies to be a full species.[49][50] The taxonomic status of the various races is not fully resolved.[51]
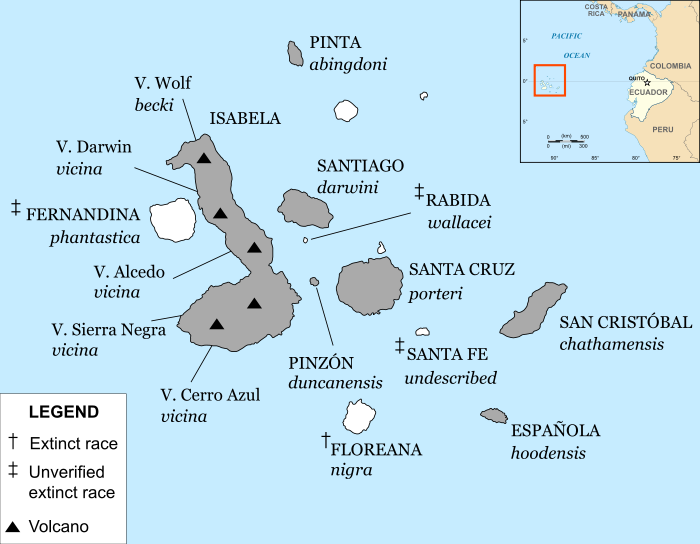 Galápagos archipelago annotated with ranges of currently recognised subspecies of Galápagos tortoise. Islands with surviving subspecies are shaded.
Galápagos archipelago annotated with ranges of currently recognised subspecies of Galápagos tortoise. Islands with surviving subspecies are shaded.Evolutionary history
All subspecies of Galápagos tortoise evolved from common ancestors that arrived from mainland South America by overwater dispersal. The minimal founding population was a pregnant female or a breeding pair.[29] Survival on the 1000 km oceanic journey is accounted for by the fact that the tortoises are buoyant, can breathe by extending their necks above the water, and are able to survive months without food or fresh water. As they are poor swimmers, the journey was probably a passive one facilitated by the Humboldt Current, which diverts westwards towards the Galápagos Islands from the mainland.[38]
The closest living relative (though not a direct ancestor) of the Galápagos giant tortoise is the Argentine tortoise (Chelonoidis chilensis), a much smaller species from South America. The divergence between C. chilensis and C. nigra probably occurred 6–12 million years ago, an evolutionary event preceding the volcanic formation of the oldest modern Galápagos Islands 5 million years ago.[52] Mitochondrial DNA analysis indicates that the oldest existing islands (Española and San Cristóbal) were colonised first, and that these populations seeded the younger islands via dispersal in a "stepping stone" fashion via local currents.[53][54] Restricted gene flow between isolated islands then resulted in the independent evolution of the populations into the divergent forms observed in the modern subspecies. The evolutionary relationships between the subspecies thus echo the volcanic history of the islands.[29]
Subspecies genetics
Modern DNA methods have revealed new information on the relationships between the subspecies:
- Isabela Island
The five populations living on the largest island, Isabela are the ones that are the most highly debated on whether they are true subspecies or just distinct populations. It is widely accepted that the population living on the most northern volcano, Volcan Wolf are genetically independent from the four other southern populations and are therefore their own subspecies.[29] They are thought to be so genetically different from the others because there were two colonization events. One in the north from the island of Santiago resulting in the Volcan Wolf species (G. becki) and the second from the island of Santa Cruz resulting in the four southern populations.[29] It is thought that they were first colonized on the Sierra Negra volcano, which was the first volcano formed on the island. The tortoises then spread north to each subsequently forming volcano, resulting in the populations living on Volcan Alcedo and then Volcan Darwin. Recent genetic evidence shows these two populations are genetically distinct from each other and from the population living on Sierra Negra and therefore form the subspecies G. vandenburghi (Alcedo) and G. microphyes (Darwin).[55] The fifth population living on the most southern volcano is thought to have split off from the Sierra Negra population more recently and is therefore not as genetically different as the other two.[55] Isabela is the youngest island which tortoises inhabit, therefore none of them have had the same amount of time to independently evolve as populations have one other islands, but according to Ciofi and associates they are all genetically different and should each be considered their own subspecies.[55]
- Floreana Island
Phylogenetic analysis may help to "resurrect" the extinct subspecies of Floreana (nigra). The subspecies was only known from subfossil remains.[40] Some tortoises from Isabela were found to be a partial match for the genetic profile of Floreana specimens from museum collections, possibly indicating the presence of hybrids from a population transposed by humans from Floreana to Isabela[50] speculated to be caused either by deliberate moving between the islands[56] or from individuals thrown overboard ships to lighten loads.[18] Nine Floreana descendants have been identified in the captive population of the Fausto Llerena Breeding Center on Santa Cruz. This permits the possibility of re-establishing a reconstructed subspecies from selective breeding of the hybrid animals.[57]
- Pinta Island
The Pinta Island subspecies (abingdoni, now extinct in the wild) is most closely related to the subspecies on the islands of San Cristóbal (chathamensis) and Española (hoodensis) which lie over 300 kilometers (190 mi) away,[29] rather than neighbouring Isabela as previously assumed. This relationship is attributable to dispersal by the strong local current from San Cristóbal towards Pinta.[58] The discovery informed further attempts for the preservation of the abingdoni lineage and the search for an appropriate mate for Lonesome George, who had been penned with females from Isabela.[59] This hope was bolstered by the discovery of an abingdoni hybrid male in the Volcán Wolf population on northern Isabela, raising the possibility that there are more living undiscovered Pinta descendants.[60]
- Santa Cruz Island
Mitochondrial DNA studies of tortoises on Santa Cruz show up to three genetically distinct lineages found in non-overlapping population distributions around the regions of Cerro Monturra, Cerro Fatal and La Caseta.[61] Although currently united in a single subspecies (porteri), the lineages are all more closely related to tortoises on other islands than to each other;[62] Cerro Monturra tortoises are most closely related to duncanensis from Pinzón, Cerro Fatal to chathamensis from San Cristóbal, and La Caseta to the four southern races of Isabela.[63]
- Subspecies of doubtful existence
Subspecies were described from three other islands, but their existence is based on scant evidence. The purported Rabida island subspecies (wallacei) was described from a single specimen removed by the Academy of Sciences in 1906,[32] which has since been lost. This individual was probably an artificial introduction from another island that was originally penned on Rabida next to a good anchorage, as no contemporary whaling or sealing logs mention removing tortoises from this island.[44] The phantastica subspecies from Fernandina is known from a single specimen from the voyage of 1906, an old male.[32] No other tortoises or remains have been found on the island, suggesting the specimen was an artificial introduction from elsewhere.[43][44][56] Fernandina has neither human settlements nor feral mammals, so if this subspecies ever did exist its extinction must have been by natural means, such as volcanic activity.[43] The Santa Fe subspecies has no binomial name, having been described from the limited evidence of bone fragments (but no shells, the most durable part) of 14 individuals, old eggs and old dung found on the island in 1906.[32] The island has never been inhabited by man nor had any introduced predators.[56] The remains are considered artificial introductions,[43] possibly from camping at the good anchorage on the island.[56]
Description
The tortoises have a large bony shell (carapace) of a dull brown colour. The plates of the carapace are fused with the ribs in a rigid protective structure that is integral to the skeleton. Lichens can grow on the shells of the slow-moving animals.[64] Tortoises keep a characteristic scute (shell segment) pattern on their shell throughout life, though the annual growth bands are not useful for determining age, because the outer layers are worn off with time. A tortoise can withdraw its head, neck and forelimbs into its shell for protection. The legs are large, stumpy, with dry scaly skin and hard scales. The front legs are five-clawed, and the back legs are four-clawed.[32]
Gigantism
The discoverer of the Galápagos Islands, Fray Tomás de Berlanga, Bishop of Panama, wrote in 1535 of "such big tortoises that each could carry a man on top of himself."[65] Naturalist Charles Darwin remarked after his trip three centuries later in 1835, "These animals grow to an immense size ... several so large that it required six or eight men to lift them from the ground".[66] The largest recorded individuals have reached weights of over 400 kilograms (880 lb)[67] and lengths of 1.87 meters (6.1 ft).[3][68] The tortoises' gigantism was probably a preadapted condition for successful colonisation of these remote oceanic islands rather than an example of evolved insular gigantism. Large tortoises would have a greater chance of surviving the journey over water from the mainland as they can hold their heads a greater height above the water level and have a smaller surface area/volume ratio, which reduces osmotic water loss. Their significant water and fat reserves would allow the tortoises to survive long ocean crossings without food or fresh water, and to endure the drought-prone climate of the islands. A larger size allowed them to better tolerate extremes of temperature due to gigantothermy.[69] Fossil giant tortoises from mainland South America have been described that support this hypothesis of preadapted gigantism.[70]
Shell shape
Galápagos tortoise shell varieties
 Saddleback (abingdoni)
Saddleback (abingdoni) Intermediate (chathamensis)
Intermediate (chathamensis) Domed (porteri)
Domed (porteri)Galapágos tortoises are the only lineage of giant tortoise exhibiting different types of shell shape.[62] They exhibit a spectrum of carapace morphology ranging from "saddleback" (denoting upward arching of the front edge of the shell resembling a saddle) to "domed" (denoting a rounded convex surface resembling a dome). When saddleback tortoises withdraw their head and forelimbs into their shells, a large unprotected gap remains over their neck, evidence of the lack of predation during the evolution of this structure. There is no saddleback/domed dualism, as tortoises can be of intermediate type with characteristics of both. Larger islands with humid highlands over 800 meters (2,600 ft) in elevation, such as Santa Cruz have abundant vegetation near the ground.[48] Native tortoises in these environments tend to have domed shells and are larger, with shorter necks and limbs. Saddleback tortoises originate from small islands less than 500 meters (1,600 ft) in elevation with dry habitats (e.g. Española and Pinzón) that are more limited in food and other resources.[38]
- Evolutionary implications
In combination with proportionally longer necks and limbs,[32] the unusual saddleback carapace structure is thought to be an adaptation to increase vertical reach, which enables the tortoise to browse tall vegetation, such as the Opuntia (prickly pear) cactus that grows in arid environments.[71] Saddlebacks are more territorial[68][72] and smaller than domed varieties, possibly adaptations to their limited resources. Alternatively, larger tortoises may be better-suited to high elevations, because they can resist the cooler temperatures when there is cloud cover or fog.[47]
A competing hypothesis is that rather than being principally a feeding adaptation, the distinctive saddle shape and longer extremities might have been a secondary sexual characteristic of saddleback males. Male competition over mates is settled by dominance displays on the basis of vertical neck height rather than body size[47] (see below). This correlates with the observation that saddleback males are more aggressive than domed males.[73] The shell distortion and elongation of the limbs and neck in saddlebacks is probably the evolutionary compromise between the need for a small body size in dry conditions and a high vertical reach for dominance displays.[47]
The saddleback carapace probably evolved independently several times in dry habitats,[68] since genetic similarity between populations does not correspond to carapace shape.[74] Saddleback tortoises are therefore not necessarily more closely related to each other than to domed counterparts, as shape is not determined by a similar genetic background, but by a similar ecological one.[47]
- Sexual dimorphism
Sexual dimorphism is most pronounced in saddleback populations, in which males have more angled and higher front openings, giving a more extreme saddled appearance.[73] Males of all varieties generally have a longer tail and a shorter, concave undershell with thickened knobs at the back edge to facilitate mating. Males are larger than females: adult males weigh around 272–317 kilograms (600–700 lb) while females are 136–181 kilograms (300–400 lb).[47]
Behaviour
 A tortoise bathing in a pool
A tortoise bathing in a pool- Routine
The tortoises are ectothermic (cold-blooded) and therefore bask for 1–2 hours after dawn to absorb the sun's heat through their dark shells before actively foraging for 8–9 hours a day.[43] They travel mostly in the early morning or late afternoon between resting and grazing areas. They have been observed to walk at a speed of 0.3 kilometres per hour (0.2 mph).[66]
On the larger and more humid islands, the tortoises seasonally migrate down between low elevations, which become grassy plains in the wet season, to meadowed areas of higher elevation (up to 2,000 ft (610 m)[32]) in the dry season. The same routes have been used for many generations, creating well-defined paths through the undergrowth known as "tortoise highways".[48] On these wetter islands, the domed tortoises are gregarious and often found in large herds, in contrast to the more solitary and territorial disposition of the saddleback tortoises.
Tortoises sometimes rest in mud wallows or rain-formed pools, which may be both a thermoregulatory response during cool nights, and a protection from parasites such as mosquitoes and ticks.[48] Parasites are countered by taking dust baths in loose soil. Tortoises have been noted to shelter at night under overhanging rocks.[75] Other tortoises are observed to sleep in a snug depression in the earth or brush called a "pallet". Local tortoises using the same pallet sites, such as on Volcán Alcedo, results in the formation of small sandy pits.[76]
- Diet
 A tortoise feeding
A tortoise feedingThe tortoises are herbivores that consume a diet of cacti, grasses, leaves, lichens, and berries. They have been documented to feed on Hippomane mancinella ('poison apple'), the endemic guava Psidium galapageium, the water fern Azolla microphylla, and the bromeliad Tillandsia insularis.[77] A tortoise eats an average of 32–36 kilograms (70–80 lb) per day, though inefficient digestion means that much of this passes through without nutritional extraction.[78]
Tortoises acquire most of their moisture from the dew and sap in vegetation (particularly the Opuntia cactus); therefore, they can spend long periods without drinking water. They can endure 18 months when deprived of all food and water,[79] surviving by breaking down their body fat to produce water as a by-product. When thirsty they may drink large quantities of water very quickly, storing it in their bladders and the "root of the neck" (the pericardium[43]), both of which used to make them useful water sources on ships.[79] On arid islands, tortoises lick morning dew from boulders, and the repeated action over many generations has formed half-sphere depressions in the rock.[43]
- Senses
Regarding their senses, Charles Darwin observed that: "The inhabitants believe that these animals are absolutely deaf; certainly they do not overhear a person walking near behind them. I was always amused, when overtaking one of these great monsters as it was quietly pacing along, to see how suddenly, the instant I passed, it would draw in its head and legs, and uttering a deep hiss fall to the ground with a heavy sound, as if struck dead."[66] Although they are not deaf,[32] tortoises depend far more on vision and smell as stimuli.[48]
- Mutualism
Tortoises share a mutualistic relationship with some species of Galápagos finch and mockingbirds. Small groups of finches initiate the process by hopping on the ground in an exaggerated fashion facing the tortoise. The tortoise signals it is ready by rising up and extending its neck and legs, enabling the birds to reach otherwise inaccessible spots on the tortoise's body such as the neck, rear legs, cloacal opening, and skin between plastron and carapace. The birds benefit from the food source and the tortoises get rid of irritating ectoparasites.[80]
Some tortoises have been observed to insidiously exploit this mutualistic relationship. After rising and extending its limbs, the bird may go beneath the tortoise to investigate, whereupon suddenly the tortoise withdraws its limbs to drop flat and kill the bird. It then steps back to eat the bird, presumably to supplement its diet with protein.[81]
- Mating
 A pair of tortoises engaging in a dominance display
A pair of tortoises engaging in a dominance displayMating occurs at any time of the year, although it does have seasonal peaks between February and June in the humid uplands during the rainy season.[48] When mature males meet in the mating season they will face each other in a ritualised dominance display, rise up on their legs and stretch up their necks with their mouths gaping open. Occasionally, head-biting occurs, but usually the shorter tortoise will back off, conceding mating rights to the victor. The behaviour is most pronounced in saddleback subspecies, which are more aggressive and have longer necks.[73]
The prelude to mating can be very aggressive, as the male forcefully rams the female's shell with his own and nips her legs.[20] Mounting is an awkward process and the male must stretch and tense to maintain equilibrium in a slanting position. The concave underside of the male's shell helps him to balance when straddled over the female's shell, and brings his cloacal vent (which houses the penis) closer to the female's dilated cloaca. During mating, the male vocalises with hoarse bellows and grunts,[75] described as "rhythmic groans".[48] This is one of the few vocalisations the tortoise makes; other noises are made during aggressive encounters, when struggling to right themselves, and hissing as they withdraw into their shells due to the forceful expulsion of air.[82]
 A young tortoise and a tortoise egg
A young tortoise and a tortoise egg- Egg-laying
Females then journey up to several kilometres in July to November to reach nesting areas of dry, sandy coast. Nest digging is a tiring and elaborate task which may take the female several hours a day over many days to complete.[48] It is carried out blindly using only the hind legs to dig a 30 cm (12 in) deep cylindrical hole, into which she lays up to sixteen spherical, hard-shelled eggs ranging from 82 to 157 grams (2.9 to 5.5 oz) in mass,[43] and the size of a billiard ball.[64] Some observations suggest that the average clutch size for domed populations (9.6 per clutch for porteri on Santa Cruz) is larger than that of saddlebacks (4.6 per clutch for duncanensis on Pinzón).[44] The female makes a muddy plug for the nest hole out of soil mixed with urine, seals the nest by pressing down firmly with her plastron, and leaves them to be incubated by the sun. Females may lay 1–4 clutches per season. Temperature plays a role in the sex of the hatchling, with lower temperature nests producing more males and higher temperature nests producing more females. This is related closely to incubation time, since clutches laid early will incubate during the cool season and have longer incubation periods (producing more males), while nests that are laid later incubate for a shorter period in the hot season (producing more females).[83]
- Early life and maturation
Young animals emerge from the nest after four to eight months and may weigh only 50 grams (1.8 oz) and measure 6 centimetres (2.4 in).[48] When the young tortoises emerge from their shells, they must dig their way to the surface, which can take up several weeks, though their yolk sac can sustain them for up to seven months.[64] In particularly dry conditions, the hatchlings may die underground if they are encased by hardened soil, while flooding of the nest area can drown them. Subspecies are initially indistinguishable as they all have domed carapaces. The young stay in warmer lowland areas for their first 10–15 years,[43] encountering hazards such as falling into cracks, being crushed by falling rock, or excessive heat stress. The Galápagos Hawk was formerly the sole native predator of the tortoise hatchlings, as Darwin wrote: "The young tortoises, as soon as they are hatched, fall prey in great numbers to the buzzard".[66] The hawk is now much rarer, but introduced feral pigs, dogs, cats and black rats have become predators of eggs and young tortoises.[84] The adult tortoises have no natural predators apart from humans, as Darwin noted: "The old ones seem generally to die from accidents, as from falling down precipices. At least several of the inhabitants told me, they had never found one dead without some such apparent cause".[66]
Sex can be determined only when the tortoise is about 15 years old, and sexual maturity is reached at around 20–25 years in captivity, possibly 40 years in the wild (when they reach their full size). Life expectancy in the wild is thought to be over 100 years,[85][86] making it one of the longest lived species in the animal kingdom. Harriet was the oldest known Galápagos tortoise, having reached an estimated age of more than 170 years before her death in 2006 in Australia Zoo.[87] Chambers notes that Harriet was probably 169 years old in 2004. The individual died in 2006 (although media outlets claimed the greater age of 175 based on a less reliable timeline.[88]
Darwin's development of theory of evolution
 Charles Darwin as a young man, probably subsequent to the Galápagos visit
Charles Darwin as a young man, probably subsequent to the Galápagos visitCharles Darwin visited the Galápagos for five weeks on the second voyage of HMS Beagle in 1835 and saw Galápagos tortoises on San Cristobal (Chatham) and Santiago (James) Islands.[89] They appeared several times in his writings and journals, and played a role in the development of the theory of evolution.
Darwin wrote in his account of the voyage:
"I have not as yet noticed by far the most remarkable feature in the natural history of this archipelago; it is, that the different islands to a considerable extent are inhabited by a different set of beings. My attention was first called to this fact by the Vice-Governor, Mr. Lawson, declaring that the tortoises differed from the different islands, and that he could with certainty tell from which island any one was brought ... The inhabitants, as I have said, state that they can distinguish the tortoises from the different islands; and that they differ not only in size, but in other characters. Captain Porter has described* those from Charles and from the nearest island to it, namely, Hood Island, as having their shells in front thick and turned up like a Spanish saddle, while the tortoises from James Island are rounder, blacker, and have a better taste when cooked."[90]
The significance of the differences in tortoises between islands did not strike him as important until it was too late, as he continued,
"I did not for some time pay sufficient attention to this statement, and I had already partially mingled together the collections from two of the islands. I never dreamed that islands, about fifty or sixty miles apart, and most of them in sight of each other, formed of precisely the same rocks, placed under a quite similar climate, rising to a nearly equal height, would have been differently tenanted".[90]
Although the Beagle departed from the Galápagos with over 30 adult tortoises on deck, these were not for scientific study but a source of fresh meat for their Pacific crossing. Their shells and bones were thrown overboard, leaving no remains with which to test any hypotheses.[91] It has been suggested[92] that this oversight was made because Darwin only reported seeing tortoises on San Cristóbal[93] (chathamensis) and Santiago[94] (darwini), both of which have an intermediate type of shell shape and are not particularly morphologically distinct from each other. Though he did visit Floreana, the nigra subspecies found there was already nearly extinct and he was unlikely to have seen any mature animals.[41]
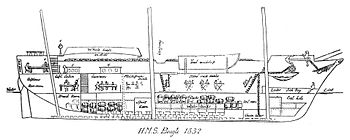 Line drawing of the HMS Beagle by F. G. King, a colleague of Darwin's. Galápagos tortoises were stacked in the lower hold.
Line drawing of the HMS Beagle by F. G. King, a colleague of Darwin's. Galápagos tortoises were stacked in the lower hold.Darwin did however have four live juvenile specimens to compare from different islands. These were pet tortoises taken by himself (from San Salvador), his captain FitzRoy (two from Española) and his servant Syms Covington (from Floreana).[95] Unfortunately they could not help to determine whether each island had its own variety because the specimens were not mature enough to exhibit morphological differences.[96] Although the British Museum had a few specimens, their provenance within the Galápagos was unknown.[97] However, conversations with the naturalist Gabriel Bibron, who had seen the mature tortoises of the Paris Natural History Museum confirmed to Darwin that there were distinct varieties.[98]
Darwin later compared the different tortoise forms with those of mockingbirds, in the first[99] tentative statement linking his observations from the Galapagos with the possibility of species transmuting:
"When I recollect the fact that [from] the form of the body, shape of scales and general size, the Spaniards can at once pronounce from which island any tortoise may have been brought; when I see these islands in sight of each other and possessed of but a scanty stock of animals, tenanted by these birds, but slightly differing in structure and filling the same place in nature; I must suspect they are only varieties ... If there is the slightest foundation for these remarks, the zoology of archipelagos will be well worth examining; for such facts would undermine the stability of species."[100]
His views on the mutability of species were restated in his notebooks: "animals on separate islands ought to become different if kept long enough apart with slightly differing circumstances. – Now Galapagos Tortoises, Mocking birds, Falkland Fox, Chiloe fox, – Inglish and Irish Hare."[101] These observations served as counterexamples to the prevailing contemporary view that species were individually created.
Darwin also found these "antediluvian animals"[93] to be a source of diversion: "I frequently got on their backs, and then giving a few raps on the hinder part of their shells, they would rise up and walk away;—but I found it very difficult to keep my balance".[66]
Conservation
Several waves of human exploitation of the tortoises as a food source caused a decline in the total wild population from around 250,000[86] when first discovered in the 16th century to a low of 3,060 individuals in a 1974 census. Modern conservation efforts have subsequently brought tortoise numbers up to 19,317 (estimate for 1995–2009).[102]
The subspecies C. n. nigra became extinct by human exploitation in the 19th century. Another subspecies, C. n. abingdoni, is now extinct in the wild and represented in captivity by a single male specimen, Lonesome George. It is the only known living specimen of the Pinta Island tortoise and the world's "rarest living creature".[103] All the other surviving subspecies are listed by the IUCN as at least "Vulnerable" in conservation status, if not worse.
Historical exploitation
An estimated 200,000 animals were taken before the 20th century.[43] The relatively immobile and defenceless tortoises were collected and stored live on board ships, where they could survive for at least a year without food or water (some anecdotal reports suggest individuals surviving two years[104]), providing valuable fresh meat, while their diluted urine and water stored in their neck bags could be used as drinking water. The 17th century British pirate, explorer and naturalist William Dampier wrote that "They are so extraordinarily large and fat, and so sweet, that no pullet eats more pleasantly,"[105] while Captain James Colnett of the British Navy wrote of "the land tortoise which in whatever way it was dressed, was considered by all of us as the most delicious food we had ever tasted."[106] US Navy captain David Porter declared that, "after once tasting the Gallipagos tortoises, every other animal food fell off greatly in our estimation ... The meat of this animal is the easiest of digestion, and a quantity of it, exceeding that of any other food, can be eaten without experiencing the slightest of inconvenience."[79] Darwin was less enthusiastic about tortoise meat, writing "the breast-plate roasted (as the Gauchos do "carne con cuero"), with the flesh on it, is very good; and the young tortoises make excellent soup; but otherwise the meat to my taste is indifferent."[107]
In the 17th century, pirates started to use the Galápagos Islands as a base for resupply, restocking on food, water and repairing vessels before attacking Spanish colonies on the South American mainland. However, the Galápagos tortoises did not struggle for survival at this point because the islands were distant to busy shipping routes and harboured few valuable natural resources. As such they remained unclaimed by any nation, uninhabited and uncharted. In comparison, the tortoises of the islands in the Indian Ocean were already facing extinction by the late 17th century.[108]
Between the 1790s and the 1860s, whaling ships and fur-sealers systematically collected tortoises in far greater numbers than the buccaneers preceding them.[109] They were used for food and many more were killed for high grade "turtle oil" from the late 19th century onward for lucrative sale to continental Ecuador.[110] A total of over 13,000 tortoises is recorded in the logs of whaling ships between 1831 and 1868, and an estimated 100,000 were taken before 1830.[104] Since it was easiest to collect tortoises around coastal zones, females were most vulnerable to depletion during the nesting season. The collection by whalers came to a halt eventually through a combination of the scarcity of tortoises that they had created and the competition from crude oil as a cheaper energy source.[111]
Population decline accelerated with the early settlement of the islands in the early 19th century leading to unregulated hunting for meat, habitat clearance for agriculture and the introduction of alien mammal species.[44] Feral pigs, dogs, cats and black rats have become predators of eggs and young tortoises, whilst goats, donkeys and cattle compete for grazing and trample nest sites. The extinction of the Floreana subspecies in the mid-19th century has been attributed to the combined pressures of hunting for the penal colony on the relatively small island, the conversion of the grazing highlands into land for farming and fruit plantations, and the introduction of feral mammals.[112]
Scientific collection expeditions took 661 tortoises between 1888 and 1930, and more than 120 tortoises have been taken by poachers since 1990. Threats continue today with the rapid expansion of the tourist industry and increasing size of human settlements on the islands.[113]
Modern conservation
The remaining subspecies of tortoise range in IUCN classification from extinct in the wild to vulnerable. Slow growth rate, late sexual maturity and island endemism make the tortoises particularly prone to extinction without the efforts of conservationists.[62] The Galápagos giant tortoise has become a flagship species for conservation efforts throughout the Galápagos.
 Tourists see tortoises at the Charles Darwin Research Station.
Tourists see tortoises at the Charles Darwin Research Station.- Legal protection
The Galápagos giant tortoise is now strictly protected and is listed on Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora.[37] The listing requires that trade in the taxon and its products are subject to strict regulation by ratifying states, and international trade for primarily commercial purposes is prohibited. In 1936 the Ecuadorian government listed the giant tortoise as a protected species. In 1959, it declared all uninhabited areas in the Galápagos to be a National Park[114] and established the Charles Darwin Foundation. In 1970, capturing or removing many species from the islands (including tortoises and their eggs) was banned.[115] To halt trade in the tortoises altogether, it became illegal to export the tortoises from Ecuador, captive or wild, continental or insular in provenance. The banning of their exportation resulted in automatic prohibition of importation to the United States under Public Law 91-135 (1969).[116] A 1971 Ecuadorian decree made it illegal to damage, remove, alter or disturb any organism, rock or other natural object in the National Park.[117]
- Captive breeding
Breeding and releasing programs began in 1965 and have successfully brought seven of the eight endangered subspecies up to less perilous population levels. Young tortoises are raised at several breeding centres across the islands to improve their survival during their vulnerable early development. Eggs are collected from threatened nesting sites, and the hatched young are given a head start by being kept in captivity for four to five years to reach a size with a much better chance of survival to adulthood before release onto their native ranges.[84][102]
The most significant population recovery was that of the Española Tortoise (hoodensis), which was saved from near-certain extinction. The population had been depleted to 3 males and 12 females that had been so widely dispersed that no mating in the wild had occurred.[118] They were brought to the Charles Darwin Research Station in 1971 for a captive breeding program.[119] In the following 33 years, these 15 tortoises gave rise to over 1,200 tortoises released onto their home island, which themselves have begun to reproduce naturally.[120][121]
- Island restoration
The Galápagos National Park Service systematically culls feral predators and competitors. Goat eradication on islands including Pinta was achieved by the technique of using "Judas" goats with electronic collars to locate the herds. Marksmen shot all the goats except the Judas, then returned weeks later to find the "Judas" and shoot the herd to which it had relocated, repeating until only the "Judas" remained, which was then killed.[122] Other measures have included dog eradication from San Cristóbal, and fencing off nests to protect them from feral pigs.[84]
Efforts are now underway to repopulate islands formerly inhabited by tortoises in order to restore their ecosystems (island restoration) to their pre-human condition. The tortoises are a keystone species, acting as ecosystem engineers[122] which help in plant seed dispersal, trampling down brush and thinning the understory of vegetation (allowing light to penetrate and germination to occur). Birds such as flycatchers perch on and fly around tortoises in order to hunt the insects they displace from the brush.[64] In May 2010, 39 sterilised tortoises of hybrid origin were introduced to Pinta Island, the first tortoises there since the evacuation of Lonesome George 38 years ago in 1972.[123] Sterile tortoises were released so that the problem of interbreeding between subspecies would be avoided if any fertile tortoises were to be released in the future. It is hoped that with the recent identification of a hybrid abingdoni tortoise, the approximate genetic constitution of the original inhabitants of Pinta may eventually be restored with the identification and relocation of appropriate specimens to this island.[60] This approach may be used to "re-tortoise" Floreana in the future, since captive individuals have been found to be descended from the extinct original stock.[57]
Notes and references
- Notes
- ^ The first navigation chart showing the individual islands was done by the pirate Ambrose Cowley in 1684. He named them after fellow pirates or English noblemen. More recently, the Ecuadorian government gave most of the islands Spanish names. While the Spanish names are official, many researchers continue to use the older English names, particularly as those were the names used when Darwin visited. This article uses the Spanish island names.
- Citations
- ^ a b c d Rhodin, A.G.J.; van Dijk, P.P. (2010). "Turtles of the world, 2010 update: Annotated checklist of taxonomy, synonymy, distribution and conservation status". In Iverson, J.B.; Shaffer, H.B. Turtle taxonomy working group. Chelonian Research Foundation. pp. 33–34. doi:10.3854/crm.5.000.checklist.v3.2010. http://www.iucn-tftsg.org/wp-content/uploads/file/Accounts/crm_5_000_checklist_v3_2010.pdf.
- ^ a b c "Chelonoidis nigra". IUCN Red List of Threatened Species. Version 2009.2. International Union for Conservation of Nature. 1996. http://www.iucnredlist.org/apps/redlist/details/9011. Retrieved 2010-09-04.
- ^ a b c d e f Quoy, J.R.C.; Gaimard, J.P. (1824b). "Sous-genre tortue de terre – testudo. brongn. tortue noire – testudo nigra. N.". In de Freycinet, M.L. (in French). Voyage autour du Monde...exécuté sur les corvettes de L. M. "L'Uranie" et "La Physicienne," pendant les années 1817, 1818, 1819 et 1820. Paris. pp. 174–175.
- ^ a b Gunther 1877, p. 85
- ^ a b c Rothschild, Walter (1901). "On a new land-tortoise from the Galapagos Islands". Novitates Zoologicae 8: 372. http://www.biodiversitylibrary.org/page/3268176. Retrieved 14 April 2011.
- ^ a b c d e f g h i Van Denburgh, John (1907). "Preliminary descriptions of four new races of gigantic land tortoises from the Galapagos Islands". Proceedings of the California Academy of Sciences, Series 4 1: 1–6. http://biodiversitylibrary.org/page/31599075. Retrieved 14 April 2011.
- ^ a b c d e f Garman, Samuel (1917). "The Galapagos tortoises". Memoirs of the Museum of Comparative Zoology at Harvard College 30 (4): 261–296.
- ^ a b c Rothschild, Walter (1903). "Description of a new species of gigantic land tortoise from Indefatigable Island". Novitates Zoologicae 10: 119. http://www.biodiversitylibrary.org/page/15771201. Retrieved 14 April 2011.
- ^ a b c d e f Günther, Albert (1875). "Description of the living and extinct races of gigantic land-tortoises. parts I. and II. introduction, and the tortoises of the Galapagos Islands". Philosophical Transactions of the Royal Society of London, Biological Sciences 165: 251–284. Bibcode 1875RSPT..165..251G. doi:10.1098/rstl.1875.0007. JSTOR 109147.
- ^ a b c Pritchard 1996, p. 49
- ^ a b c Quoy, J.R.C.; Gaimard, J.P. (1824). "Description d'une nouvelle espèce de tortue et de trois espèces nouvelles de scinques" (in French). Bulletin des Sciences Naturelles et de Géologie (Paris) 1: 90–91.
- ^ a b Harlan, Richard (1827). "Description of a land tortoise, from the Galápagos Islands, commonly known as the "Elephant Tortoise"". Journal of the Academy of Natural Sciences of Philadelphia 5: 284–292. http://biodiversitylibrary.org/page/24657342. Retrieved 14 April 2011.
- ^ a b c Duméril, André Marie Constant; Bibron, Gabriel (1834) (in French), Erpétologie générale; ou, histoire naturelle complète des reptiles, France: Librarie Encyclopédique de Roret, http://www.archive.org/details/erptologiegnral00bibrgoog
- ^ Gray, John Edward (1853). "Description of a new species of tortoise (Testudo planiceps), from the Galapagos Islands". Proceedings of the Zoological Society of London 21 (1): 12–13. doi:10.1111/j.1469-7998.1853.tb07165.x. http://biodiversitylibrary.org/page/30747129. Retrieved 14 April 2011.
- ^ Williams, Ernest E.; Anthony, Harold Elmer; Goodwin, George Gilbert (1952). "A new fossil tortoise from Mona Island West Indies and a tentative arrangement of the tortoises of the world". Bulletin of the American Museum of Natural History 99 (9): 541–560. hdl:2246/418.
- ^ Pritchard, Peter Charles Howard (1967). Living turtles of the world. New Jersey: TFH Publications. p. 156.
- ^ Bour, R. (1980). "Essai sur la taxinomie des Testudinidae actuels (Reptilia, Chelonii)" (in French). Bulletin du Museum D'Histoire Naturelle, Paris (Paris) 4 (2): 546.
- ^ a b c d Baur, G. (1889). "The gigantic land tortoises of the Galapagos Islands" (PDF). The American Naturalist 23 (276): 1039–1057. doi:10.1086/275045. http://www.iucn-tftsg.org/wp-content/uploads/file/Articles/Baur_1889b.pdf.
- ^ a b c Rothschild, Walter (1902). "Description of a new species of gigantic land tortoise from the Galápagos Islands". Novitates Zoologicae 9: 619. http://biodiversitylibrary.org/page/3269130. Retrieved 14 April 2011.
- ^ a b DeSola, Ralph (1930). "The liebespiel of Testudo vandenburghi, a new name for the mid-Albemarle Island Galápagos tortoise". Copeia 1930 (3): 79–80. doi:10.2307/1437060. JSTOR 1437060?.
- ^ Stewart, P.D. (2006). Galápagos: the islands that changed the world. New Haven: Yale University Press. p. 43. ISBN 978-0-300-12230-5.
- ^ Pritchard 1996, p. 17
- ^ Jackson, Michael Hume (1993). Galápagos, a natural history. Calgary: University of Calgary Press. p. 1. ISBN 1-895176-07-6.
- ^ Chambers 2004, p. 27
- ^ Perrault, Claude (1676) (in French), Suite des memoires pour servir a l'histoire naturelle des animaux, Paris: Académie de Sciences, pp. 92–205
- ^ Schneider, Johann Gottlob (1783) (in German). Allgemeine Naturgeschischte der Schildkröten nebs einem systematischen Verzeichnisse der einzelnen Arten und zwey Kupfen. Leipzig. pp. 355–356. http://books.google.com/books?id=wl5HAAAAYAAJ&pg=PA355.
- ^ Schweigger, August Friedrich (1812). "Prodromi monographiae chelonorum sectio prima" (in Latin). Archivfur Naturwissenschaft und Mathematik (Köningsberg) 1: 271–368; 406–462.
- ^ Gunther 1877, p. 9
- ^ a b c d e f Caccone, Adalgisa; Gibbs, James P. ; Ketmaier, Valerio; Suatoni, Elizabeth; Powell, Jeffrey R. (1999). "Origin and evolutionary relationships of giant Galapagos tortoises". Proceedings of the National Academy of Science 96 (23): 13223–13228. doi:10.1073/pnas.96.23.13223. http://www.pnas.org/content/96/23/13223.full.
- ^ Austin, Jeremy; Arnold, E. Nicholas (2001). "Ancient mitochondrial DNA and morphology elucidate an extinct island radiation of Indian Ocean giant tortoises (Cylindraspis)". Proceedings of the Royal Society B: Biological Sciences 268 (1485): 2515–2523. doi:10.1098/rspb.2001.1825. http://rspb.royalsocietypublishing.org/content/268/1485/2515.full.pdf+html.
- ^ Austin, Jeremy; Arnold, E. Nicholas; Bour, Roger (2003). "Was there a second adaptive radiation of giant tortoises in the Indian Ocean? Using mitochondrial DNA to investigate speciation and biogeography of Aldabrachelys" (PDF). Molecular Ecology 12 (6): 1415–1424. doi:10.1046/j.1365-294X.2003.01842.x. PMID 12755871.
- ^ a b c d e f g h Van Denburgh, J. (1914). "The gigantic land tortoises of the Galapagos archipelago". Proceedings of the California Academy of Sciences, Series 4 2 (1): 203–374. http://biodiversitylibrary.org/page/31626507. Retrieved 14 April 2011.
- ^ Pritchard, Peter Charles Howard (1984). "Further thoughts on Lonesome George". Noticias de Galápagos 39: 20–23.
- ^ Pritchard, Peter Charles Howard (1997). "Galapagos tortoise nomenclature: a reply". Chelonian Conservation and Biology (University of California) 2: 619.
- ^ Le, Minh; Raxworthy, Christopher J.; McCord, William P.; Mertz, Lisa (2006). "A molecular phylogeny of tortoises (Testudines:Testudinidae) based on mitochondrial and nuclear genes". Molecular Phylogenetics and Evolution 40 (2): 517–531. doi:10.1016/j.ympev.2006.03.003. PMID 16678445.
- ^ Fritz, U.; Havaš, P. (2007). "Checklist of chelonians of the world". Vertebrate Zoology 57 (2): 149–368. http://www.vertebrate-zoology.de/vz57-2/57-2_Fritz_149-368.pdf.
- ^ a b "CITES Appendices I, II and III 2010". http://www.cites.org/eng/app/appendices.shtml.
- ^ a b c Caccone, Adalgisa; Gentile, Gabriele; Gibbs, James P.; Fritts, Thomas H.; Snell, Howard L.; Betts, Jessica; R. Powell, Jeffrey (2002). "Phylogeography and history of the giant Galapagos tortoises". Evolution 56 (10): 2052–2066. JSTOR 3094648. PMID 12449492.
- ^ Broom, R. (1929). "On the extinct Galápagos tortoise that inhabited Charles Island". Zoologica 9 (8): 313–320.
- ^ a b Steadman, David W. (1986). "Holocene vertebrate fossils from Isla Floreana, Galápagos". Smithsonian Contributions to Zoology 413: 1–103. http://si-pddr.si.edu/dspace/bitstream/10088/5333/2/SCtZ-0413-Lo_res.pdf.
- ^ a b Sulloway, F.J. (1984). Darwin and the Galápagos. In Berry, R.J. "Evolution in the Galápagos Islands". Biological Journal of the Linnean Society 21 (1–2): 29–60. doi:10.1111/j.1095-8312.1984.tb02052.x.
- ^ Pritchard 1996, p. 63
- ^ a b c d e f g h i j k Swingland, I.R. (1989). Geochelone elephantopus. Galapagos giant tortoises. In: Swingland I.R. and Klemens M.W. (eds.) The conservation biology of tortoises. Occasional Papers of the IUCN Species Survival Commission (SSC), No. 5, pp. 24–28. Gland, Switzerland: IUCN. ISBN 2-88032-986-8.
- ^ a b c d e MacFarland, Craig G.; Villa, José; Toro, Basilio (1974). "The Galápagos giant tortoises (Geochelone elephantopus). I. Status of the surviving populations". Biological Conservation 6 (2): 118–133. doi:10.1016/0006-3207(74)90024-X.
- ^ MacFarland, Craig G.; Villa, José; Toro, Basilio (1974b). "The Galápagos giant tortoises (Geochelone elephantopus). Part II: Conservation methods". Biological Conservation 6 (3): 198–212. doi:10.1016/0006-3207(74)90068-8.
- ^ a b Márquez, Cruz; Fritts, Thomas H.; Koster, Friedmann; Rea, Solanda; Cepeda, Fausto; Llerena, y Fausto (1995), "Comportamiento de apareamiento al azar en tortugas gigantes. Juveniles en cautiverio el las Islas Galápagos" (in Spanish), Revista de Ecología Latinoamericana 3: 13–18, http://www.ciens.ula.ve/~cires/recol-v3n1a03.pdf
- ^ a b c d e f Fritts, Thomas H. (1984). "Evolutionary divergence of giant tortoises in Galapagos". Biological Journal of the Linnean Society 21 (1–2): 165–176. doi:10.1111/j.1095-8312.1984.tb02059.x.
- ^ a b c d e f g h i De Vries, T.J. (1984). The giant tortoises: a natural history disturbed by man. Oxford: Pergamon Press. pp. 145–156. ISBN 0-08-027996-1.
- ^ Ernst, Carl H.; Barbour, Roger (1989). Turtles of the world. Washington, D.C.: Smithsonian Institution Press. ISBN 0-87474-414-8. http://nlbif.eti.uva.nl/bis/turtles.php?selected=beschrijving&menuentry=soorten&record=Geochelone.
- ^ a b Poulakakis, Nikos; Glaberman, Scott; Russello, Michael; Beheregaray, Luciano B.; Ciofi, Claudio; Powell, Jeffrey R.; Caccone, Adalgisa (2008). "Historical DNA analysis reveals living descendants of an extinct species of Galapagos tortoise". Proceedings of the National Academy of Sciences 105 (40): 15464–15469. doi:10.1073/pnas.0805340105. http://www.pnas.org/content/105/40/15464.full.
- ^ Zug, George R. (1997). "Galapagos tortoise nomenclature: still unresolved". The Journal of Chelonian Conservation and Biology 2 (4): 618–619. hdl:10088/11695.
- ^ White, William M.; McBirney, Alexander R.; Duncan, Robert A. (1993). "Petrology and geochemistry of the Galápagos Islands: portrait of a pathological mantle plume". Journal of Geophysical Research 98 (B11): 19533–19564. Bibcode 1993JGR....9819533W. doi:10.1029/93JB02018.
- ^ Nicholls 2006, p. 68
- ^ Beheregaray, Luciano B.; Gibbs, James P.; Havill, Nathan; Fritts, Thomas H.; Powell, Jeffrey R.; Caccone, Adalgisa (2004). "Giant tortoises are not so slow: rapid diversification and biogeographic consensus in the Galápagos". Proceedings of the National Academy of Sciences 101 (17): 6514–6519. doi:10.1073/pnas.0400393101. http://www.pnas.org/content/101/17/6514.full.
- ^ a b c Ciofi, Claudio; Wilson, Gregory A.; Beheregaray, Luciano B.; Marquez, Cruz; Gibbs, James P.; Tapia, Washington; Snell, Howard L.; Caccone, Adalgisa; Powell, Jeffrey R. (2006). "Phylogeographic history and gene flow among giant Galápagos tortoises on southern Isabela Island". Genetics 172 (3): 1727–1744. doi:10.1534/genetics.105.047860. PMC 1456292. PMID 16387883. http://www.genetics.org/cgi/content/full/172/3/1727.
- ^ a b c d Pritchard 1996, p. 65
- ^ a b Russello, Michael A.; Poulakakis, Nikos; Gibbs, James P.; Tapia, Washington; Benavides, Edgar; Powell, Jeffrey R.; Caccone, Adalgisa (2010). Knapp, Michael. ed. "DNA from the past informs ex situ conservation for the future: an extinct species of Galapagos tortoise identified in captivity". PLoS One 5 (1): e8683. doi:10.1371/journal.pone.0008683. PMC 2800188. PMID 20084268. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2800188. Retrieved 2011-03-15.
- ^ Pak, Hasong; Zaneveld, J.R.V. (1973). "The Cromwell Current on the east side of the Galapagos Islands". Journal of Geophysical Research 78 (33): 7845–7859. Bibcode 1973JGR....78.7845P. doi:10.1029/JC078i033p07845. http://www.agu.org/pubs/crossref/1973/JC078i033p07845.shtml. Retrieved 2011-03-15.
- ^ Nicholls 2006, p. 161
- ^ a b Russello, Michael A.; Beheregaray, Luciano B.; Gibbs, James P.; Fritts, Thomas; Havill, Nathan; Powell, Jeffrey R.; Caccone, Adalgisa (2007). "Lonesome George is not alone among Galapagos tortoises". Current Biology 17 (9): R317–R318. doi:10.1016/j.cub.2007.03.002. PMID 17470342.
- ^ Russello, Michael A.; Glaberman, Scott; Gibbs, James P.; Marquez, Cruz; Powell, Jeffrey R.; Caccone, Adalgisa (2005). "A cryptic taxon of Galapagos tortoise in conservation peril". Biological Letters 1 (3): 287–290. doi:10.1098/rsbl.2005.0317. http://rsbl.royalsocietypublishing.org/content/1/3/287.full.
- ^ a b c Chiari, Ylenia; Hyseni, Chaz; Fritts, Tom H.; Glaberman, Scott; Marquez, Cruz; Gibbs, James P.; Claude, Julien; Caccone, Adalgisa (2009). Gemmell, Neil John. ed. "Morphometrics parallel genetics in a newly discovered and endangered taxon of Galapagos tortoise". PLoS One 4 (7): e6272. doi:10.1371/journal.pone.0006272. PMC 2707613. PMID 19609441. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2707613.
- ^ Ciofi, Claudio; Milinkovitch, Michel C.; Gibbs, James P.; Caccone, Adalgisa; Powell, Jeffrey R. (2002). "Microsatellite analysis of genetic divergence among populations of giant Galápagos tortoises". Molecular Ecology 11 (11): 2265–2283. doi:10.1046/j.1365-294X.2002.01617.x. PMID 12406238.
- ^ a b c d MacFarland, C.G. (1972). "Giant tortoises, goliaths of the Galapagos". National Geographic Magazine 142: 632–649.
- ^ de Berlanga, Tomás (1535) (in Spanish). Letter to His Majesty ... describing his voyage from Panamá to Puerto Viejo. 41. Madrid: Manuel G. Hernandez. pp. 538–544.
- ^ a b c d e f Darwin 1839, pp. 462–466
- ^ Ebersbach, V.K. (2001) (in German). Zur Biologie und Haltung der Aldabra-Riesenschildkröte (Geochelone gigantea) und der Galapagos-Riesenschildkröte (Geochelone elephantopus) in menschlicher Obhut unter besonderer Berücksichtigung der Fortpflanzun (PhD thesis). Hannover: Tierärztliche Hochschule. http://elib.tiho-hannover.de/dissertations/ebersbachk_2001.pdf.
- ^ a b c Fritts, T.H. (1983). "Morphometrics of Galapagos tortoises: evolutionary implications". In Bowman, I.R.; Berson, M.; Leviton, A.E. Patterns of evolution in Galapagos organisms. San Francisco: American Association for the Advancement of Science. pp. 107–122. ISBN 0-934394-05-9.
- ^ Pritchard 1996, p. 18
- ^ Auffenberg, Walter (1971). "A new fossil tortoise, with remarks on the origin of South American testudinines". Copeia 1971 (1): 106. JSTOR 1441604. (subscription required)
- ^ Dawson, E.Y. (1966). "Cacti in the Galápagos islands, with special reference to their relations with tortoises". In Bowman, R.I. The Galápagos. Berkeley: University of California Press. pp. 209–214. http://books.google.com/books?id=f8T3Dl5tokYC&pg=PA209.
- ^ de Neira, Lynn E. Fowler; Roe, John H. (1984). "Emergence success of tortoise nests and the effect of feral burros on nest success on Volcán Alcedo, Galapagos". Copeia 1984 (3): 702–707. doi:10.2307/1445152. JSTOR 1445152.
- ^ a b c Schafer, Susan F.; Krekorian, C. O'Neill (1983). "Agonistic behavior of the Galapagos tortoise, Geochelone elephantopus, with emphasis on its relationship to saddle-backed shell shape". Herpetologica 39 (4): 448–456. JSTOR 3892541.
- ^ Marlow, Ronald William; Patton, James L. (1981). "Biochemical relationships of the Galápagos giant tortoises (Geochelone elephantopus)". Journal of Zoology 195 (3): 413–422. doi:10.1111/j.1469-7998.1981.tb03474.x.
- ^ a b Carpenter, Charles C. (1966). "Notes on the behavior and ecology of the Galapagos tortoise on Santa Cruz island". Proceedings of the Oklahoma Academy of Science 46: 28–32.
- ^ Jackson, M.H. (1993). Galápagos, a natural history. University of Calgary Press. p. 107. ISBN 1-895176-07-6.
- ^ Cayot, Linda Jean (1987). Ecology of giant tortoises (Geochelone elephantopus) in the Galápagos Islands (Ph.D. thesis). Syracuse: Syracuse University.
- ^ Hatt, Jean-Michel; Class, Marcus; Gisler, Ricarda; Liesegang, Annette; Wanner, Marcel (2005). "Fiber digestibility in juvenile Galapagos tortoises (Geochelone nigra) and implications for the development of captive animals". Zoo Biology 24 (2): 185–191. doi:10.1002/zoo.20039.
- ^ a b c Porter, David (1815). Journal of the cruise made to the Pacific Ocean by Captain Porter in the United States Frigate Essex in the years 1812, 1813, 1814. New York: Wiley & Halsted. p. 151. http://books.google.com/books?id=IiYFAAAAMAAJ&pg=PA151.
- ^ MacFarland, Craig G.; Reeder, W.G. (1974). "Cleaning symbiosis involving Galápagos tortoises and two species of Darwin's finches". Zeitschrift für Tierpsychologie 34 (5): 464–483. doi:10.1111/j.1439-0310.1974.tb01816.x.
- ^ Bonin, Franck; Devaux, Bernard; Dupré, Alain (2006). Turtles of the world. Pritchard, Peter Charles Howard. Baltimore: Johns Hopkins University Press. ISBN 0-8018-8496-9.
- ^ Hayes, Floyd E.; Beaman, Kent R. Beaman; Hayes, William K.; Harris, Lester E. Jr. (1988). "Defensive behavior in the Galapagos tortoise (Geochelone elephantopus), with comments on the evolution of insular gigantism". Herpetologica 44 (1): 11–17. JSTOR 3892193.
- ^ Ciofi, Claudio; Swingland, Ian R. (1995). "Environmental sex determination in reptiles". Applied Animal Behaviour Science 51 (3): 251–265. http://www.seaturtle.org/PDF/Ciofi_1997_AppAnimBehavSci.pdf.
- ^ a b c Cayot, L.J. (1994). Captive management and conservation of amphibians and reptiles. 11. Ithaca: Society for the Study of Amphibians and Reptiles. 297–305. ISBN 0-916984-33-8.
- ^ "Galápagos tortoise Geochelone elephantophus". National Geographic Society. http://animals.nationalgeographic.com/animals/reptiles/galapagos-tortoise/. Retrieved 2010-09-04.
- ^ a b "Giant tortoises of the Galápagos". American Museum of Natural History. http://www.amnh.org/exhibitions/darwin/cam/about.php. Retrieved 2010-09-04.
- ^ Chambers, Paul (2004). "The origin of Harriet". New Scientist (2464). http://www.newscientist.com/article/mg18324645.000-the-origin-of-harriet.html?full=true. (subscription required)
- ^ "Harriet the tortoise dies". ABC News Online. 2006. http://www.abc.net.au/news/newsitems/200606/s1670600.htm. Retrieved 2010-09-13.
- ^ Grant, K.T.; Estes, G.B. (2009), Darwin in Galapagos: footsteps to a new world, Princeton: Princeton University Press, ISBN 978-0-691-14210-4
- ^ a b Darwin, C.R. (1845). Journal of researches into the natural history and geology of the countries visited during the voyage of H.M.S. Beagle round the world, under the Command of Capt. Fitz Roy (2nd ed.). London: John Murray. pp. 393–394. http://darwin-online.org.uk/content/frameset?viewtype=side&itemID=F14&pageseq=406.
- ^ Chambers 2004, p. 51
- ^ Nicholls 2006, p. 62
- ^ a b Darwin 1839, p. 456
- ^ Keynes, R.D. (2001). Charles Darwin's Beagle diary. Cambridge: Cambridge University Press. p. 362. ISBN 0-521-00317-2. http://darwin-online.org.uk/content/frameset?viewtype=side&itemID=F1925&pageseq=394.
- ^ Chambers 2004, p. 41
- ^ Chambers 2004, p. 52
- ^ Gunther 1877, p. 62 for example
- ^ Darwin, C.R. (1827; 1828–1829; 1837–1839), Edinburgh notebook, CUL-DAR118, p. 99, http://darwin-online.org.uk/content/frameset?viewtype=side&itemID=CUL-DAR118.-&pageseq=41
- ^ Chambers 2004, p. 47
- ^ Barlow, N. (1963), "Darwin's ornithological notes", Bulletin of the British Museum (Natural History). Historical Series 2 (7): 262, http://darwin-online.org.uk/content/frameset?viewtype=side&itemID=F1577&pageseq=64
- ^ Darwin, C.R. (1837–1838), Notebook B: [Transmutation of species], p. 7, http://darwin-online.org.uk/content/frameset?viewtype=side&itemID=CUL-DAR121.-&pageseq=9
- ^ a b "Reproduction, breeding, repatriation, and monitoring of tortoises". Galapagos National Park. 2009-06-29. http://www.galapagospark.org/nophprg.php?page=parque_nacional_nativas_endemicas_tortugas. Retrieved 2010-09-04.
- ^ Footman, T. (2000). Guinness World Records 2001. Guinness Records. p. 182. ISBN 0-85112-102-0.
- ^ a b Townsend, Charles Haskins (1925). "The Galapagos tortoises in their relation to the whaling industry: a study of old logbooks". Zoologica 4: 55–135. http://mysite.du.edu/~ttyler/ploughboy/townsendgaltort.htm.
- ^ Dampier, William (2007) [1697]. "Chapter 5". A new voyage around the world. Dampier: Dover Publications. ISBN 978-1-933698-04-5. http://www.galapagos.to/TEXTS/DAMPIER-1.HTM#galapagos.
- ^ Colnett, J. (1968) [1798]. A voyage to the south Atlantic. USA: Da Capo Press. p. 158. http://www.archive.org/stream/cihm_33242#page/n195/mode/2up.
- ^ Darwin 1839, p. 459
- ^ Chambers 2004, p. 85
- ^ Chambers 2004, p. 94
- ^ Beck, RH. (1903). "In the home of the giant tortoise". 7th Annual Report of the New York Zoological Society (New York): 1–17.
- ^ Chambers 2004, p. 104
- ^ Hoeck, H.N. (1984). "Introduced fauna". In Perry, R. Key environments. Galápagos. Pergamon Press. pp. 233–245. ISBN 0-08-027996-1.
- ^ Watkins, Graham; Cruz, Felipe (2007). "Galapagos at risk: a socioeconomic analysis". Charles Darwin Foundation. http://www.darwinfoundation.org/english/_upload/Galapagos_At-Risk-2007.pdf.
- ^ Ecuador (1959). (In Spanish). Decreto ley de emergencia, por el cual se declaran parques nacionales de reserva de exclusivo dominio del estado, para la preservación de la fauna y flora, todas las tierras que forman las islas del Archipiélago de Colón o Galápagos, Registro Oficial No. 873 (1959) Quito, 20 de Julio, 1959
- ^ Ecuador (1970). (In Spanish). Ley de Protección de la Fauna Silvestre y de los Recursos Ictiológicos, Registro Oficial No. 104 (1970) Quito, 20 de Noviembre 1970
- ^ United States 1969. Endangered Species Conservation Act. 83 Stat. 275; 16 USC 668 cc-1 to 668 cc-6 (1969)
- ^ Ecuador (1971). (In Spanish). Decreto Supremo No. 1306 (1971) Quito, 27 de Agosto 1971
- ^ Corley Smith, G.T. (1977). "The present status of the giant tortoise Geochelone elephantopus on the Galapagos islands". International Zoo Yearbook 17 (1): 109–112. doi:10.1111/j.1748-1090.1977.tb00877.x.
- ^ Caporaso, Fred (1991). "The Galápagos tortoise conservation program: the plight and future for the Pinzón Island tortoise". Proceedings of the first international symposium on turtles & tortoises: conservation and captive husbandry. Chapman University. pp. 113–116. http://www.tortoise.org/archives/pinzon.html.
- ^ Pritchard 1996, p. 59
- ^ Milinkovitch, Michel C.; Monteyne, Daniel; Gibbs, James P.; Fritts, Thomas H.; Tapia, Washington; Snell, Howard L.; Tiedemann, Ralph; Caccone, Adalgisa; Powell, Jeffrey R. (2004). "Genetic analysis of a successful repatriation programme: giant Galapagos tortoises". Proceedings of the Royal Society of London B 271 (1537): 341–345. doi:10.1098/rspb.2003.2607. http://rspb.royalsocietypublishing.org/content/271/1537/341.full.pdf.
- ^ a b Carroll, R. (2010-06-27). "Galápagos giant tortoise saved from extinction by breeding programme". The Observer. http://www.guardian.co.uk/environment/2010/jun/27/giant-tortoise-galapagos-saved-extinction. Retrieved 2010-09-01.
- ^ "Project Pinta". Galapagos Conservancy. http://www.galapagos.org/2008/index.php?id=68. Retrieved 2010-09-04.
- Bibliography
- Chambers, Paul (2004). A sheltered life: the unexpected history of the giant tortoise. London: John Murray. ISBN 0-7195-6528-6.
- Darwin, Charles (2009) [1839]. Narrative of the surveying voyages of His Majesty's Ships Adventure and Beagle between the years 1826 and 1836, describing their examination of the southern shores of South America, and the Beagle's circumnavigation of the globe. Journal and remarks. (facsimile ed.). General Books LLC. ISBN 978-1-151-09182-6.
- Günther, Albert Carl Ludwig Gotthilf (2010) [1877]. The gigantic land-tortoises (living and extinct) in the collection of the British Museum (facsimile reprint ed.). Nabu Press. ISBN 978-1-141-78978-8. http://books.google.com/books?id=DO8ZAAAAYAAJ&pg=PP1.
- Nicholls, Henry (2006). Lonesome George: The life and loves of a conservation icon. Hampshire, England: Palgrave Macmillan. ISBN 1-4039-4576-4.
- Pritchard, Peter Charles Howard (1996). The Galapagos tortoises: nomenclatural and survival status. Chelonian Research Foundation. ISBN 0-9653540-0-8.
External links
- ARKive – images and movies of the Galápagos giant tortoise (Geochelone spp.)
- American Museum of Natural History, Galápagos Giant Tortoise webpage
Tortoise family of turtles (Testudinidae) Genus Species of the Tortoise family
Aldabrachelys Astrochelys Angonoka tortoise · Radiated tortoiseChelonoidis Argentine tortoise · Brazilian giant tortoise · Chaco tortoise · Galápagos tortoise · Red-footed tortoiseChersina Cylindraspis† Geochelone Gopherus Homopus Beaked cape tortoise · Berger's cape tortoise · Boulenger's cape tortoise · Karoo cape tortoise · Speckled padloper tortoiseIndotestudo Elongated tortoise · Forsten's tortoise · Travancore tortoiseKinixys Bell's hinge-back tortoise · Forest hinge-back tortoise · Home's hinge-back tortoise · Lobatse hinge-back tortoise · Natal hinge-back tortoise · Speke's hinge-back tortoiseMalacochersus Manouria Psammobates Pyxis Stigmochelys Testudo Phylogenetic arrangement of turtles based on turtles of the world 2010 update: annotated checklist of taxonomy, synonymy, distribution and conservation status. Key: †=extinct. 1=classification unclearOrder Testudines (turtles) Suborder SuperfamilySubfamily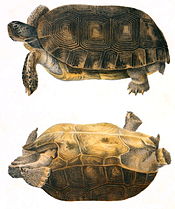
Cryptodira Chelydra · MacrochelysCaretta · Chelonia · Eretmochelys · Lepidochelys · NatatorDermochelysDermatemydidaeDermatemysStaurotypinaeClaudius · StaurotypusBatagur · Cuora · Cyclemys · Geoclemys · Geoemyda · Hardella · Heosemys · Leucocephalon · Malayemys · Mauremys · Melanochelys · Morenia · Notochelys · Orlitia · Pangshura · Rhinoclemmys · Sacalia · Siebenrockiella · VijayachelysAldabrachelys · Astrochelys · Chelonoidis · Chersina · Cylindraspis · Geochelone · Gopherus · Homopus · Indotestudo · Kinixys · Malacochersus · Manouria · Psammobates · Pyxis · Stigmochelys · TestudoTrionychiaCarettochelyidaeCarettochelysTrionychinaePleurodira ChelidinaeChelodininaeHydromedusinaePelomedusa · PelusiosPhylogenetic arrangement based on turtles of the world 2010 update: annotated checklist. Extinct turtles not included.
See also List of Testudines families
 Portal ·
Portal ·  WikiProjectCategories:
WikiProjectCategories:- IUCN Red List vulnerable species
- Chelonoidis
- Endemic fauna of the Galápagos Islands
- Megafauna
- C. n. abingdoni (Extinct in the wild)















Wikimedia Foundation. 2010.