- P-type ATPase
-
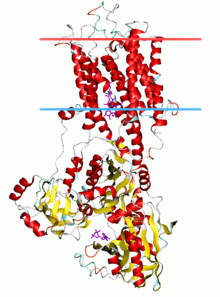
Calcium ATPase, E2-Pi state Identifiers Symbol E1-E2_ATPase Pfam PF00122 InterPro IPR008250 PROSITE PDOC00139 SCOP 1eul TCDB 3.A.3 OPM family 22 OPM protein 3b9b Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary Cation transporting ATPase, C-terminus 
crystal structure of the sr ca2+-atpase with bound amppcp Identifiers Symbol Cation_ATPase_C Pfam PF00689 InterPro IPR006068 SCOP 1eul TCDB 3.A.3 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary Cation transporter/ATPase, N-terminus 
model of neurospora crassa proton atpase Identifiers Symbol Cation_ATPase_N Pfam PF00690 InterPro IPR004014 SCOP 1eul TCDB 3.A.3 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary The P-type ATPases, also known as E1-E2 ATPases, are a large group of evolutionarily related ion pumps that are found in bacteria, archaea and eukaryotes. They are α-helical bundle primary transporters referred to as P-type ATPases because they catalyze auto- (or self-) phosphorylation of a key conserved aspartate residue within the pump. In addition, they all appear to interconvert between at least two different conformations, denoted by E1 and E2.
Most members of this transporter family are specific for the pumping of a large array of cations, however one subfamily is involved in flipping phospholipids to maintain the asymmetric nature of the biomembrane.
Prominent examples of P-type ATPases are the sodium-potassium pump (Na+,K+-ATPase), the proton pump (H+-ATPase), the proton-potassium pump (H+,K+-ATPase) and the calcium pump (Ca2+-ATPase).
Contents
Discovery
The first P-type ATPase discovered was the Na+,K+-ATPase, which Nobel laureate Jens Christian Skou isolated in 1957[1]. The Na+,K+-ATPase was only the first member of a large and still growing protein family, which in June 2007 had around 400 confirmed and unique members in Swiss-Prot (Prosite motif PS00154).
Phylogenetic classification
A phylogenetic analysis of 159 sequences made in 1998 by Axelsen and Palmgren showed that P-type ATPases can be divided into five subfamilies, based strictly on a conserved sequence kernel excluding the highly variable N and C terminal regions. The phylogenetic analysis grouped the proteins independent of the organism from which they are isolated and showed that the diversification of the P-type ATPase family occurred prior to the separation of eubacteria, archaea, and eucaryota. This underlines the significance of this protein family for cell survival.[2]
- Type I consists of the transition/heavy metal ATPases.
- Type IA ATPases are involved in K+ import. They are atypical P-type ATPases because, unlike other P-type ATPases, they function as part of a heterotetrameric complex (called KdpFABC), where the actual K+ transport is mediated by another subcomponent of the complex.
- Type IB ATPases are involved in transport of the soft Lewis acids: Cu+, Ag+, Cu2+, Zn2+, Cd2+, Pb2+ and Co2+. They are key elements for metal resistance and metal homeostasis in a wide range of organisms.
- Type II ATPases are split into four groups.
- Type IIA transports Ca2+. SERCA1a is a type IIA pump.
- Type IIB transports Ca2+.
- Type IIC consists of the closely related Na+/K+ and H+/K+ ATPases from animal cells.
- Type IID contains a small number of fungal ATPases of unknown function.
- Type III ATPases contains the plasma membrane H+-ATPases from plants and fungi (IIIA) and a small subdivision with Mg2+-ATPases from three bacterial species (IIIB).
- Type IV ATPases have been shown to be involved in the transport of phospholipids.[3] However the transport specificity of the P-IV type ATPases still remains a somewhat controversial subject.
- Type V ATPases have unknown specificity. This large group are only found in eukaryotes and are believed to be involved in cation transport in the endoplasmic reticulum.
Human genes
Human genes encoding P-type ATPases or P-type ATPase-like proteins include:
- Na+/K+ transporting: ATP1A1, ATP1A2, ATP1A3, ATP1A4, ATP1B1, ATP1B2, ATP1B3, ATP1B4
- Ca++ transporting: ATP2A1, ATP2A2, ATP2A3, ATP2B1, ATP2B2, ATP2B3, ATP2B4, ATP2C1
- Cu++ transporting: ATP7A, ATP7B
- Class I, type 8: ATP8A1, ATP8B1, ATP8B2, ATP8B3, ATP8B4
- Class II, type 9: ATP9A, ATP9B
- Class V, type 10: ATP10A, ATP10B, ATP10D
- Class VI, type 11: ATP11A, ATP11B, ATP11C
- H+/K+ transporting, nongastric: ATP12A
- type 13: ATP13A1, ATP13A2, ATP13A3, ATP13A4, ATP13A5
- ATP2C2;
- ATP4A;
- KIAA0195
Evolutionary origin
One of the open reading frames in the genome of the archaea Methanococcus jannaschii deserves special attention, because it might reveal information about the evolution of the ancestor P-type ATPase. This open reading frame covers the big cytoplasmic loop common to all P-type ATPases, which contain all the amino acids involved in ATP binding and hydrolysis. It is speculated that this soluble protein has fused with an ion channel, thus creating the first P-type ATPase.[4]
Structure
Most of our knowledge about the structure and function of P-type ATPases originates from SERCA1a, a sarco(endo)plasmic reticulum Ca2+-ATPase of fast twitch muscle from adult rabbit. It is generally acknowledged that the structure of SERCA1a is representative for the family of P-type ATPases.[5]
Structures of the Na+/K+-ATPase and H+-ATPase are also available.
Structure of SERCA1a
SERCA1a is composed of a cytoplasmic section and a transmembrane section with two Ca2+ binding sites. The cytoplasmic section consists of three cytoplasmic domains, designated the P, N and A domains, containing over half the mass of the protein. The transmembrane section has ten transmembrane helices (M1-M10), with the two Ca2+ binding sites located near the midpoint of the bilayer. The binding sites are formed by side-chains and backbone carbonyls from M4, M5, M6, and M8. M4 is unwound in this region due to a conserved proline (P308). This unwinding of M4 is recognised as a key structural feature of P-type ATPases.
The P domain contains the canonical aspartic acid phosphorylated during the reaction cycle. It is composed of two parts widely separated in sequence. These two parts assemble into a seven stranded parallel ß-sheet with eight short associated a-helices, forming a Rossmann fold.
The N domain is inserted between the two segments of the P domain, and is formed of a seven strand antiparallel ß-sheet between two helix bundles. This domain contains the ATP-binding pocket, pointing out toward the solvent near the P-domain.
The A domain is the smallest of the three domains. It consists of a distorted jellyroll structure and two short helices. It is the actuator domain modulating the occlusion of Ca2+ in the transmembrane binding sites, and it is pivot in transposing the energy from the hydrolysis of ATP in the cytoplasmic domains to the vectorial transport of cations in the transmembrane domain. The A domain dephosphorylates the P domain as part of the reaction cycle using a highly conserved TGES motif located at one end of the jellyroll.
ATP hydrolysis occurs in the cytoplasmic headpiece at the interface between domain N and P. Two Mg-ion sites forms part of the active site. ATP hydrolysis is tightly coupled to Ca2+ translocation through the membrane, more than 40 Å away, by the A domain.[6]
It is interesting to note that the folding pattern and the locations of the critical amino acids for phosphorylation in P-type ATPases has the haloacid dehalogenase fold characteristic of the haloacid dehalogenase (HAD) superfamily, as predicted by sequence homology. The HAD superfamily functions on the common theme of an aspartate ester formation by an SN2 reaction mechanism. This SN2 reaction is clearly observed in the solved structure of SERCA with ADP plus AlF4-.[7]
Differences from SERCA1a
Various subfamilies of P-type ATPases also need additional subunits for proper function. Both P-IA and P-IV pumps needs extra subunits to function. The functional unit of Na+/K+-ATPase consists of two additional subunits, beta, and gamma, involved in trafficking, folding, and regulation of these pumps. SERCA1a and other P-IIA ATPases are also regulated by phospholamban and sarcolipin in vivo. Probably other subfamilies also need additional subunits for the proper function in vivo.
Some members of the family have additional domains fused to the pump. Heavy metal pumps can have several N- and C-terminal heavy metal-binding domains that have been found to be involved in regulation.
The proton pumps (IIIA) have a C-terminal regulatory domain (called the R domain), which, when unphosphorylated, inhibit pumping.
While most subfamilies have 10 transmembrane helices, there are some notable exceptions. The P-IA ATPases are predicted to have 7, and the large subfamily of heavy metal pumps (IB) is predicted to have 8 transmembrane helices. Type V appears to have a total of 12 transmembrane helices.
Mechanism
All P-type ATPases use the energy derived from ATP to drive vectorial transport. They form a high-energy aspartyl-phosphoranhydride intermediate in the reaction cycle, and they interconvert between at least two different conformations, denoted by E1 and E2. The E1-E2 notation stems from the initial studies on this family of enzymes made on the Na+,K+-ATPase, where the sodium form and the potassium form are referred to as E1 and E2, respectively, in the "Post-Albers scheme".
The E1-E2 schema has been proven to work, but there exist more than two major conformational states. However, the E1-E2 notation highlights the selectivity of the enzyme. In E1, the pump has high affinity for the exported substrate and low affinity for the imported substrate. In E2, it has low affinity of the exported substrate and high affinity for the imported substrate.
Four major enzyme states form the cornerstones in the reaction cycle. Several additional reaction intermediates occur interposed. These are termed E1~P, E2P, E2-P*, and E1/E2, described below.[8]
In the case of SERCA1a, energy from ATP is used to transport 2 Ca2+-ions from the cytoplasmic side to the lumen of the sarcoplasmatic reticulum, and to countertransport 1-3 protons into the cytoplasm.
Starting in the E1/E2 state, the reaction cycle begins as the enzyme releases 1-3 protons from the cation-ligating residues, in exchange for cytoplasmic Ca2+-ions. This leads to assembly of the phosphorylation site between the ATP-bound N domain and the P domain, while the A domain directs the occlusion of the bound Ca2+. In this occluded state, the Ca2+ ions are buried in a proteinacious environment with no access to either side of the membrane.
The Ca2E1~P state becomes formed through a kinase reaction, where the P domain becomes phosphorylated, producing ADP. The cleavage of the ß,-phosphordiester bond releases the gamma-phosphate from ADP and unleashes the N domain from the P domain.
This then allows the A domain to rotate towards the phosphorylation site, making a firm association with both the P and the N domain. This movement of the A domain exerts a downward push on M3-M4 and a drag on M1-M2, forcing the pump to open at the luminal side and forming the E2P state. During this transition, the transmembrane Ca2+-binding residues are forces apart, destroying the high-affinity binding site. This is in agreement with the general model form substrate translocation (cf. 1.2), showing that energy in primary transport is not used to bind the substrate but to release it again from the buried counter ions. At the same time the N domain becomes exposed to the cytosol, ready for ATP exchange at the nucleotide-binding site.
As the Ca2+ dissociate to the luminal side, the cation binding sites are neutralised by proton binding, and this make a closure of the transmembrane segments favourable. This closure is coupled to a downwards rotation of the A domain and a movement of the P domain, which then leads to the E2-P* occluded state. Meanwhile, the N domain exchanges ADP for ATP.
The P domain is dephosphorylated by the A domain, and the cycle completes when the phosphate is released from the enzyme, stimulated by the newly bound ATP, while a cytoplasmic pathway opens to exchange the protons for two new Ca2+-ions.[8].
See also
References
- ^ SKOU JC (February 1957). "The influence of some cations on an adenosine triphosphatase from peripheral nerves". Biochim. Biophys. Acta 23 (2): 394401. doi:10.1016/0006-3002(57)90343-8. PMID 13412736.
- ^ Axelsen KB, Palmgren MG (January 1998). "Evolution of substrate specificities in the P-type ATPase superfamily". J. Mol. Evol. 46 (1): 84101. doi:10.1007/PL00006286. PMID 9419228. http://link.springer-ny.com/link/service/journals/00239/bibs/46n1p84.html.
- ^ Lenoir G, Williamson P, Holthuis JC (December 2007). "On the origin of lipid asymmetry: the flip side of ion transport". Curr Opin Chem Biol 11 (6): 65461. doi:10.1016/j.cbpa.2007.09.008. PMID 17981493.
- ^ Ogawa H, Haga T, Toyoshima C (April 2000). "Soluble P-type ATPase from an archaeon, Methanococcus jannaschii". FEBS Lett. 471 (1): 99102. doi:10.1016/S0014-5793(00)01374-0. PMID 10760521. http://linkinghub.elsevier.com/retrieve/pii/S0014-5793(00)01374-0.
- ^ Stokes DL, Green NM (2003). "Structure and function of the calcium pump". Annu Rev Biophys Biomol Struct 32: 44568. doi:10.1146/annurev.biophys.32.110601.142433. PMID 12598367.
- ^ Toyoshima C, Nakasako M, Nomura H, Ogawa H (June 2000). "Crystal structure of the calcium pump of sarcoplasmic reticulum at 2.6 A resolution". Nature 405 (6787): 64755. doi:10.1038/35015017. PMID 10864315.
- ^ PDB 1T5T; Sørensen TL, Møller JV, Nissen P (June 2004). "Phosphoryl transfer and calcium ion occlusion in the calcium pump". Science 304 (5677): 1672–5. doi:10.1126/science.1099366. PMID 15192230.
- ^ a b Olesen C, Picard M, Winther AM et al. (December 2007). "The structural basis of calcium transport by the calcium pump". Nature 450 (7172): 103642. doi:10.1038/nature06418. PMID 18075584.
F- and V-type ATPase (3.A.2) P-type ATPase (3.A.3) - 3.A.3.1.1: Na+/K+ transporting: ATP1A1, ATP1A2, ATP1A3, ATP1A4, ATP1B1, ATP1B2, ATP1B3, ATP1B4, ATP1G1
- 3.A.3.1.2: H+/K+, H+/K+ exchanging: ATP4A, ATP4B
- 3.A.3.1.4: H+/K+ transporting, nongastric: ATP12A
- 3.A.3.2: Ca+ (SERCA, PMCA, SPCA) / Ca++ transporting: ATP2A1, ATP2A2, ATP2A3, ATP2B1, ATP2B2, ATP2B3, ATP2B4, ATP2C1
- 3.A.3.8.8: flippase: ATP8A2
Other/ungrouped:
see also ATPase disorders
B memb: cead, trns (1A, 1C, 1F, 2A, 3A1, 3A2-3, 3D), othrMembranes: Membrane transport Passive transport Diffusion: Facilitated diffusion ([Uniporter])
OsmosisActive transport -cytosis Exocytosis (Degranulation) - Endocytosis (Phagocytosis, transcytosis, fluid-phase pinocytosis, non-specific, adsorptive pinocytosis, Receptor-mediated endocytosis, Potocytosis, Efferocytosis)B memb: cead, trns (1A, 1C, 1F, 2A, 3A1, 3A2-3, 3D), othr Hydrolases: acid anhydride hydrolases (EC 3.6) 3.6.1 3.6.2 3.6.3-4: ATPase 3.6.3Cu++ (3.6.3.4)Ca+ (3.6.3.8)Na+/K+ (3.6.3.9)H+/K+ (3.6.3.10)ATP4AOther P-type ATPase3.6.43.6.5: GTPase 3.6.5.1: Heterotrimeric G protein3.6.5.2: Small GTPase > Ras superfamily3.6.5.3: Protein-synthesizing GTPase3.6.5.5-6: Polymerization motorsB enzm: 1.1/2/3/4/5/6/7/8/10/11/13/14/15-18, 2.1/2/3/4/5/6/7/8, 2.7.10, 2.7.11-12, 3.1/2/3/4/5/6/7, 3.1.3.48, 3.4.21/22/23/24, 4.1/2/3/4/5/6, 5.1/2/3/4/99, 6.1-3/4/5-6 Categories:- EC 3.6.3
- Membrane biology
- Integral membrane proteins
- Transport proteins
- Physiology
- Type I consists of the transition/heavy metal ATPases.

Wikimedia Foundation. 2010.