- V-ATPase
[
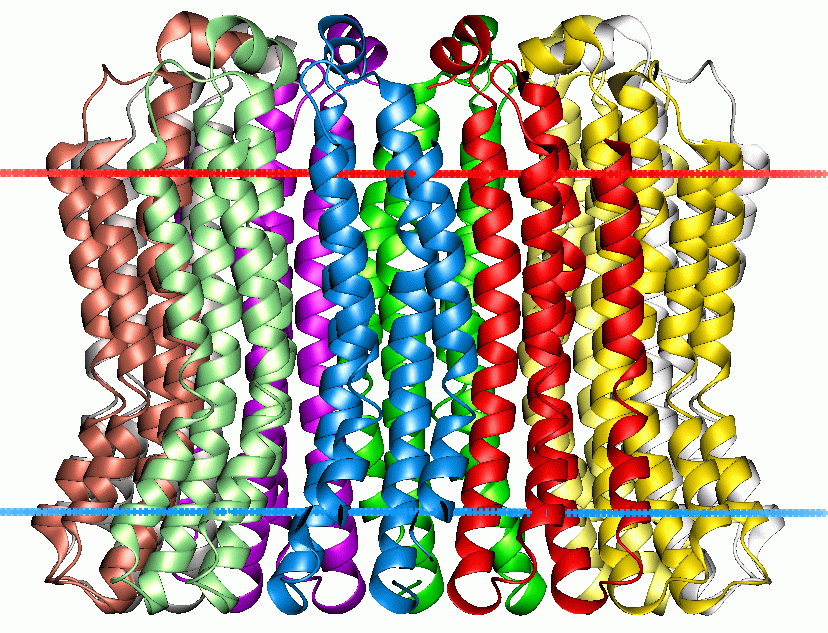
thumb|200px|V-type_sodium_ATPase from "Enterococcus hirae". Calculated hydrocarbon boundaries of thelipid bilayer are shown by red and blue dots]Vacuolar type H+-ATPase (V-ATPase) is a highly conserved evolutionarily ancient
enzyme with remarkably diverse functions in eukaryotic organisms. [cite journal |author=Nelson N, Perzov N, Cohen A, Hagai K, Padler V, Nelson H |title=The cellular biology of proton-motive force generation by V-ATPases |journal=J. Exp. Biol. |volume=203 |issue=Pt 1 |pages=89–95 |year=2000 |pmid=10600677 |url=http://jeb.biologists.org/cgi/reprint/203/1/89] V-ATPases acidifiy a wide array of intracellularorganelles and pump protons across the plasma membranes of numerous cell types. V-ATPases couple the energy ofATP hydrolysis to proton transport across intracellular and plasma membranes of eukaryotic cells.Roles played by V-ATPases
V-ATPases are found within the membranes of many organelles, such as
endosomes ,lysosomes and secretory vesicles where they play a variety of roles crucial for the function of these organelles. For example, the proton gradient across the yeast vacuolar membrane generated by V-ATPases drives calcium uptake into the vacuole through an H+/Ca2+ antiporter system (Ohya, 1991). V-ATPases also play an important role in synaptic transmission in neuronal cells.Norepinephrine enters vesicles in exchange for protons pumped by V-ATPase.V-ATPases are also found in the plasma membranes of a wide variety of cells such as
intercalated cells of thekidney ,osteoclasts (bone resorbing cells),macrophages ,neutrophils , sperm,midgut cells ofinsect s and certaintumor cells. Plasma membrane V-ATPases are involved in processes such aspH homeostasis , coupled transport and tumormetastasis . V-ATPases in the acrosomal membrane of sperm acidify theacrosome . This acidification activatesproteases required to drill through theplasma membrane of the egg. V-ATPases in the osteoclast plasma membrane pump protons onto the bone surface which is necessary for bone resorption. In the intercalated cells of the kidney, V-ATPases pump protons into theurine , allowing forbicarbonate reabsorption into the blood.V-ATPase structure
The
yeast V-ATPase is the best characterized. There are at least 13 subunits identified to form a functional V-ATPase complex, which consists of two domains. The subunits belong to either the Vo domain (membrane associated subunits, lower case letters on the figure), or the V1 domain (peripherally associated subunits, upper case letters on the figure).The V1 includes 8 subunits, A-H, with three copies of the A and B subunits, one or two copies of E, and two copies of subunit G. The V1 domain contains tissue specific subunit isoforms including B, C, E, and G. Mutations to the B1 isoform result in the human disease distal renal tubular
acidosis and sensorineural deafness.The Vo domain contains 6 different subunits, a, d, c, c', c" and e, with possibly four copies of c and one copy of the remaining subunits. The mammalian Vo domain contains tissue specific isoforms for subunits a and d, while yeast V-ATPase contains two organelle specific subunit isoforms of a, Vph1p and Stv1p. Mutations to the a3 isoform result in the human disease infantile malignant
osteopetrosis , and mutations to the a4 isoform result in distal renal tubular acidosis, in some cases with sensorineural deafness.The V1 domain is responsible for ATP hydrolysis whereas the Vo domain is responsible for proton translocation. ATP hydrolysis at the catalytic nucleotide binding sites on subunit A drives rotation of a central stalk composed of subunits D and F, which in turn drives rotation of a barrel of c subunits relative to the a subunit. The amino-terminal of a (NT-a) along with subunits C, E, G and H compose the peripheral stalk. The carboxy-terminal of subunit a (CT-a) is held fixed relative to the A3B3 head by this peripheral stalk. Movement of the barrel of c subunits past the a subunit is thought to drive proton transport across the membrane. A
stoichiometry of two protons translocated for each ATP hydrolyzed has been proposed by (Johnson, 1982).In addition to the structural subunits of yeast V-ATPase, associated proteins have been identified that are necessary for assembly. These associated proteins are essential for Vo domain assembly and are termed Vma12p, Vma21p and Vma22p (Hirata, 1993; Ho, 1993; Hill, 1994; Jackson, 1997). Two of the three proteins, Vma12p and Vma22p form a complex that binds transiently to Vph1p (subunit a) to aid its assembly and maturation (Hill, 1994; Hill, 1995; Graham, 1998; Graham, 2003). Vma21p coordinates assembly of the Vo subunits as well as escorting the Vo domain into vesicles for transport to the Golgi (Malkus, 2004).
V-ATPase assembly
Yeast V-ATPases fail to assemble when any of the genes that encode subunits are deleted except for subunits H and c" (Whyteside, 2005; Forgac, 1999; Stevens, 1997). Without subunit H, the assembled V-ATPase is not active (Ho, 1993; Parra, 2000) and the loss of the c" subunit results in uncoupling of enzymatic activity (Whyteside, 2005).
The precise mechanisms by which V-ATPases assembly are still controversial with evidence suggesting two different possibilities. Mutational analysis and "in vitro" assays have shown that preassembled Vo and V1 domains can combine to form one complex in a process called independent assembly. Support for independent assembly includes the findings that the assembled Vo domain can be found at the vacuole in the absence of the V1 domain, whereas free V1 domains can be found in the
cytoplasm and not at thevacuole (Kane, 1995; Sumner, 1995). In contrast, "in vivo" pulse-chase experiments have revealed early interactions between Vo and V1 subunits, specifically the a and B subunits, suggesting that subunits are added in a step-wise fashion to form a single complex in a concerted assembly process (Kane, 1999).Regulation of V-ATPase activity
"In vivo" regulation of V-ATPase activity is accomplished by reversible dissociation of the V1 domain from the Vo domain. After initial assembly, both the insect
Manduca sexta and yeast V-ATPases can reversibly disassemble into free Vo and V1 domains after a 2-5 min deprivation of glucose (Kane, 1995). Reversible disassembly may be a general mechanism of regulating V-ATPase activity since it exists in yeast and insects. Reassembly is proposed to be aided by a complex termedRAVE (regulator of H+-ATPase of vacuolar and endosomal membranes) (Kane and Smardon, 2003). Interestingly, dissasembly and reassembly of V-ATPases does not require new protein synthesis but does need an intact microtubular network (Holliday, 2000).Human diseases
Osteopetrosis
Osteopetrosis is generic name that represents a group of heritable conditions in which there is a defect in osteoclastic bone resorption. Both dominant and recessive osteopetrosis occur in humans {Michigami, 2002; Frattini, 2000}. Autosomal dominant osteopetrosis shows mild symptoms in adults who experience frequent bone fractures due to brittle bones {Michigami, 2002}. A form of osteopetrosis that is clinically more severe is termed autosomal recessive infantile malignant osteopetrosis {Frattini, 2000; Sobacchi, 2001; Fasth, 1999}. Three genes have been identified which are responsible for recessive osteopetrosis in humans. Interestingly, they are all directly involved in the proton generation and secretion pathways that are essential for bone resorption. One gene iscarbonic anhydrase II (CAII) that when mutated causes osteopetrosis withrenal tubular acidosis (type 3) {Sly, 1983}. Mutations to the chloride channel ClC7 gene also lead to both dominant and recessive osteopetrosis {Michigami, 2002}. Approximately 50% of patients with recessive infantile malignant osteopetrosis have mutations to the a3 subunit isoform of V-ATPase {Sobacchi, 2001; Kornak, 2000; Frattini, 2003}. In humans, 26 mutations have been identified in V-ATPase subunit isoform a3, found in osteoclasts, that result in the bone disease autosomal recessive osteopetrosis {Frattini, 2000; Kornak, 2000; Sobacchi, 2001; Susani, 2004}Distal renal tubular acidosis (dRTA)
The importance of V-ATPase activity in renal proton secretion is highlighted by the inherited disease distal
renal tubular acidosis . In all cases, renal tubular acidosis results from a failure of the normalrenal mechanisms that regulate systemic pH. There are four types of renal tubular acidosis. Type 1 is distal renal tubular acidosis and results from a failure of thecortical collecting duct to acidify the urine below pH 5. {Alper, 2002}. Some patients withautosomal recessive dRTA also havesensorineural hearing loss {Karet, 1999}. Inheritance of this type of RTA results from mutations to either V-ATPase subunit isoforms B1 or a4, or mutations ofband 3 (also called AE1), a Cl-/HCO3- exchanger {Stehberger, 2003; Karet, 1999; Karet, 1998}. Twelve different mutations to V-ATPase isoform B1 (Stover, 2002) and twenty-four different mutations in a4 lead to dRTA {Smith, 2000; Karet, 1999; Stover, 2005}.Reverse transcription polymerase chain reaction studies have shown expression of the a4 subunit in the intercalated cell of the kidney and in thecochlea {Stover, 2002}. dRTA caused by mutations in the a4 subunit gene in some cases can be associated with deafness due to a failure to normally acidify theendolymph of theinner ear {Stehberger, 2003}.Nomenclature
The term Vo has a lowercase letter "o" (not the number "zero") in subscript. The "o" stands for
oligomycin .See also
*
ATPases
*F-ATPase
*Na+/K+-ATPase References
*Alper, S. L. "Genetic diseases of acid-base transporters." Annu Rev Physiol, 2002. 64: p. 899-923. PMID 11826292
*Fasth, A., and Porras, O. "Human malignant osteopetrosis: pathophysiology, management and the role of bone marrow transplantation." Pediatr Transplant, 1999. 3 Suppl 1: p. 102-7. PMID 10587979
*Forgac, M. "The vacuolar H+-ATPase of clathrin-coated vesicles is reversibly inhibited by S-nitrosoglutathione." J Biol Chem, 1999. 274(3): p. 1301-5. PMID 9880499
*Frattini, A., "et al.", "Defects in TCIRG1 subunit of the vacuolar proton pump are responsible for a subset of human autosomal recessive osteopetrosis." Nat Genet, 2000. 25(3): p. 343-6. PMID 10888887
*Frattini, A., "et al.", "Chloride channel ClCN7 mutations are responsible for severe recessive, dominant, and intermediate osteopetrosis." J Bone Miner Res, 2003. 18(10): p. 1740-7. PMID 14584882
*Graham, L. A., Hill, K. J., and Stevens, T. H. "Assembly of the yeast vacuolar H+-ATPase occurs in the endoplasmic reticulum and requires a Vma12p/Vma22p assembly complex." J Cell Biol, 1998. 142(1): p. 39-49. PMID 9660861
*Graham, L. A., Flannery, A. R., and Stevens, T. H. "Structure and assembly of the yeast V-ATPase." J Bioenerg Biomembr, 2003. 35(4): p. 301-12. PMID 14635776
*Hill, K. J., "et al.", "Vma21p is a yeast membrane protein retained in the endoplasmic reticulum by a di-lysine motif and is required for the assembly of the vacuolar H(+)-ATPase complex." Molecular Biology of the Cell, 1994. 5(9): p. 1039-50. PMID 7841520
*Hill, K. J., and Stevens, T. H. "Vma22p is a novel endoplasmic reticulum-associated protein required for assembly of the yeast vacuolar H(+)-ATPase complex." The Journal of Biological Chemistry, 1995. 270(38): p. 22329-36. PMID 7673216
*Hirata, R., "et al.", "VMA12 is essential for assembly of the vacuolar H(+)-ATPase subunits onto the vacuolar membrane in Saccharomyces cerevisiae." Journal of Biological Chemistry, 1993. 268(2): p. 961-7. PMID 8419376
*Ho, M. N., "et al.", "VMA13 encodes a 54-kDa vacuolar H(+)-ATPase subunit required for activity but not assembly of the enzyme complex in Saccharomyces cerevisiae." Journal of Biological Chemistry, 1993. 268(1): p. 221-7. PMID 8349704
*Holliday, L. S., "et al.", "The amino-terminal domain of the B subunit of vacuolar H+-ATPase contains a filamentous actin binding site." J Biol Chem, 2000. 275(41): p. 32331-7. PMID 10915794
*Jackson, D. D., and Stevens, T. H. "VMA12 encodes a yeast endoplasmic reticulum protein required for vacuolar H+-ATPase assembly." J Biol Chem, 1997. 272(41): p. 25928-34. PMID 9325326
*Johnson, R. G., "et al.", "H+ ATPase of chromaffin granules. Kinetics, regulation, and stoichiometry." J Biol Chem, 1982. 257(18): p. 10701-7. PMID 6213624
*Kane, P. M. "Disassembly and reassembly of the yeast vacuolar H(+)-ATPase "in vivo"." J Biol Chem, 1995. 270(28): p. 17025-32. PMID 7622524
*Kane, P. M., "et al.", "Early steps in assembly of the yeast vacuolar H+-ATPase." J Biol Chem, 1999. 274(24): p. 17275-83. PMID 10358087
*Kane, P.M., and Smardon, A.M., "Assembly and regulation of the yeast vacuolar H+-ATPase." J Bioenerg Biomembr, 2003. 35(4) : p. 313-21. PMID 14635777
*Karet, F. E., "et al.", "Mutations in the chloride-bicarbonate exchanger gene AE1 cause autosomal dominant but not autosomal recessive distal renal tubular acidosis." Proc Natl Acad Sci U S A, 1998. 95: p. 6337-42. PMID 9600966
*Karet, F. E., "et al.", "Mutations in the gene encoding B1 subunit of H+-ATPase cause renal tubular acidosis with sensorineural deafness." Nat Genet, 1999. 21(1): p. 84-90. PMID 9916796
*Kornak, U., "et al.", "Mutations in the a3 subunit of the vacuolar H(+)-ATPase cause infantile malignant osteopetrosis." Hum Mol Genet, 2000. 9(13): p. 2059-63. PMID 10942435
*Malkus, P., "et al.", "Role of Vma21p in assembly and transport of the yeast vacuolar ATPase." Mol Biol Cell, 2004. 15: p. 5075-91. PMID 15356264
*Michigami, T., "et al.", "Novel mutations in the a3 subunit of vacuolar H(+)-adenosine triphosphatase in a Japanese patient with infantile malignant osteopetrosis." Bone, 2002. 30(2),: p. 436-9. PMID 11856654
*Ohya, Y., "et al.", "Calcium-sensitive cls mutants of Saccharomyces cerevisiae showing a Pet- phenotype are ascribable to defects of vacuolar membrane H(+)-ATPase activity." J Biol Chem, 1991. 266(21): p. 13971-7. PMID 1830311
*Parra, K. J., "et al.", "The H subunit (Vma13p) of the yeast V-ATPase inhibits the ATPase activity of cytosolic V1 complexes." J Biol Chem, 2000. 275(28): p. 21761-7. PMID 10781598
*Sly, W. S., "et al.", "Carbonic anhydrase II deficiency identified as the primary defect in the autosomal recessive syndrome of osteopetrosis with renal tubular acidosis and cerebral calcification." Proc Natl Acad Sci U S A, 1983. 80: p. 2752-6. PMID 6405388
*Sobacchi, C., "et al.", "The mutational spectrum of human malignant autosomal recessive osteopetrosis." Hum Mol Genet, 2001. 10(17): p. 1767-73. PMID 11532986
*Stehberger, P. A., "et al.", "Localization and regulation of the ATP6V0A4 (a4) vacuolar H+-ATPase subunit defective in an inherited form of distal renal tubular acidosis." J Am Soc Nephrol, 2003. 14(12): p. 3027-38. PMID 14638902
*Stevens, T. H., and Forgac, M. "Structure, function and regulation of the vacuolar (H+)-ATPase." Annu. Rev. Cell Dev. Biol., 1997. 13: p. 779-808. PMID 9442887
*Stover, E. H., "et al.", "Novel ATP6V1B1 and ATP6V0A4 mutations in autosomal recessive distal renal tubular acidosis with new evidence for hearing loss." J Med Genet, 2002. 39(11): p. 796-803. PMID 12414817
*Sumner, J.P., "et al.", "Regulation of plasma membrane V-ATPase activity by dissociation of peripheral subunits." J Biol Chem, 1995. 270(10): p. 5649-53. PMID 7890686
*Susani, L., "et al.", "TCIRG1-dependent recessive osteopetrosis: mutation analysis, functional identification of the splicing defects, and "in vitro" rescue by U1 snRNA." Hum Mutat, 2004. 24(3): p. 225-35. PMID 15300850
*Whyteside, G., "et al.", "Assembly of the yeast vacuolar H+-ATPase and ATP hydrolysis occurs in the absence of subunit c"." FEBS Letters, 2005. 579: p. 2981-5. PMID 15907326External links
* - Orientations of proton or sodium translocating F- and V-type ATPases in membrane
*
Wikimedia Foundation. 2010.