- Paleontology
-
"Palaeontology" redirects here. For the scientific journal, see Palaeontology (journal).
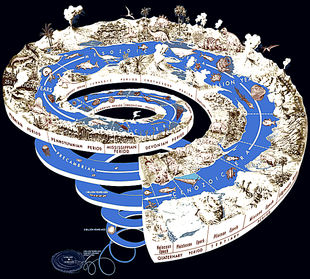 Paleontology studies the entire history of life on Earth.
Paleontology studies the entire history of life on Earth.
Paleontology (pronounced /ˌpælɪɒnˈtɒlədʒi/; British: palaeontology; from Greek: παλαιός (palaeos) "old, ancient", ὄν, ὀντ- (on, ont-) "being, creature", and λόγος (logos) "speech, thought") is the study of prehistoric life. It includes the study of fossils to determine organisms' evolution and interactions with each other and their environments (their paleoecology). As a "historical science" it attempts to explain causes rather than conduct experiments to observe effects. Paleontological observations have been documented as far back as the 5th century BC. The science became established in the 18th century as a result of Georges Cuvier's work on comparative anatomy, and developed rapidly in the 19th century. Fossils found in China since the 1990s have provided new information about the earliest evolution of animals, early fish, dinosaurs and the evolution of birds and mammals. Paleontology lies on the border between biology and geology, and shares with archaeology a border that is difficult to define. It now uses techniques drawn from a wide range of sciences, including biochemistry, mathematics and engineering. As knowledge has increased, paleontology has developed specialized sub-divisions, some of which focus on different types of fossil organisms while others study ecology and environmental history, such as ancient climates.
Body fossils and trace fossils are the principal types of evidence about ancient life, and geochemical evidence has helped to decipher the evolution of life before there were organisms large enough to leave fossils. Estimating the dates of these remains is essential but difficult: sometimes adjacent rock layers allow radiometric dating, which provides absolute dates that are accurate to within 0.5%, but more often paleontologists have to rely on relative dating by solving the "jigsaw puzzles" of biostratigraphy. Classifying ancient organisms is also difficult, as many do not fit well into the Linnean taxonomy that is commonly used for classifying living organisms, and paleontologists more often use cladistics to draw up evolutionary "family trees". The final quarter of the 20th century saw the development of molecular phylogenetics, which investigates how closely organisms are related by measuring how similar the DNA is in their genomes. Molecular phylogenetics has also been used to estimate the dates when species diverged, but there is controversy about the reliability of the molecular clock on which such estimates depend.
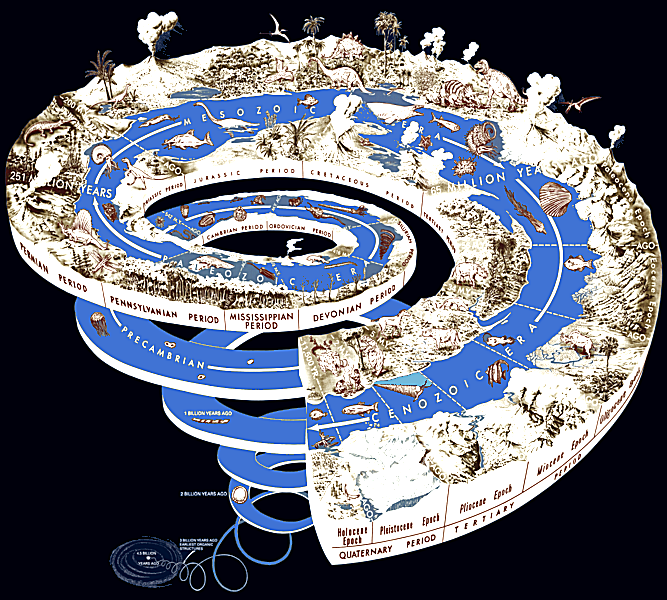
Use of all these techniques has enabled paleontologists to discover much of the evolutionary history of life, almost all the way back to when Earth became capable of supporting life, about 3,800 million years ago. For about half of that time the only life was single-celled micro-organisms, mostly in microbial mats that formed ecosystems only a few millimeters thick. Earth's atmosphere originally contained virtually no oxygen, and its oxygenation began about 2,400 million years ago. This may have caused an accelerating increase in the diversity and complexity of life, and early multicellular plants and fungi have been found in rocks dated from 1,700 to 1,200 million years ago. The earliest multicellular animal fossils are much later, from about 580 million years ago, but animals diversified very rapidly and there is a lively debate about whether most of this happened in a relatively short Cambrian explosion or started earlier but has been hidden by lack of fossils. All of these organisms lived in water, but plants and invertebrates started colonizing land from about 490 million years ago and vertebrates followed them about 370 million years ago. The first dinosaurs appeared about 230 million years ago and birds evolved from one dinosaur group about 150 million years ago. During the time of the dinosaurs, mammals' ancestors survived only as small, mainly nocturnal insectivores, but after the non-avian dinosaurs became extinct in the Cretaceous–Tertiary extinction event 65 million years ago mammals diversified rapidly. Flowering plants appeared and rapidly diversified between 130 million years ago and 90 million years ago, possibly helped by coevolution with pollinating insects. Social insects appeared around the same time and, although they have relatively few species, now form over 50% of the total mass of all insects. Humans evolved from a lineage of upright-walking apes whose earliest fossils date from over 6 million years ago, and anatomically modern humans appeared under 200,000 years ago. The course of evolution has been changed several times by mass extinctions that wiped out previously dominant groups and allowed other to rise from obscurity to become major components of ecosystems.
Contents
Definition
 A paleontologist carefully chips rock from a column of dinosaur vertebrae.
A paleontologist carefully chips rock from a column of dinosaur vertebrae. A paleontologist at work at John Day Fossil Beds National Monument
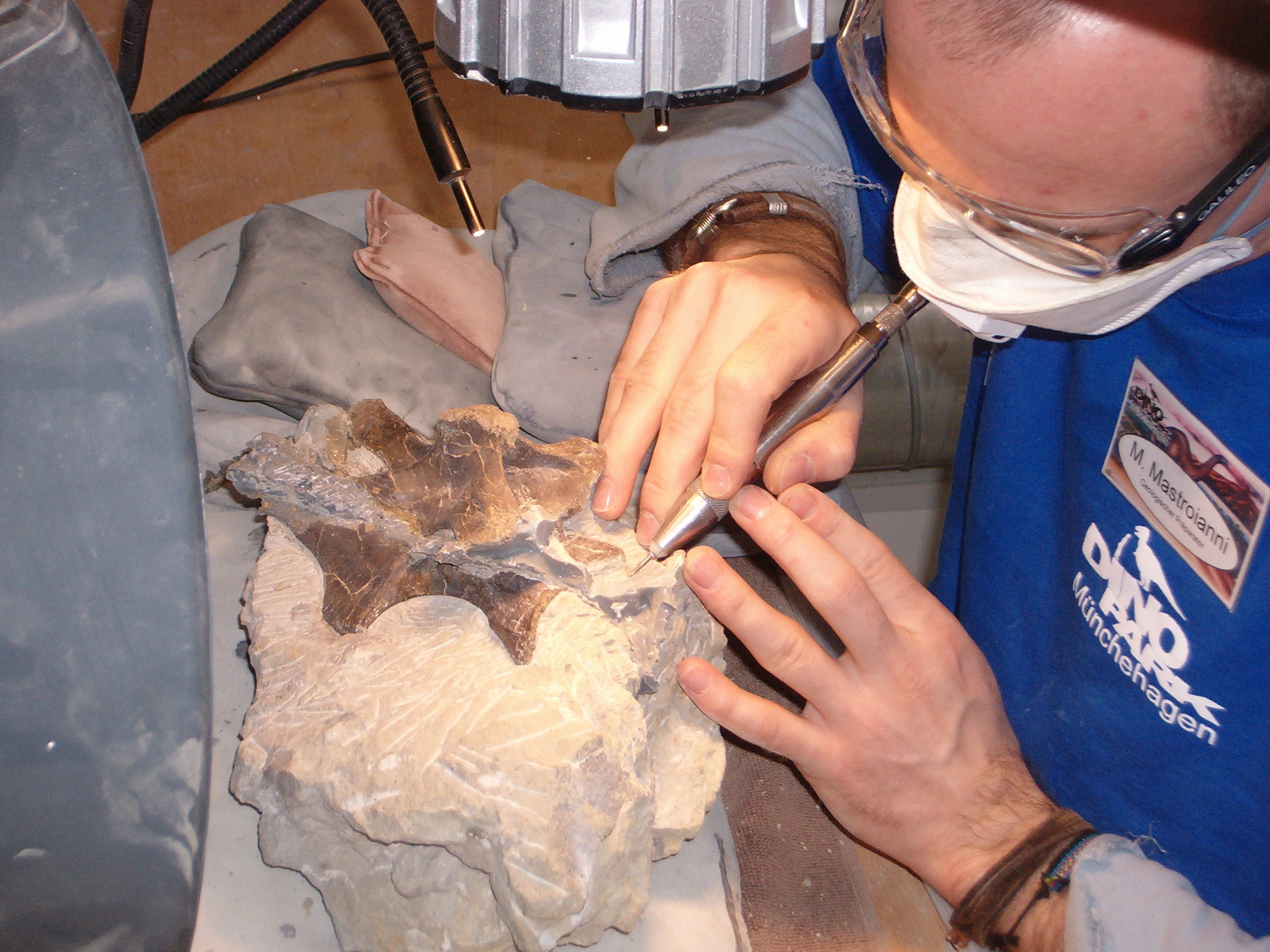
A paleontologist at work at John Day Fossil Beds National Monument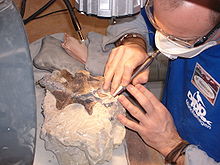 The preparation of the fossilized bones of Europasaurus holgeri
The preparation of the fossilized bones of Europasaurus holgeriThe simplest definition is "the study of ancient life".[1] Paleontology seeks information about several aspects of past organisms: "their identity and origin, their environment and evolution, and what they can tell us about the Earth's organic and inorganic past".[2]
A historical science
Paleontology is one of the historical sciences, along with archaeology, geology, biology, astronomy, cosmology, philology and history itself.[3] This means that it aims to describe phenomena of the past and reconstruct their causes.[4] Hence it has three main elements: description of the phenomena; developing a general theory about the causes of various types of change; and applying those theories to specific facts.[3]
When trying to explain past phenomena, paleontologists and other historical scientists often construct a set of hypotheses about the causes and then look for a "smoking gun", a piece of evidence which indicates that one of the hypotheses is a better explanation than the others. Sometimes the "smoking gun" is discovered by a fortunate accident during other research, for example the discovery by Luis Alvarez and Walter Alvarez of an iridium-rich layer at the Cretaceous–Tertiary boundary made asteroid impact and volcanism the most favored explanations for the Cretaceous–Tertiary extinction event.[4]
The other main type of science is experimental science, which is often said to work by conducting experiments to disprove hypotheses about the workings and causes of natural phenomena – note that this approach cannot prove a hypothesis is correct, since some later experiment may disprove it. However, when confronted with totally unexpected phenomena, such as the first evidence for invisible radiation, experimental scientists often use the same approach as historical scientists: construct a set of hypotheses about the causes and then look for a "smoking gun".[4]
Related sciences
Paleontology lies on the boundary between biology and geology since paleontology focuses on the record of past life but its main source of evidence is fossils, which are found in rocks.[5] For historical reasons paleontology is part of the geology departments of many universities, because in the 19th century and early 20th century geology departments found paleontological evidence important for estimating the ages of rocks while biology departments showed little interest.[6]
Paleontology also has some overlap with archaeology, which primarily works with objects made by humans and with human remains, while paleontologists are interested in the characteristics and evolution of humans as organisms. When dealing with evidence about humans, archaeologists and paleontologists may work together – for example paleontologists might identify animal or plant fossils around an archaeological site, to discover what the people who lived there ate; or they might analyze the climate at the time when the site was inhabited by humans.[7]
 Analyses using engineering techniques show that Tyrannosaurus had a devastating bite, but raise doubts about how fast it could move.
Analyses using engineering techniques show that Tyrannosaurus had a devastating bite, but raise doubts about how fast it could move.In addition paleontology often uses techniques derived from other sciences, including biology, ecology, chemistry, physics and mathematics.[1] For example geochemical signatures from rocks may help to discover when life first arose on Earth,[8] and analyses of carbon isotope ratios may help to identify climate changes and even to explain major transitions such as the Permian–Triassic extinction event.[9] A relatively recent discipline, molecular phylogenetics, often helps by using comparisons of different modern organisms' DNA and RNA to re-construct evolutionary "family trees"; it has also been used to estimate the dates of important evolutionary developments, although this approach is controversial because of doubts about the reliability of the "molecular clock".[10] Techniques developed in engineering have been used to analyse how ancient organisms might have worked, for example how fast Tyrannosaurus could move and how powerful its bite was.[11][12]
A combination of paleontology, biology, and archaeology, paleoneurology is the study of endocranial casts (or endocasts), of species related to humans in order to analyze the evolution of human brains. [13]
Paleontology even contributes to astrobiology, the investigation of possible life on other planets, by developing models of how life may have arisen and by providing techniques for detecting evidence of life.[14]
Subdivisions
As knowledge has increased, paleontology has developed specialised subdivisons.[15] Vertebrate paleontology concentrates on fossils of vertebrates, from the earliest fish to the immediate ancestors of modern mammals. Invertebrate paleontology deals with fossils of invertebrates such as molluscs, arthropods, annelid worms and echinoderms. Paleobotany focuses on the study of fossil plants, but traditionally includes the study of fossil algae and fungi. Palynology, the study of pollen and spores produced by land plants and protists, straddles the border between paleontology and botany, as it deals with both living and fossil organisms. Micropaleontology deals with all microscopic fossil organisms, regardless of the group to which they belong.[16]
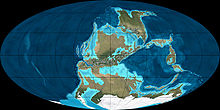 In the Carboniferous period, the continents were not in the same places as they are today, and there was extensive glaciation.
In the Carboniferous period, the continents were not in the same places as they are today, and there was extensive glaciation.Instead of focusing on individual organisms, paleoecology examines the interactions between different organisms, such as their places in food chains, and the two-way interaction between organisms and their environment[17] – for example the development of oxygenic photosynthesis by bacteria hugely increased the productivity and diversity of ecosystems,[18] and also caused the oxygenation of the atmosphere, which in turn was a prerequisite for the evolution of the most complex eucaryotic cells, from which all multicellular organisms are built.[19] Paleoclimatology, although sometimes treated as part of paleoecology,[16] focuses more on the history of Earth's climate and the mechanisms which have changed it[20] – which have sometimes included evolutionary developments, for example the rapid expansion of land plants in the Devonian period removed more carbon dioxide from the atmosphere, reducing the greenhouse effect and thus helping to cause an ice age in the Carboniferous period.[21]
Biostratigraphy, the use of fossils to work out the chronological order in which rocks were formed, is useful to both paleontologists and geologists.[22] Biogeography studies the spatial distribution of organisms, and is also linked to geology, which explains how Earth's geography has changed over time.[23]
Sources of evidence
Body fossils
 This Marrella specimen illustrates how clear and detailed the fossils from the Burgess Shale lagerstätte are.
This Marrella specimen illustrates how clear and detailed the fossils from the Burgess Shale lagerstätte are.Fossils of organisms' bodies are usually the most informative type of evidence. The most common types are wood, bones, and shells.[24] Fossilisation is a rare event, and most fossils are destroyed by erosion or metamorphism before they can be observed. Hence the fossil record is very incomplete, increasingly so further back in time. Despite this, it is often adequate to illustrate the broader patterns of life's history.[25] There are also biases in the fossil record: different environments are more favorable to the preservation of different types of organism or parts of organisms.[26] Further, only the parts of organisms that were already mineralised are usually preserved, such as the shells of molluscs. Since most animal species are soft-bodied, they decay before they can become fossilised. As a result, although there are 30-plus phyla of living animals, two-thirds have never been found as fossils.[27]
Occasionally, unusual environments may preserve soft tissues. These lagerstätten allow paleontologists to examine the internal anatomy of animals that in other sediments are represented only by shells, spines, claws, etc. – if they are preserved at all. However, even lagerstätten present an incomplete picture of life at the time. The majority of organisms living at the time are probably not represented because lagerstätten are restricted to a narrow range of environments, e.g. where soft-bodied organisms can be preserved very quickly by events such as mudslides; and the exceptional events that cause quick burial make it difficult to study the normal environments of the animals.[28] The sparseness of the fossil record means that organisms are expected to exist long before and after they are found in the fossil record – this is known as the Signor-Lipps effect.[29]
Trace fossils
 Main article: Trace fossil
Main article: Trace fossilTrace fossils consist mainly of tracks and burrows, but also include coprolites (fossil feces) and marks left by feeding.[24][30] Trace fossils are particularly significant because they represent a data source that is not limited to animals with easily-fossilized hard parts, and which reflects organisms' behaviour. Also many traces date from significantly earlier than the body fossils of animals that are thought to have been capable of making them.[31] Whilst exact assignment of trace fossils to their makers is generally impossible, traces may for example provide the earliest physical evidence of the appearance of moderately complex animals (comparable to earthworms).[30]
Geochemical observations
Main article: GeochemistryGeochemical observations may help to deduce the global level of biological activity, or the affinity of a certain fossil. For example geochemical features of rocks may reveal when life first arose on Earth,[8] and may provide evidence of the presence of eucaryotic cells, the type from which all multicellular organisms are built.[32] Analyses of carbon isotope ratios may help to explain major transitions such as the Permian–Triassic extinction event.[9]
Classifying ancient organisms
Main articles: Biological classification , Cladistics , Phylogenetic nomenclature , and Evolutionary taxonomySimple example cladogramTetrapods Amniotes Synapsids Extinct Synapsids
Reptiles Extinct reptiles
Archosaurs
?Extinct
ArchosaursCrocodilians
Dinosaurs
?Extinct
Dinosaurs
?
Warm-bloodedness evolved somewhere in the
synapsid–mammal transition.
? Warm-bloodedness must also have evolved at one of
these points – an example of convergent evolution.[33]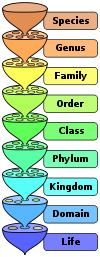 Levels in the Linnean taxonomy
Levels in the Linnean taxonomyNaming groups of organisms in a way that is clear and widely agreed is important, as some disputes in palaeontology have been based just on misunderstandings over names.[34] Linnean taxonomy is commonly used for classifying living organisms, but runs into difficulties when dealing with newly-discovered organisms that are significantly different from known ones. For example: it is hard to decide at what level to place a new higher-level grouping, e.g. genus or family or order; this is important since the Linnean rules for naming groups are tied to their levels, and hence if a group is moved to a different level it has to be renamed.[35]
Paleontologists generally use approaches based on cladistics, a technique for working out the evolutionary "family tree" of a set of organisms.[34] It works by the logic that, if groups B and C have more similarities to each other than either has to group A, then B and C are more closely related to each other than either is to A. Characters that are compared may be anatomical, such as the presence of a notochord, or molecular, by comparing sequences of DNA or proteins. The result of a successful analysis is a hierarchy of clades – groups that share a common ancestor. Ideally the "family tree" has only two branches leading from each node ("junction"), but sometimes there is too little information to achieve this and paleontologists have to make do with junctions that have several branches. The cladistic technique is sometimes fallible, as some features, such as wings or camera eyes, evolved more than once, convergently – this must be taken into account in analyses.[33]
Evolutionary developmental biology, commonly abbreviated to "Evo Devo", also helps paleontologists to produce "family trees". For example the embryological development of some modern brachiopods suggests that brachiopods may be descendants of the halkieriids, which became extinct in the Cambrian period.[36]
Estimating the dates of organisms
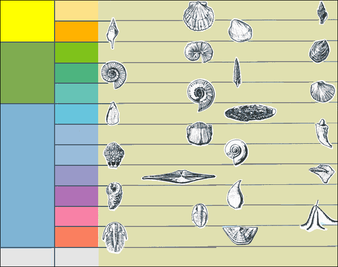 Pecten gibbusBathyurus extansNeptunea tabulataNerinea trinodosaProlecanites gurleyiHexamocaras hertzeriTetragraptus fructicosusBillingsella corrugataCommon index fossils used to date rocks in North-East USA
Pecten gibbusBathyurus extansNeptunea tabulataNerinea trinodosaProlecanites gurleyiHexamocaras hertzeriTetragraptus fructicosusBillingsella corrugataCommon index fossils used to date rocks in North-East USAPaleontology seeks to map out how living things have changed through time. A substantial hurdle to this aim is the difficulty of working out how old fossils are. Beds which preserve fossils typically lack the radioactive elements needed for radiometric dating. This technique is our only means of giving rocks greater than about 50 million years old an absolute age, and can be accurate to within 0.5% or better.[37] Although radiometric dating requires very careful laboratory work, its basic principle is simple: the rates at which various radioactive elements decay are known, and so the ratio of the radioactive element to the element into which it decays shows how long ago the radioactive element was incorporated into the rock. Radioactive elements are common only in rocks with a volcanic origin, and so the only fossil-bearing rocks that can be dated radiometrically are a few volcanic ash layers.[37]
Consequently, paleontologists must usually rely on stratigraphy to date fossils. Stratigraphy is the science of deciphering the "layer-cake" that is the sedimentary record, and has been compared to a jigsaw puzzle.[38] Rocks normally form relatively horizontal layers, with each layer younger than the one underneath it. If a fossil is found between two layers whose ages are known, the fossil's age must lie between the two known ages.[39] Because rock sequences are not continuous, but may be broken up by faults or periods of erosion, it is very difficult to match up rock beds that are not directly next to one another. However, fossils of species that survived for a relatively short time can be used to link up isolated rocks: this technique is called biostratigraphy. For instance, the conodont Eoplacognathus pseudoplanus has a short range in the Middle Ordovician period.[40] If rocks of unknown age are found to have traces of E. pseudoplanus, they must have a mid-Ordovician age. Such index fossils must be distinctive, be globally distributed and have a short time range to be useful. However, misleading results are produced if the index fossils turn out to have longer fossil ranges than first thought.[41] Stratigraphy and biostratigraphy can in general provide only relative dating (A was before B), which is often sufficient for studying evolution. However, this is difficult for some time periods, because of the problems involved in matching up rocks of the same age across different continents.[42]
Family-tree relationships may also help to narrow down the date when lineages first appeared. For instance, if fossils of B or C date to X million years ago and the calculated "family tree" says A was an ancestor of B and C, then A must have evolved more than X million years ago.
It is also possible to estimate how long ago two living clades diverged – i.e. approximately how long ago their last common ancestor must have lived – by assuming that DNA mutations accumulate at a constant rate. These "molecular clocks", however, are fallible, and provide only a very approximate timing: for example, they are not sufficiently precise and reliable for estimating when the groups that feature in the Cambrian explosion first evolved,[43] and estimates produced by different techniques may vary by a factor of two.[10]
Overview of the history of life
Main article: Evolutionary history of lifeThe evolutionary history of life stretches back to over 3,000 million years ago, possibly as far as 3,800 million years ago. Earth formed about 4,570 million years ago and, after a collision that formed the Moon about 40 million years later, may have cooled quickly enough to have oceans and an atmosphere about 4,440 million years ago.[44] However there is evidence on the Moon of a Late Heavy Bombardment from 4,000 to 3,800 million years ago. If, as seem likely, such a bombardment struck Earth at the same time, the first atmosphere and oceans may have been stripped away.[45] The oldest clear evidence of life on Earth dates to 3,000 million years ago, although there have been reports, often disputed, of fossil bacteria from 3,400 million years ago and of geochemical evidence for the presence of life 3,800 million years ago.[8][46] Even the simplest modern organisms are too complex to have emerged directly from non-living materials.[47] Some scientists have proposed that life on Earth was "seeded" from elsewhere,[48] but most research concentrates on various explanations of how life could have arisen independently on Earth.[49]
 This wrinkled "elephant skin" texture is a trace fossil of a non-stromatolite microbial mat.
This wrinkled "elephant skin" texture is a trace fossil of a non-stromatolite microbial mat.
The image shows the location, in the Burgsvik beds of Sweden, where the texture was first identified as evidence of a microbial mat.[50]For about 2,000 million years microbial mats, multi-layered colonies of different types of bacteria, were the dominant life on Earth.[51] The evolution of oxygenic photosynthesis enabled them to play the major role in the oxygenation of the atmosphere[52] from about 2,400 million years ago. This change in the atmosphere increased their effectiveness as nurseries of evolution.[53] While eukaryotes, cells with complex internal structures, may have been present earlier, their evolution speeded up when they acquired the ability to transform oxygen from a poison to a powerful source of energy in their metabolism. This innovation may have come from primitive eukaryotes capturing oxygen-powered bacteria as endosymbionts and transforming them into organelles called mitochondria.[54] The earliest evidence of complex eukaryotes with organelles such as mitochondria, dates from 1,850 million years ago.[19]
Multicellular life is composed only of eukaryotic cells, and the earliest evidence for it is the Francevillian Group Fossils from 2,100 million years ago,[55] although specialization of cells for different functions first appears between 1,430 million years ago (a possible fungus) and 1,200 million years ago (a probable red alga). Sexual reproduction may be a prerequisite for specialization of cells, as an asexual multicellular organism might be at risk of being taken over by rogue cells that retain the ability to reproduce.[56][57]

The earliest known animals are cnidarians from about 580 million years ago, but these are so modern-looking that the earliest animals must have appeared before then.[58] Early fossils of animals are rare because they did not develop mineralized hard parts that fossilize easily until about 548 million years ago.[59] The earliest modern-looking bilaterian animals appear in the Early Cambrian, along with several "weird wonders" that bear little obvious resemblance to any modern animals. There is a long-running debate about whether this Cambrian explosion was truly a very rapid period of evolutionary experimentation; alternative views are that modern-looking animals began evolving earlier but fossils of their precursors have not yet been found, or that the "weird wonders" are evolutionary "aunts" and "cousins" of modern groups.[60] Vertebrates remained an obscure group until the first fish with jaws appeared in the Late Ordovician.[61][62]
The spread of life from water to land required organisms to solve several problems, including protection against drying out and supporting themselves against gravity.[63][64] The earliest evidence of land plants and land invertebrates date back to about 476 million years ago and 490 million years ago respectively.[64][65] The lineage that produced land vertebrates evolved later but very rapidly between 370 million years ago and 360 million years ago;[66] recent discoveries have overturned earlier ideas about the history and driving forces behind their evolution.[67] Land plants were so successful that they caused an ecological crisis in the Late Devonian, until the evolution and spread of fungi that could digest dead wood.[21]
 At about 13 centimetres (5.1 in) the Early Cretaceous Yanoconodon was longer than the average mammal of the time.[68]
At about 13 centimetres (5.1 in) the Early Cretaceous Yanoconodon was longer than the average mammal of the time.[68]During the Permian period synapsids, including the ancestors of mammals, may have dominated land environments,[70] but the Permian–Triassic extinction event 251 million years ago came very close to wiping out complex life.[71] During the slow recovery from this catastrophe a previously obscure group, archosaurs, became the most abundant and diverse terrestrial vertebrates. One archosaur group, the dinosaurs, were the dominant land vertebrates for the rest of the Mesozoic,[72] and birds evolved from one group of dinosaurs.[69] During this time mammals' ancestors survived only as small, mainly nocturnal insectivores, but this apparent set-back may have accelerated the development of mammalian traits such as endothermy and hair.[73] After the Cretaceous–Tertiary extinction event 65 million years ago killed off the non-avian dinosaurs – birds are the only surviving dinosaurs – mammals increased rapidly in size and diversity, and some took to the air and the sea.[74][75][76]
 A modern social insect collects pollen from a modern flowering plant.
A modern social insect collects pollen from a modern flowering plant.Fossil evidence indicates that flowering plants appeared and rapidly diversified in the Early Cretaceous, between 130 million years ago and 90 million years ago.[77] Their rapid rise to dominance of terrestrial ecosystems is thought to have been propelled by coevolution with pollinating insects.[78] Social insects appeared around the same time and, although they account for only small parts of the insect "family tree", now form over 50% of the total mass of all insects.[79]
Humans evolved from a lineage of upright-walking apes whose earliest fossils date from over 6 million years ago.[80] Although early members of this lineage had chimp-sized brains, about 25% as big as modern humans', there are signs of a steady increase in brain size after about 3 million years ago.[81] There is a long-running debate about whether modern humans are descendants of a single small population in Africa, which then migrated all over the world less than 200,000 years ago and replaced previous hominine species, or arose worldwide at the same time as a result of interbreeding.[82]
Mass extinctions
Apparent extinction intensity, i.e. the fraction of genera going extinct at any given time, as reconstructed from the fossil record (graph not meant to include recent epoch of Holocene extinction event)Main article: Mass extinctionLife on earth has suffered occasional mass extinctions at least since 542 million years ago. Although they are disasters at the time, mass extinctions have sometimes accelerated the evolution of life on earth. When dominance of particular ecological niches passes from one group of organisms to another, it is rarely because the new dominant group is "superior" to the old and usually because an extinction event eliminates the old dominant group and makes way for the new one.[83][84]
The fossil record appears to show that the rate of extinction is slowing down, with both the gaps between mass extinctions becoming longer and the average and background rates of extinction decreasing. However, it is not certain whether the actual rate of extinction has altered, since both of these observations could be explained in several ways:[85]
- The oceans may have become more hospitable to life over the last 500 million years and less vulnerable to mass extinctions: dissolved oxygen became more widespread and penetrated to greater depths; the development of life on land reduced the run-off of nutrients and hence the risk of eutrophication and anoxic events; marine ecosystems became more diversified so that food chains were less likely to be disrupted.[86][87]
- Reasonably complete fossils are very rare, most extinct organisms are represented only by partial fossils, and complete fossils are rarest in the oldest rocks. So paleontologists have mistakenly assigned parts of the same organism to different genera which were often defined solely to accommodate these finds – the story of Anomalocaris is an example of this.[88] The risk of this mistake is higher for older fossils because these are often unlike parts of any living organism. Many of the "superfluous" genera are represented by fragments which are not found again and the "superfluous" genera appear to become extinct very quickly.[85]
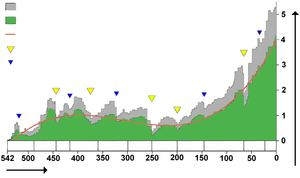 All genera"Well-defined" generaTrend line"Big Five" mass extinctionsOther mass extinctionsMillion years agoThousands of generaPhanerozoic biodiversity as shown by the fossil record
All genera"Well-defined" generaTrend line"Big Five" mass extinctionsOther mass extinctionsMillion years agoThousands of generaPhanerozoic biodiversity as shown by the fossil recordBiodiversity in the fossil record, which is
-
- "the number of distinct genera alive at any given time; that is, those whose first occurrence predates and whose last occurrence postdates that time"[89]
shows a different trend: a fairly swift rise from 542 to 400 million years ago, a slight decline from 400 to 200 million years ago, in which the devastating Permian–Triassic extinction event is an important factor, and a swift rise from 200 million years ago to the present.[89]
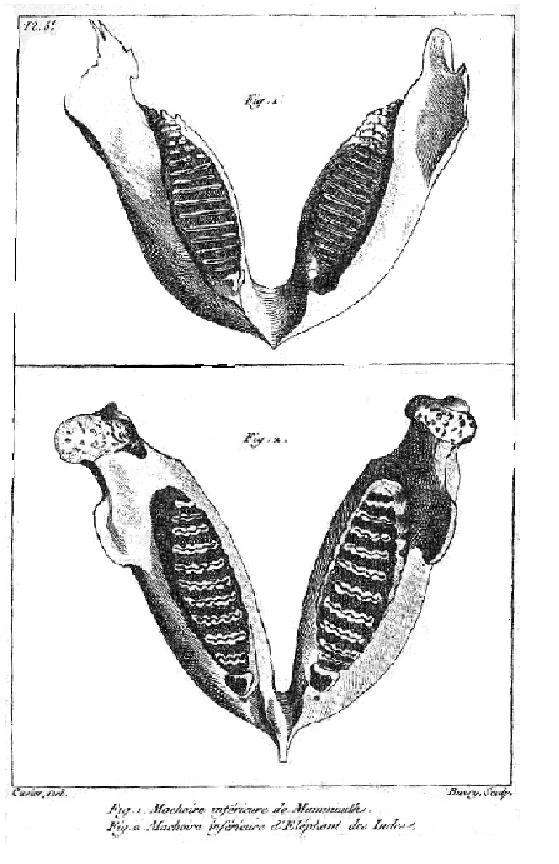
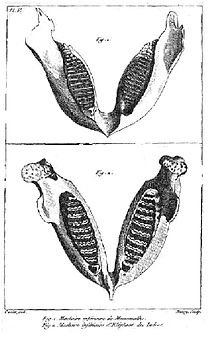 This illustration of an Indian elephant jaw and a mammoth jaw (top) is from Cuvier's 1796 paper on living and fossil elephants.
This illustration of an Indian elephant jaw and a mammoth jaw (top) is from Cuvier's 1796 paper on living and fossil elephants.History of paleontology
Main article: History of paleontologyAlthough paleontology became established around 1800, earlier thinkers had noticed aspects of the fossil record. The ancient Greek philosopher Xenophanes (570–480 BC) concluded from fossil sea shells that some areas of land were once under water.[90] During the Middle Ages the Persian naturalist Ibn Sina, known as Avicenna in Europe, discussed fossils and proposed a theory of petrifying fluids on which Albert of Saxony elaborated in the 14th century.[91] The Chinese naturalist Shen Kuo (1031–1095) proposed a theory of climate change based on the presence of petrified bamboo in regions that in his time were too dry for bamboo.[92]
In early modern Europe, the systematic study of fossils emerged as an integral part of the changes in natural philosophy that occurred during the Age of Reason. At the end of the 18th century Georges Cuvier's work established comparative anatomy as a scientific discipline and, by proving that some fossil animals resembled no living ones, demonstrated that animals could become extinct, leading to the emergence of paleontology.[93] The expanding knowledge of the fossil record also played an increasing role in the development of geology, particularly stratigraphy.[94]
The first half of the 19th century saw geological and paleontological activity become increasingly well organized with the growth of geologic societies and museums[95][96] and an increasing number of professional geologists and fossil specialists. Interest increased for reasons that were not purely scientific, as geology and paleontology helped industrialists to find and exploit natural resources such as coal.[97]
This contributed to a rapid increase in knowledge about the history of life on Earth and to progress in the definition of the geologic time scale, largely based on fossil evidence. In 1822 Henri Marie Ducrotay de Blanville, editor of Journal de Phisique, coined the word "paleontology" to refer to the study of ancient living organisms through fossils.[98] As knowledge of life's history continued to improve, it became increasingly obvious that there had been some kind of successive order to the development of life. This encouraged early evolutionary theories on the transmutation of species.[99] After Charles Darwin published Origin of Species in 1859, much of the focus of paleontology shifted to understanding evolutionary paths, including human evolution, and evolutionary theory.[99]
The last half of the 19th century saw a tremendous expansion in paleontological activity, especially in North America.[101] The trend continued in the 20th century with additional regions of the Earth being opened to systematic fossil collection. Fossils found in China near the end of the 20th century have been particularly important as they have provided new information about the earliest evolution of animals, early fish, dinosaurs and the evolution of birds.[102] The last few decades of the 20th century saw a renewed interest in mass extinctions and their role in the evolution of life on Earth.[103] There was also a renewed interest in the Cambrian explosion that apparently saw the development of the body plans of most animal phyla. The discovery of fossils of the Ediacaran biota and developments in paleobiology extended knowledge about the history of life back far before the Cambrian.[60]
Increasing awareness of Gregor Mendel's pioneering work in genetics led first to the development of population genetics and then in the mid-20th century to the modern evolutionary synthesis, which explains evolution as the outcome of events such as mutations and horizontal gene transfer which provide genetic variation, with genetic drift and natural selection driving changes in this variation over time.[104] Within the next few years the role and operation of DNA in genetic inheritance were discovered, leading to what is now known as the "Central Dogma" of molecular biology.[105] In the 1960s molecular phylogenetics, the investigation of evolutionary "family trees" by techniques derived from biochemistry, began to make an impact, particularly when it was proposed that the human lineage had diverged from apes much more recently than was generally thought at the time.[106] Although this early study compared proteins from apes and humans, most molecular phylogenetics research is now based on comparisons of RNA and DNA.[107]
See also
- Archaeobiology
- Dinosaurs
- Evolutionary history of life
- Fossil collecting
- Fossils
- Geology
- History of paleontology
- Important publications in paleontology
- List of fossil sites (with link directory)
- List of notable fossils
- List of transitional fossils
- Radiometric dating
- Taxonomy of commonly fossilised invertebrates
- Timeline of paleontology
- Treatise on Invertebrate Paleontology
Notes
References
- ^ a b Cowen, R. (2000). History of Life (3rd ed.). Blackwell Science. p. xi. ISBN 063204444-6.
- ^ Laporte, L.F. (October 1988). "What, after All, Is Paleontology?". PALAIOS 3 (5): 453. doi:10.2307/3514718. JSTOR 3514718.
- ^ a b Laudan, R. (1992). "What's so Special about the Past?". In Nitecki, M.H., and Nitecki, D.V.. History and Evolution. SUNY Press. p. 58. ISBN 0791412113. http://books.google.com/?id=kyLRtsvLS2AC&pg=PA55&dq=%22What%27s+so+Special+about+the+Past%22+laudan&q=%22What%27s%20so%20Special%20about%20the%20Past%22%20laudan. Retrieved 7 February 2010.
- ^ a b c Cleland, C.E. (September 2002). "Methodological and Epistemic Differences between Historical Science and Experimental Science" (PDF). Philosophy of Science 69: 474–496. doi:10.1086/342453. http://spot.colorado.edu/~cleland/articles/Cleland.PS.Pdf. Retrieved September 17, 2008.
- ^ McGraw-Hill Encyclopedia of Science & Technology. McGraw-Hill. 2002. p. 58. ISBN 0079136656.
- ^ Laudan, R. (1992). "What's so Special about the Past?". In Nitecki, M.H., and Nitecki, D.V.. History and Evolution. SUNY Press. p. 57. ISBN 0791412113.
- ^ "How does paleontology differ from anthropology and archaeology?". University of California Museum of Paleontology. http://www.ucmp.berkeley.edu/faq.php#anthro. Retrieved September 17, 2008.
- ^ a b c Brasier, M., McLoughlin, N., Green, O., and Wacey, D. (June 2006). "A fresh look at the fossil evidence for early Archaean cellular life" (PDF). Philosophical Transactions of the Royal Society: Biology 361 (1470): 887–902. doi:10.1098/rstb.2006.1835. PMC 1578727. PMID 16754605. http://physwww.mcmaster.ca/~higgsp/3D03/BrasierArchaeanFossils.pdf. Retrieved August 30, 2008.
- ^ a b Twitchett RJ, Looy CV, Morante R, Visscher H, Wignall PB (2001). "Rapid and synchronous collapse of marine and terrestrial ecosystems during the end-Permian biotic crisis". Geology 29 (4): 351–354. Bibcode 2001Geo....29..351T. doi:10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2.
- ^ a b Peterson, Kevin J., and Butterfield, N.J. (2005). "Origin of the Eumetazoa: Testing ecological predictions of molecular clocks against the Proterozoic fossil record". Proceedings of the National Academy of Sciences 102 (27): 9547–52. Bibcode 2005PNAS..102.9547P. doi:10.1073/pnas.0503660102. PMC 1172262. PMID 15983372. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1172262.
- ^ Hutchinson, John R.; Garcia, M (28 February 2002). "Tyrannosaurus was not a fast runner". Nature 415 (6875): 1018–1021. doi:10.1038/4151018a. PMID 11875567. http://www.nature.com/nature/journal/v415/n6875/full/4151018a.html. Summary in press release No Olympian: Analysis hints T. rex ran slowly, if at all
- ^ Meers, M.B. (August 2003). "Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior". Historical Biology: A Journal of Paleobiology 16 (1): 1–12. doi:10.1080/0891296021000050755. http://www.ingentaconnect.com/content/tandf/ghbi/2003/00000016/00000001/art00001.
- ^ Bruner, Emiliano (November 2004). "Geometric morphometrics and paleoneurology: brain shape evolution in the genus Homo". Journal of Human Evolution 47 (5): 279–303. doi:10.1016/j.jhevol.2004.03.009. http://www.sciencedirect.com/science/article/pii/S0047248404001149. Retrieved 27 September 2011.
- ^ Cady, S.L. (April 1998). "Astrobiology: A New Frontier for 21st Century Paleontologists". PALAIOS 13 (2): 95–97. doi:10.2307/3515482. JSTOR 3515482. PMID 11542813.
- ^ Plotnick, R.E.. "A Somewhat Fuzzy Snapshot of Employment in Paleontology in the United States". Palaeontologia Electronica (Coquina Press) 11 (1). ISSN 1094-8074. http://palaeo-electronica.org/2008_1/commentary/comment.htm. Retrieved September 17, 2008.
- ^ a b "What is Paleontology?". University of California Museum of Paleontology. http://www.ucmp.berkeley.edu/FAQ/whatispaleo.html. Retrieved September 17, 2008.
- ^ Kitchell, J.A. (1985). "Evolutionary Paleocology: Recent Contributions to Evolutionary Theory". Paleobiology 11 (1): 91–104. http://www.ucmp.berkeley.edu/FAQ/whatispaleo.html. Retrieved September 17, 2008.
- ^ Hoehler, T.M., Bebout, B.M., and Des Marais, D.J. (19 July 2001). "The role of microbial mats in the production of reduced gases on the early Earth". Nature 412 (6844): 324–327. doi:10.1038/35085554. PMID 11460161. http://www.nature.com/nature/journal/v412/n6844/full/412324a0.html. Retrieved July 14, 2008.
- ^ a b Hedges, S.B., Blair, J.E, Venturi, M.L., and Shoe, J.L. (January 2004). "A molecular timescale of eukaryote evolution and the rise of complex multicellular life". BMC Evolutionary Biology 4 (2): 2. doi:10.1186/1471-2148-4-2. PMC 341452. PMID 15005799. http://www.biomedcentral.com/1471-2148/4/2/abstract/. Retrieved July 14, 2008.
- ^ "Paleoclimatology". Ohio State University. http://matrix.geology.ohio-state.edu/ProspectiveGradStudents/school-of-earth-sciences-directory/specialties/global-and-environmental-change/paleoclimatology. Retrieved September 17, 2008.
- ^ a b Algeo, T.J., and Scheckler, S.E. (1998). "Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events". Philosophical Transactions of the Royal Society: Biology 353: 113–130. doi:10.1098/rstb.1998.0195. PMC 1692181. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1692181. Retrieved September 5, 2008.
- ^ "Biostratigraphy: William Smith". http://evolution.berkeley.edu/evosite/history/biostratigraphy.shtml. Retrieved September 17, 2008.
- ^ "Biogeography: Wallace and Wegener (1 of 2)". University of California Museum of Paleontology and University of California at Berkeley. http://evolution.berkeley.edu/evosite/history/biogeography.shtml. Retrieved September 17, 2008.
- ^ a b "What is paleontology?". University of California Museum of Paleontology. http://www.ucmp.berkeley.edu/faq.php#paleo. Retrieved September 17, 2008.
- ^ Benton MJ, Wills MA, Hitchin R (2000). "Quality of the fossil record through time". Nature 403 (6769): 534–7. Bibcode 2000Natur.403..534B. doi:10.1038/35000558. PMID 10676959.
- Non-technical summary
- ^ Butterfield, N.J. (2003). "Exceptional Fossil Preservation and the Cambrian Explosion". Integrative and Comparative Biology 43 (1): 166–177. doi:10.1093/icb/43.1.166. PMID 21680421. http://icb.oxfordjournals.org/cgi/content/abstract/43/1/166. Retrieved June 28, 2008.
- ^ Cowen, R. (2000). History of Life (3rd ed.). Blackwell Science. p. 61. ISBN 063204444-6.
- ^ Butterfield, N.J. (2001). "Ecology and evolution of Cambrian plankton". The Ecology of the Cambrian Radiation (New York: Columbia University Press): 200–216. http://66.102.1.104/scholar?hl=en&lr=&ie=UTF-8&q=cache:9xeRu1SdF0QJ:www.earthscape.org/r3/ES14785/ch09.pdf+. Retrieved September 27, 2007.
- ^ Signor, P.W. (1982). "Sampling bias, gradual extinction patterns and catastrophes in the fossil record". Geological implications of impacts of large asteroids and comets on the earth (Boulder, CO: Geological Society of America): 291–296. A 84–25651 10–42. http://www.csa.com/partners/viewrecord.php?requester=gs&collection=TRD&recid=A8425675AH. Retrieved January 1, 2008.
- ^ a b Fedonkin, M.A., Gehling, J.G., Grey, K., Narbonne, G.M., Vickers-Rich, P. (2007). The Rise of Animals: Evolution and Diversification of the Kingdom Animalia. JHU Press. pp. 213–216. ISBN 0801886791. http://books.google.com/?id=OFKG6SmPNuUC&pg=PA213&lpg=PA213&dq=trace+fossil+complex+animal. Retrieved November 14, 2008.
- ^ e.g. Seilacher, A. (1994). "How valid is Cruziana Stratigraphy?" (PDF). International Journal of Earth Sciences 83 (4): 752–758. http://www.springerlink.com/index/WP279834395100KH.pdf. Retrieved September 9, 2007.
- ^ Brocks, J.J., Logan, G.A., Buick, R., and Summons, R.E. (1999). "Archaean molecular fossils and the rise of eukaryotes". Science 285 (5430): 1033–1036. doi:10.1126/science.285.5430.1033. PMID 10446042. http://www.sciencemag.org/cgi/content/abstract/285/5430/1033. Retrieved September 2, 2008.
- ^ a b Cowen, R. (2000). History of Life (3rd ed.). Blackwell Science. pp. 47–50. ISBN 063204444-6.
- ^ a b Brochu, C.A, and Sumrall, C.D. (July 2001). "Phylogenetic Nomenclature and Paleontology". Journal of Paleontology 75 (4): 754–757. doi:10.1666/0022-3360(2001)075<0754:PNAP>2.0.CO;2. JSTOR 1306999.
- ^ Ereshefsky, M. (2001). The Poverty of the Linnaean Hierarchy: A Philosophical Study of Biological Taxonomy. Cambridge University Press. p. 5. ISBN 0521781701. http://books.google.com/?id=tM6E8-_vSD0C&dq=%22The+Poverty+of+the+Linnaean+Hierarchy:+A+Philosophical+Study+of+Biological+Taxonomy%22&pg=PP1.
- ^ Cohen, B. L. and Holmer, L. E. and Luter, C. (2003). "The brachiopod fold: a neglected body plan hypothesis" (PDF). Palaeontology 46 (1): 59–65. doi:10.1111/1475-4983.00287. http://eprints.gla.ac.uk/2920/01/Cohen_2920.pdf. Retrieved August 7, 2008.
- ^ a b Martin, M.W.; Grazhdankin, D.V.; Bowring, S.A.; Evans, D.A.D.; Fedonkin, M.A.; Kirschvink, J.L. (May 5, 2000). "Age of Neoproterozoic Bilaterian Body and Trace Fossils, White Sea, Russia: Implications for Metazoan Evolution" (abstract). Science 288 (5467): 841–5. Bibcode 2000Sci...288..841M. doi:10.1126/science.288.5467.841. PMID 10797002.
- ^ Pufahl, P.K., Grimm, K.A., Abed, A.M., and Sadaqah, R.M.Y. (October 2003). "Upper Cretaceous (Campanian) phosphorites in Jordan: implications for the formation of a south Tethyan phosphorite giant". Sedimentary Geology 161 (3-4): 175–205. Bibcode 2003SedG..161..175P. doi:10.1016/S0037-0738(03)00070-8.
- ^ "Geologic Time: Radiometric Time Scale". U.S. Geological Survey. http://pubs.usgs.gov/gip/geotime/radiometric.html. Retrieved September 20, 2008.
- ^ Löfgren, A. (2004). "The conodont fauna in the Middle Ordovician Eoplacognathus pseudoplanus Zone of Baltoscandia". Geological Magazine 141 (04): 505–524. doi:10.1017/S0016756804009227. http://journals.cambridge.org/production/action/cjoGetFulltext?fulltextid=235488. Retrieved November 17, 2008.
- ^ Gehling, James; Jensen, Sören; Droser, Mary; Myrow, Paul; Narbonne, Guy (March 2001). "Burrowing below the basal Cambrian GSSP, Fortune Head, Newfoundland". Geological Magazine 138 (2): 213–218. doi:10.1017/S001675680100509X. http://www.journals.cambridge.org/action/displayAbstract?fromPage=online&aid=74669. Retrieved November 17, 2008.
- ^ e.g. Gehling, James; Jensen, Sören; Droser, Mary; Myrow, Paul; Narbonne, Guy (March 2001). "Burrowing below the basal Cambrian GSSP, Fortune Head, Newfoundland". Geological Magazine 138 (2): 213–218. doi:10.1017/S001675680100509X. http://www.journals.cambridge.org/action/displayAbstract?fromPage=online&aid=74669. Retrieved November 17, 2008.
- ^ Hug, L.A., and Roger, A.J. (2007). "The Impact of Fossils and Taxon Sampling on Ancient Molecular Dating Analyses". Molecular Biology and Evolution 24 (8): 889–1897. doi:10.1093/molbev/msm115. PMID 17556757.
- ^ * "Early Earth Likely Had Continents And Was Habitable". 2005-11-17. http://www.colorado.edu/news/releases/2005/438.html.
* Cavosie, A. J.; J. W. Valley, S. A., Wilde, and E.I.M.F. (July 15, 2005). "Magmatic δ18O in 4400-3900 Ma detrital zircons: A record of the alteration and recycling of crust in the Early Archean". Earth and Planetary Science Letters 235 (3-4): 663–681. Bibcode 2005E&PSL.235..663C. doi:10.1016/j.epsl.2005.04.028. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6V61-4GDKB05-3&_coverDate=07%2F15%2F2005&_alid=382434001&_rdoc=1&_fmt=&_orig=search&_qd=1&_cdi=5801&_sort=d&view=c&_acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=be47e49c535d059be188b66c6e596dd5. - ^ Dauphas, N., Robert, F., and Marty, B. (December 2000). "The Late Asteroidal and Cometary Bombardment of Earth as Recorded in Water Deuterium to Protium Ratio". Icarus 148 (2): 508–512. Bibcode 2000Icar..148..508D. doi:10.1006/icar.2000.6489.
- ^ Schopf, J. (2006). "Fossil evidence of Archaean life". Philos Trans R Soc Lond B Biol Sci 361 (1470): 869–85. doi:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1578735.
- ^ Cowen, R. (2000). History of Life (3rd ed.). Blackwell Science. p. 6. ISBN 0632044446.
- ^ * Arrhenius, S. (1903). "The Propagation of Life in Space". Die Umschau volume=7. Reprinted in Goldsmith, D.,, ed. The Quest for Extraterrestrial Life. University Science Books. ISBN 0198557043.
* Hoyle, F., and Wickramasinghe, C. (1979). "On the Nature of Interstellar Grains". Astrophysics and Space Science 66: 77–90. Bibcode 1979Ap&SS..66...77H. doi:10.1007/BF00648361.
* Crick, F. H.; Orgel, L. E. (1973). "Directed Panspermia". Icarus 19: 341–348. Bibcode 1973Icar...19..341C. doi:10.1016/0019-1035(73)90110-3. - ^ Peretó, J. (2005). "Controversies on the origin of life" (PDF). Int. Microbiol. 8 (1): 23–31. PMID 15906258. http://www.im.microbios.org/0801/0801023.pdf. Retrieved October 7, 2007.
- ^ Manten, A.A. (1966). "Some problematic shallow-marine structures". Marine Geol 4: 227–232. doi:10.1016/0025-3227(66)90023-5. http://igitur-archive.library.uu.nl/geo/2006-1213-210452/UUindex.html. Retrieved June 18, 2007.
- ^ Krumbein, W.E., Brehm, U., Gerdes, G., Gorbushina, A.A., Levit, G. and Palinska, K.A. (2003). "Biofilm, Biodictyon, Biomat Microbialites, Oolites, Stromatolites, Geophysiology, Global Mechanism, Parahistology". In Krumbein, W.E., Paterson, D.M., and Zavarzin, G.A.. Fossil and Recent Biofilms: A Natural History of Life on Earth. Kluwer Academic. pp. 1–28. ISBN 1402015976. http://134.106.242.33/krumbein/htdocs/Archive/397/Krumbein_397.pdf. Retrieved July 9, 2008.
- ^ Hoehler, T.M., Bebout, B.M., and Des Marais, D.J. (July 19, 2001). "The role of microbial mats in the production of reduced gases on the early Earth". Nature 412 (6844): 324–327. doi:10.1038/35085554. PMID 11460161. http://www.nature.com/nature/journal/v412/n6844/full/412324a0.html. Retrieved July 14, 2008.
- ^ Nisbet, E.G., and Fowler, C.M.R. (December 7, 1999). "Archaean metabolic evolution of microbial mats" (PDF). Proceedings of the Royal Society: Biology 266 (1436): 2375. doi:10.1098/rspb.1999.0934. PMC 1690475. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1690475. Retrieved July 16, 2008.
- ^ Gray MW, Burger G, Lang BF (March 1999). "Mitochondrial evolution". Science 283 (5407): 1476–81. Bibcode 1999Sci...283.1476G. doi:10.1126/science.283.5407.1476. PMID 10066161.
- ^ El Albani, Abderrazak; Bengtson, Stefan; Canfield, Donald E.; Bekker, Andrey; Macchiarelli, Reberto (July 2010). "Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago". Nature 466 (7302): 100–104. Bibcode 2010Natur.466..100A. doi:10.1038/nature09166. PMID 20596019.
- ^ Butterfield, N.J. (September 2000). "Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes". Paleobiology 26 (3): 386–404. doi:10.1666/0094-8373(2000)026<0386:BPNGNS>2.0.CO;2. http://paleobiol.geoscienceworld.org/cgi/content/abstract/26/3/386. Retrieved 2008-09-02.
- ^ Butterfield, N.J. (2005). "Probable Proterozoic fungi". Paleobiology 31 (1): 165–182. doi:10.1666/0094-8373(2005)031<0165:PPF>2.0.CO;2. http://paleobiol.geoscienceworld.org/cgi/content/abstract/31/1/165. Retrieved 2008-09-02.
- ^ Chen, J.-Y., Oliveri, P., Gao, F., Dornbos, S.Q., Li, C-W., Bottjer, D.J. and Davidson, E.H. (August 2002). "Precambrian Animal Life: Probable Developmental and Adult Cnidarian Forms from Southwest China" (PDF). Developmental Biology 248 (1): 182–196. doi:10.1006/dbio.2002.0714. PMID 12142030. http://www.uwm.edu/~sdornbos/PDF's/Chen%20et%20al.%202002.pdf. Retrieved September 3, 2008.
- ^ Bengtson, S. (2004). Early skeletal fossils. In Lipps, J.H., and Waggoner, B.M.. "Neoproterozoic — Cambrian Biological Revolutions". Paleontological Society Papers 10: 67–78. http://www.cosmonova.org/download/18.4e32c81078a8d9249800021554/Bengtson2004ESF.pdf. Retrieved July 18, 2008.
- ^ a b Marshall, C.R. (2006). "Explaining the Cambrian "Explosion" of Animals". Annu. Rev. Earth Planet. Sci. 34: 355–384. Bibcode 2006AREPS..34..355M. doi:10.1146/annurev.earth.33.031504.103001. http://arjournals.annualreviews.org/doi/abs/10.1146/annurev.earth.33.031504.103001?journalCode=earth. Retrieved November 6, 2007.
- ^ Conway Morris, S. (August 2, 2003). "Once we were worms". New Scientist 179 (2406): 34. http://cas.bellarmine.edu/tietjen/Evolution/once_we_were_worms.htm. Retrieved September 5, 2008.
- ^ Sansom I.J., Smith, M.M., and Smith, M.P. (2001). "The Ordovician radiation of vertebrates". In Ahlberg, P.E.. Major Events in Early Vertebrate Evolution. Taylor and Francis. pp. 156–171. ISBN 0-415-23370-4.
- ^ Selden, P.A. (2001). ""Terrestrialization of Animals"". In Briggs, D.E.G., and Crowther, P.R.. Palaeobiology II: A Synthesis. Blackwell. pp. 71–74. ISBN 0632051493. http://books.google.com/?id=AHsrhGOTRM4C&pg=PA71&lpg=PA71&dq=%22Terrestrialization+of+Animals%22+selden. Retrieved September 5, 2008.
- ^ a b Kenrick, P., and Crane, P.R. (September 1997). "The origin and early evolution of plants on land" (PDF). Nature 389 (6646): 33. Bibcode 1997Natur.389...33K. doi:10.1038/37918. http://blog.richmond.edu/paleontology/files/2008/03/nature-evol-plants-kendrickcrane1997.pdf. Retrieved 2010-11-10.
- ^ MacNaughton, R.B., Cole, J.M., Dalrymple, R.W., Braddy, S.J., Briggs, D.E.G., and Lukie, T.D. (May 2002). "First steps on land: Arthropod trackways in Cambrian-Ordovician eolian sandstone, southeastern Ontario, Canada". Geology 30 (5): 391–394. Bibcode 2002Geo....30..391M. doi:10.1130/0091-7613(2002)030<0391:FSOLAT>2.0.CO;2. http://geology.geoscienceworld.org/cgi/content/abstract/30/5/391. Retrieved 2008-09-05.
- ^ Gordon, M.S, Graham, J.B., and Wang, T. (September/October 2004). "Revisiting the Vertebrate Invasion of the Land". Physiological and Biochemical Zoology 77 (5): 697–699. doi:10.1086/425182.
- ^ Clack, J.A. (November 2005). "Getting a Leg Up on Land". Scientific American. http://www.sciam.com/article.cfm?id=getting-a-leg-up-on-land. Retrieved September 6, 2008.
- ^ Luo, Z., Chen, P., Li, G., & Chen, M. (March 2007). "A new eutriconodont mammal and evolutionary development in early mammals". Nature 446 (7133): 288–293. Bibcode 2007Natur.446..288L. doi:10.1038/nature05627. PMID 17361176.
- ^ a b Padian, Kevin. (2004). "Basal Avialae". In Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. ISBN 0-520-24209-2.
- ^ Sidor, C.A., O'Keefe, F.R., Damiani, R., Steyer, J.S., Smith, R.M.H., Larsson, H.C.E., Sereno, P.C., Ide, O, and Maga, A. (April 2005). "Permian tetrapods from the Sahara show climate-controlled endemism in Pangaea". Nature 434 (7035): 886–889. Bibcode 2005Natur.434..886S. doi:10.1038/nature03393. PMID 15829962. http://www.nature.com/nature/journal/v434/n7035/full/nature03393.html. Retrieved September 8, 2008.
- ^ Benton M.J. (2005). When Life Nearly Died: The Greatest Mass Extinction of All Time. Thames & Hudson. ISBN 978-0500285732.
- ^ Benton, M.J. (March 1983). "Dinosaur Success in the Triassic: a Noncompetitive Ecological Model". Quarterly Review of Biology 58 (1). http://palaeo.gly.bris.ac.uk/Benton/reprints/1983success.pdf. Retrieved September 8, 2008.
- ^ Ruben, J.A., and Jones, T.D. (2000). "Selective Factors Associated with the Origin of Fur and Feathers". American Zoologist 40 (4): 585–596. doi:10.1093/icb/40.4.585. http://icb.oxfordjournals.org/cgi/content/full/40/4/585.
- ^ Alroy J. (March 1999). "The fossil record of North American mammals: evidence for a Paleocene evolutionary radiation". Systematic biology 48 (1): 107–18. doi:10.1080/106351599260472. PMID 12078635.
- ^ Simmons, N.B., Seymour,K.L., Habersetzer, J.,and Gunnell, G.F. (February 2008). "Primitive Early Eocene bat from Wyoming and the evolution of flight and echolocation". Nature 451 (7180): 818–821. Bibcode 2008Natur.451..818S. doi:10.1038/nature06549. PMID 18270539.
- ^ J. G. M. Thewissen, S. I. Madar, and S. T. Hussain (1996). "Ambulocetus natans, an Eocene cetacean (Mammalia) from Pakistan". Courier Forschungsinstitut Senckenberg 191: 1–86.
- ^ Crane, P.R., Friis, E.M., and Pedersen, K.R. (2000). "The Origin and Early Diversification of Angiosperms". In Gee, H.. Shaking the Tree: Readings from Nature in the History of Life. University of Chicago Press. pp. 233–250. ISBN 0226284964. http://books.google.com/?id=ZJe_Dmdbm-QC&pg=PA233&dq=evolution+flowering+plant+angiosperm. Retrieved September 9, 2008.
- ^ Crepet, W.L. (November 2000). "Progress in understanding angiosperm history, success, and relationships: Darwin’s abominably "perplexing phenomenon"". Proceedings of the National Academy of Sciences 97 (24): 12939–12941. Bibcode 2000PNAS...9712939C. doi:10.1073/pnas.97.24.12939. PMC 34068. PMID 11087846. http://www.pnas.org/content/97/24/12939.full.pdf+html. Retrieved September 9 , 2008.
- ^ Brunet M., Guy, F., Pilbeam, D., Mackaye, H.T., et al. (July 2002). "A new hominid from the Upper Miocene of Chad, Central Africa". Nature 418 (6894): 145–151. doi:10.1038/nature00879. PMID 12110880. http://www.nature.com/nature/journal/v418/n6894/abs/nature00879.html. Retrieved September 9, 2008.
- ^ De Miguel, C., and M. Henneberg, M. (2001). "Variation in hominid brain size: How much is due to method?". HOMO — Journal of Comparative Human Biology 52 (1): 3–58. doi:10.1078/0018-442X-00019. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B7GW4-4DPCHXC-2&_user=10&_rdoc=1&_fmt=&_orig=search&_sort=d&view=c&_version=1&_urlVersion=0&_userid=10&md5=aef79dbca1f189c885cfe9f36636b131. Retrieved September 9, 2008.
- ^ Leakey, Richard (1994). The Origin of Humankind. Science Masters Series. New York, NY: Basic Books. pp. 87–89. ISBN 0465053130.
- ^ Benton, M.J. (2004). "6. Reptiles Of The Triassic". Vertebrate Palaeontology. Blackwell. ISBN 0045660026. http://www.blackwellpublishing.com/book.asp?ref=0632056371. Retrieved November 17, 2008.
- ^ Van Valkenburgh, B. (1999). "Major patterns in the history of xarnivorous mammals". Annual Review of Earth and Planetary Sciences 26: 463–493. Bibcode 1999AREPS..27..463V. doi:10.1146/annurev.earth.27.1.463. http://arjournals.annualreviews.org/doi/abs/10.1146/annurev.earth.27.1.463.
- ^ a b MacLeod, Norman (2001-01-06). "Extinction!". http://www.firstscience.com/home/articles/earth/extinction-page-2-1_1258.html. Retrieved September 11, 2008.
- ^ Martin, R.E. (1995). "Cyclic and secular variation in microfossil biomineralization: clues to the biogeochemical evolution of Phanerozoic oceans". Global and Planetary Change 11 (1): 1. Bibcode 1995GPC....11....1M. doi:10.1016/0921-8181(94)00011-2.
- ^ Martin, R.E. (1996). "Secular increase in nutrient levels through the Phanerozoic: Implications for productivity, biomass, and diversity of the marine biosphere". PALAIOS 11 (3): 209–219. doi:10.2307/3515230. JSTOR 3515230.
- ^ Gould, S.J. (1990). Wonderful Life: The Burgess Shale and the Nature of History. Hutchinson Radius. pp. 194–206. ISBN 0091742714.
- ^ a b Rohde, R.A., and Muller, R.A. (March 2005). "Cycles in fossil diversity" (PDF). Nature 434 (7030): 208–210. Bibcode 2005Natur.434..208R. doi:10.1038/nature03339. PMID 15758998. http://muller.lbl.gov/papers/Rohde-Muller-Nature.pdf. Retrieved September 22, 2008.
- ^ Rudwick, Martin J.S. (1985). The Meaning of Fossils (2nd ed.). The University of Chicago Press. p. 39. ISBN 0-226-73103-0.
- ^ Rudwick, Martin J.S. (1985). The Meaning of Fossils (2nd ed.). The University of Chicago Press. p. 24. ISBN 0-226-73103-0.
- ^ Needham, Joseph (1986). Science and Civilization in China: Volume 3, Mathematics and the Sciences of the Heavens and the Earth. Caves Books Ltd. p. 614. ISBN 0-253-34547-2.
- ^ McGowan, Christopher (2001). The Dragon Seekers. Persus Publishing. pp. 3–4. ISBN 0-7382-0282-7.
- ^ Palmer, D. (2005). Earth Time: Exploring the Deep Past from Victorian England to the Grand Canyon. Wiley. ISBN 780470022214.
- ^ Greene, Marjorie; David Depew (2004). The Philosophy of Biology: An Episodic History. Cambridge University Press. pp. 128–130. ISBN 0-521-64371-6.
- ^ Bowler, Peter J.; Iwan Rhys Morus (2005). Making Modern Science. The University of Chicago Press. pp. 168–169. ISBN 0-226-06861-7.
- ^ Rudwick, Martin J.S. (1985). The Meaning of Fossils (2nd ed.). The University of Chicago Press. pp. 200–201. ISBN 0-226-73103-0.
- ^ Rudwick, Martin J.S. (2008). Worlds Before Adam: The Reconstruction of Geohistory in the Age of Reform. The University of Chicago Press. p. 48. ISBN 0-226-73128-6.
- ^ a b Buckland W & Gould SJ (1980). Geology and Mineralogy Considered With Reference to Natural Theology (History of Paleontology). Ayer Company Publishing. ISBN 978-0405127069.
- ^ Shu, D-G., Conway Morris, S., Han, J., et al. (January 2003). "Head and backbone of the Early Cambrian vertebrate Haikouichthys". Nature 421 (6922): 526–529. Bibcode 2003Natur.421..526S. doi:10.1038/nature01264. PMID 12556891. http://www.nature.com/nature/journal/v421/n6922/abs/nature01264.html. Retrieved September 21, 2008.
- ^ Everhart, Michael J. (2005). Oceans of Kansas: A Natural History of the Western Interior Sea. Indiana University Press. p. 17. ISBN 0-253-34547-2.
- ^ Gee, H., ed (2001). Rise of the Dragon: Readings from Nature on the Chinese Fossil Record. Chicago, Ill. ;London: University of Chicago Press. pp. 276. ISBN 0226284913. http://books.google.com/?id=6WYZQwmyWy0C&dq=Rise+of+the+Dragon++By+Henry+Gee&printsec=frontcover. Retrieved September 21, 2008.
- ^ Bowler, Peter J. (2003). Evolution:The History of an Idea. University of California Press. pp. 351–352. ISBN 0-52023693-9.
- ^ Bowler, Peter J. (2003). Evolution:The History of an Idea. University of California Press. pp. 325–339. ISBN 0-52023693-9.
- ^ Crick, F.H.C. (1955). "On degenerate templates and the adaptor hypothesis" (PDF). http://genome.wellcome.ac.uk/assets/wtx030893.pdf. Retrieved October 4, 2008.
- ^ Sarich V.M., and Wilson A.C. (December 1967). "Immunological time scale for hominid evolution". Science 158 (3805): 1200–1203. Bibcode 1967Sci...158.1200S. doi:10.1126/science.158.3805.1200. PMID 4964406. http://www.sciencemag.org/cgi/content/abstract/158/3805/1200. Retrieved September 21, 2008.
- ^ Page, R.D.M, and Holmes, E.C. (1998). Molecular Evolution: A Phylogenetic Approach. Oxford: Blackwell Science. p. 2. ISBN 0865428891.
External links
- Smithsonian's Paleobiology website: a good introduction
- University of California Museum of Paleontology FAQ About Paleontology
- The Paleontological Society
- The Palaeontological Association
- The Paleontology Portal
- International Palaeoentomological Society
Branches of Biology Anatomy · Astrobiology · Biochemistry · Biogeography · Biomechanics · Biophysics · Bioinformatics · Biostatistics · Botany · Cell biology · Cellular microbiology · Chemical biology · Chronobiology · Conservation biology · Developmental biology · Ecology · Epidemiology · Epigenetics · Evolutionary biology · Genetics · Genomics · Histology · Human biology · Immunology · Marine biology · Mathematical biology · Microbiology · Molecular biology · Mycology · Neuroscience · Nutrition · Origin of life · Paleontology · Parasitology · Pathology · Pharmacology · Physiology · Quantum biology · Systematics · Systems biology · Taxonomy · Toxicology · ZoologyCategories:- Earth sciences
- Geology
- Paleontology
- Zoology
- Fossils
- Historical geology











Wikimedia Foundation. 2010.