- Visual cortex
-
Brain: Visual cortex 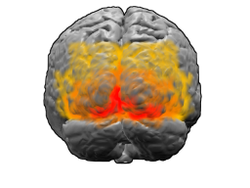
View of the brain from behind. Red = Brodmann area 17 (primary visual cortex); orange = area 18; yellow = area 19 
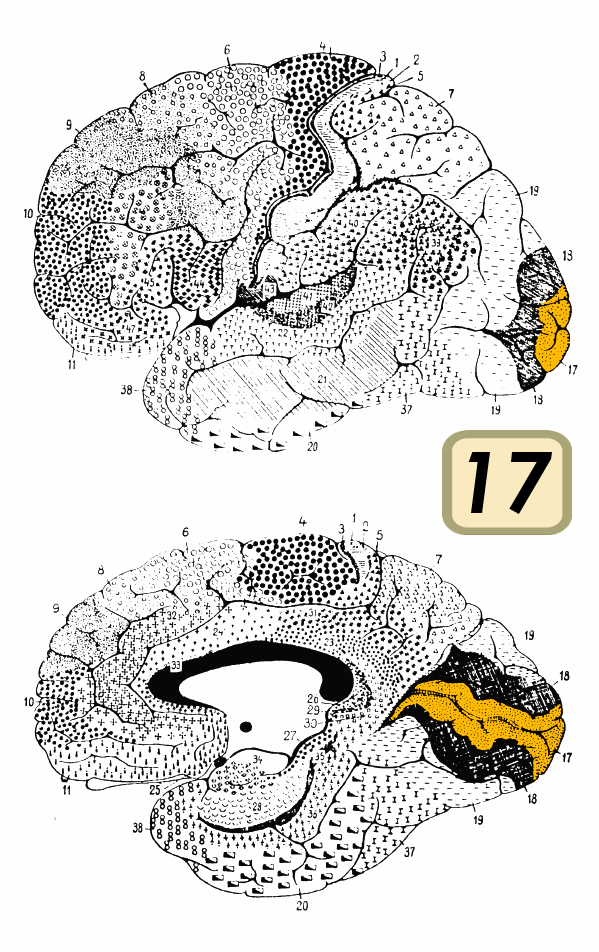
Brain shown from the side, facing left. Above: view from outside, below: cut through the middle. Orange = Brodmann area 17 (primary visual cortex) Latin Cortex visualis  The dorsal stream (green) and ventral stream (purple) are shown. They originate from primary visual cortex.
The dorsal stream (green) and ventral stream (purple) are shown. They originate from primary visual cortex.
The visual cortex of the brain is the part of the cerebral cortex responsible for processing visual information. It is located in the occipital lobe, in the back of the brain.
The term visual cortex refers to the primary visual cortex (also known as striate cortex or V1) and extrastriate visual cortical areas such as V2, V3, V4, and V5. The primary visual cortex is anatomically equivalent to Brodmann area 17, or BA17. The extrastriate cortical areas consist of Brodmann area 18 and Brodmann area 19.
There is a visual cortex in each hemisphere of the brain. The left hemisphere visual cortex receives signals from the right visual field and the right visual cortex from the left visual field.
The body of this article describes the primate (especially, human) visual cortex.
Contents
Introduction
The primary visual cortex, V1, is the koniocortex (sensory type) located in and around the calcarine fissure in the occipital lobe. Each hemisphere's V1 receives information directly from its ipsilateral lateral geniculate nucleus.
Each V1 transmits information to two primary pathways, called the dorsal stream and the ventral stream:
- The dorsal stream begins with V1, goes through Visual area V2, then to the dorsomedial area and Visual area MT (also known as V5) and to the posterior parietal cortex. The dorsal stream, sometimes called the "Where Pathway" or "How Pathway", is associated with motion, representation of object locations, and control of the eyes and arms, especially when visual information is used to guide saccades or reaching.[1]
- The ventral stream begins with V1, goes through visual area V2, then through visual area V4, and to the inferior temporal cortex. The ventral stream, sometimes called the "What Pathway", is associated with form recognition and object representation. It is also associated with storage of long-term memory.
The dichotomy of the dorsal/ventral pathways (also called the "where/what" or "action/perception" streams) [1] was first defined by Ungerleider and Mishkin [2] and is still contentious among vision scientists and psychologists. It is probably an over-simplification of the true state of affairs in the visual cortex. It is based on the findings that visual illusions such as the Ebbinghaus illusion may distort judgements of a perceptual nature, but when the subject responds with an action, such as grasping, no distortion occurs. However, recent work [3] suggests that both the action and perception systems are equally fooled by such illusions.
Neurons in the visual cortex fire action potentials when visual stimuli appear within their receptive field. By definition, the receptive field is the region within the entire visual field which elicits an action potential. But for any given neuron, it may respond best to a subset of stimuli within its receptive field. This property is called neuronal tuning. In the earlier visual areas, neurons have simpler tuning. For example, a neuron in V1 may fire to any vertical stimulus in its receptive field. In the higher visual areas, neurons have complex tuning. For example, in the inferior temporal cortex (IT), a neuron may only fire when a certain face appears in its receptive field.
The visual cortex receives its blood supply primarily from the calcarine branch of the posterior cerebral artery.
Current research
Research on the primary visual cortex can involve recording action potentials from electrodes within the brain of cats, ferrets, rats, mice, or monkeys, or through recording intrinsic optical signals from animals or EEG, MEG, or fMRI signals from human and monkey V1.
One recent discovery concerning the human V1 is that signals measured by fMRI show very large attentional modulation. This result strongly contrasts with macaque physiology research showing very small changes (or no changes) in firing associated with attentional modulation. Research with the macaque monkey is usually performed by measuring spiking activity from single neurons. The neural basis of the fMRI signal on the other hand is mostly related to post synaptic potentiation (PSP)[citation needed]. This difference therefore does not necessarily indicate a difference between macaque and human physiology.
Other current work on V1 seeks to fully characterize its tuning properties, and to use it as a model area for the canonical cortical circuit.
Lesions to primary visual cortex usually lead to a scotoma, or hole in the visual field. Interestingly, patients with scotomas are often able to make use of visual information presented to their scotomas, despite being unable to consciously perceive it. This phenomenon, called blindsight, is widely studied by scientists interested in the neural correlate of consciousness.
Primary visual cortex (V1)

 Micrograph showing the visual cortex (pink). The pia mater and arachnoid mater including blood vessels is seen at the top of the image. Subcortical white matter (blue) is seen at the bottom of the image. HE-LFB stain.
Micrograph showing the visual cortex (pink). The pia mater and arachnoid mater including blood vessels is seen at the top of the image. Subcortical white matter (blue) is seen at the bottom of the image. HE-LFB stain.The primary visual cortex is the best studied visual area in the brain. In all mammals studied, it is located in the posterior pole of the occipital cortex (the occipital cortex is responsible for processing visual stimuli). It is the simplest, earliest cortical visual area. It is highly specialized for processing information about static and moving objects and is excellent in pattern recognition.
The functionally defined primary visual cortex is approximately equivalent to the anatomically defined striate cortex. The name "striate cortex" is derived from the stria of Gennari, a distinctive stripe visible to the naked eye that represents myelinated axons from the lateral geniculate body terminating in layer 4 of the gray matter.
The primary visual cortex is divided into six functionally distinct layers, labeled 1 through 6. Layer 4, which receives most visual input from the lateral geniculate nucleus (LGN), is further divided into 4 layers, labelled 4A, 4B, 4Cα, and 4Cβ. Sublamina 4Cα receives most magnocellular input from the LGN, while layer 4Cβ receives input from parvocellular pathways.
The average number of neurons in the adult human primary visual cortex, in each hemisphere, has been estimated at around 140 million (Leuba & Kraftsik, Anatomy and Embryology, 1994).
Function
V1 has a very well-defined map of the spatial information in vision. For example, in humans the upper bank of the calcarine sulcus responds strongly to the lower half of visual field (below the center), and the lower bank of the calcarine to the upper half of visual field. Conceptually, this retinotopic mapping is a transformation of the visual image from retina to V1. The correspondence between a given location in V1 and in the subjective visual field is very precise: even the blind spots are mapped into V1. Evolutionarily, this correspondence is very basic and found in most animals that possess a V1. In human and animals with a fovea in the retina, a large portion of V1 is mapped to the small, central portion of visual field, a phenomenon known as cortical magnification. Perhaps for the purpose of accurate spatial encoding, neurons in V1 have the smallest receptive field size of any visual cortex microscopic regions.
The tuning properties of V1 neurons (what the neurons respond to) differ greatly over time. Early in time (40 ms and further) individual V1 neurons have strong tuning to a small set of stimuli. That is, the neuronal responses can discriminate small changes in visual orientations, spatial frequencies and colors. Furthermore, individual V1 neurons in human and animals with binocular vision have ocular dominance, namely tuning to one of the two eyes. In V1, and primary sensory cortex in general, neurons with similar tuning properties tend to cluster together as cortical columns. David Hubel and Torsten Wiesel proposed the classic ice-cube organization model of cortical columns for two tuning properties: ocular dominance and orientation. However, this model cannot accommodate the color, spatial frequency and many other features to which neurons are tuned. The exact organization of all these cortical columns within V1 remains a hot topic of current research.
Current consensus seems to be that early responses of V1 neurons consists of tiled sets of selective spatiotemporal filters. In the spatial domain, the functioning of V1 can be thought of as similar to many spatially local, complex Fourier transforms, or more accurately, Gabor transforms. Theoretically, these filters together can carry out neuronal processing of spatial frequency, orientation, motion, direction, speed (thus temporal frequency), and many other spatiotemporal features. Experiments of neurons substantiate these theories, but also raise new questions.
Later in time (after 100 ms) neurons in V1 are also sensitive to the more global organisation of the scene (Lamme & Roelfsema, 2000). These response properties probably stem from recurrent processing (the influence of higher-tier cortical areas on lower-tier cortical areas) and lateral connections from pyramidal neurons (Hupe et al. 1998). While feedforward connections are mainly driving, feedback connections are mostly modulatory in their effects (Angelucci et al., 2003; Hupe et al., 2001). Evidence shows that feedback originating in higher level areas such as V4, IT or MT, with bigger and more complex receptive fields, can modify and shape V1 responses, accounting for contextual or extra-classical receptive field effects (Guo et al., 2007; Harrison et al., 2007; Huang et al., 2007; Sillito et al., 2006).
The visual information relayed to V1 is not coded in terms of spatial (or optical) imagery, but rather as the local contrast. As an example, for an image comprising half side black and half side white, the divide line between black and white has strongest local contrast and is encoded, while few neurons code the brightness information (black or white per se). As information is further relayed to subsequent visual areas, it is coded as increasingly non-local frequency/phase signals. Importantly, at these early stages of cortical visual processing, spatial location of visual information is well preserved amid the local contrast encoding.
V2
Visual area V2, also called prestriate cortex,[4] is the second major area in the visual cortex, and the first region within the visual association area. It receives strong feedforward connections from V1 (direct and via the pulvinar) and sends strong connections to V3, V4, and V5. It also sends strong feedback connections to V1.
Anatomically, V2 is split into four quadrants, a dorsal and ventral representation in the left and the right hemispheres. Together these four regions provide a complete map of the visual world. Functionally, V2 has many properties in common with V1. Cells are tuned to simple properties such as orientation, spatial frequency, and color. The responses of many V2 neurons are also modulated by more complex properties, such as the orientation of illusory contours and whether the stimulus is part of the figure or the ground (Qiu and von der Heydt, 2005).
Recent research has shown that V2 cells show a small amount of attentional modulation (more than V1, less than V4), are tuned for moderately complex patterns, and may be driven by multiple orientations at different subregions within a single receptive field.
It is argued that the entire ventral visual-to-hippocampal stream is important for visual memory.[5] This theory, unlike the dominant one, predicts that object-recognition memory (ORM) alterations could result from the manipulation in V2, an area that is highly interconnected within the ventral stream of visual cortices. In the monkey brain, this area receives strong feedforward connections from the primary visual cortex (V1) and sends strong projections to other secondary visual cortices (V3, V4, and V5) (,[6][7]). Most of the neurons of this area are tuned to simple visual characteristics such as orientation, spatial frequency, size, color, and shape (,[8][9] and [10]). V2 cells also respond to various complex shape characteristics, such as the orientation of illusory contours ([10]) and whether the stimulus is part of the figure or the ground ([11]). Anatomical studies implicate layer 3 of area V2 in visual-information processing. In contrast to layer 3, layer 6 of the visual cortex is composed of many types of neurons, and their response to visual stimuli is more complex.
In a recent study, the Layer 6 cells of the V2 cortex were found to play a very important role in the storage of Object Recognition Memory as well as the conversion of short-term object memories into long-term memories[12].
Third visual complex, including area V3
The term third visual complex refers to the region of cortex located immediately in front of V2, which includes the region named visual area V3 in humans. The "complex" nomenclature is justified by the fact that some controversy still exists regarding the exact extent of area V3, with some researchers proposing that the cortex located in front of V2 may include two or three functional subdivisions. For example, David Van Essen and others (1986) have proposed the existence of a "dorsal V3" in the upper part of the cerebral hemisphere, which is distinct from the "ventral V3" (or ventral posterior area, VP) located in the lower part of the brain. Dorsal and ventral V3 have distinct connections with other parts of the brain, appear different in sections stained with a variety of methods, and contain neurons that respond to different combinations of visual stimulus (for example, colour-selective neurons are more common in the ventral V3). Additional subdivisions, including V3A and V3B have also been reported in humans. These subdivisions are located near dorsal V3, but do not adjoin V2.
Dorsal V3 is normally considered to be part of the dorsal stream, receiving inputs from V2 and from the primary visual area and projecting to the posterior parietal cortex. It may be anatomically located in Brodmann area 19. Recent work with fMRI has suggested that area V3/V3A may play a role in the processing of global motion [13] Other studies prefer to consider dorsal V3 as part of a larger area, named the dorsomedial area (DM), which contains a representation of the entire visual field. Neurons in area DM respond to coherent motion of large patterns covering extensive portions of the visual field (Lui and collaborators, 2006).
Ventral V3 (VP), has much weaker connections from the primary visual area, and stronger connections with the inferior temporal cortex. While earlier studies proposed that VP only contained a representation of the upper part of the visual field (above the point of fixation), more recent work indicates that this area is more extensive than previously appreciated, and like other visual areas it may contain a complete visual representation. The revised, more extensive VP is referred to as the ventrolateral posterior area (VLP) by Rosa and Tweedale.[14]
V4
Visual area V4 is one of the visual areas in the extrastriate visual cortex. It is located anterior to V2 and posterior to posterior inferotemporal area (PIT). It comprises at least four regions (left and right V4d, left and right V4v), and some groups report that it contains rostral and caudal subdivisions as well. It is unknown what the human homologue of V4 is, and this issue is currently the subject of much scrutiny.[citation needed]
V4 is the third cortical area in the ventral stream, receiving strong feedforward input from V2 and sending strong connections to the PIT. It also receives direct inputs from V1, especially for central space. In addition, it has weaker connections to V5 and dorsal prelunate gyrus (DP).
V4 is the first area in the ventral stream to show strong attentional modulation. Most studies indicate that selective attention can change firing rates in V4 by about 20%. A seminal paper by Moran and Desimone characterizing these effects was the first paper to find attention effects anywhere in the visual cortex [1].[15]
Like V1, V4 is tuned for orientation, spatial frequency, and color. Unlike V1, V4 is tuned for object features of intermediate complexity, like simple geometric shapes, although no one has developed a full parametric description of the tuning space for V4. Visual area V4 is not tuned for complex objects such as faces, as areas in the inferotemporal cortex are.
The firing properties of V4 were first described by Semir Zeki in the late 1970s, who also named the area. Before that, V4 was known by its anatomical description, the prelunate gyrus. Originally, Zeki argued that the purpose of V4 was to process color information. Work in the early 1980s proved that V4 was as directly involved in form recognition as earlier cortical areas. This research supported the Two Streams hypothesis, first presented by Ungerleider and Mishkin in 1982.
Recent work has shown that V4 exhibits long-term plasticity, encodes stimulus salience, is gated by signals coming from the frontal eye fields and shows changes in the spatial profile of its receptive fields with attention.
V5/MT
Visual area V5, also known as visual area MT (middle temporal), is a region of extrastriate visual cortex that is thought to play a major role in the perception of motion, the integration of local motion signals into global percepts and the guidance of some eye movements.[16]
Connections
MT is connected to a wide array of cortical and subcortical brain areas. Its inputs include the visual cortical areas V1, V2, and dorsal V3 (dorsomedial area),[17][18] the koniocellular regions of the LGN,[19] and the inferior pulvinar. The pattern of projections to MT changes somewhat between the representations of the foveal and peripheral visual fields, with the latter receiving inputs from areas located in the midline cortex and retrosplenial region [20]
A standard view is that V1 provides the "most important" input to MT.[16] Nonetheless, several studies have demonstrated that neurons in MT are capable of responding to visual information, often in a direction-selective manner, even after V1 has been destroyed or inactivated.[21] Moreover, research by Semir Zeki and collaborators has suggested that certain types of visual information may reach MT before it even reaches V1.
MT sends its major outputs to areas located in the cortex immediately surrounding it, including areas FST, MST and V4t (middle temporal crescent). Other projections of MT target the eye movement-related areas of the frontal and parietal lobes (frontal eye field and lateral intraparietal area).
Function
The first studies of the electrophysiological properties of neurons in MT showed that a large portion of the cells were tuned to the speed and direction of moving visual stimuli[22][23] These results suggested that MT played a significant role in the processing of visual motion.
Lesion studies have also supported the role of MT in motion perception and eye movements. Neuropsychological studies of a patient who could not see motion, seeing the world in a series of static "frames" instead, suggested that MT in the primate is homologous to V5 in the human.[24][25]
However, since neurons in V1 are also tuned to the direction and speed of motion, these early results left open the question of precisely what MT could do that V1 could not. Much work has been carried out on this region as it appears to integrate local visual motion signals into the global motion of complex objects.[26] For example, lesion to the V5 lead to deficits in perceiving motion and processing of complex stimuli. It contains many neurons selective for the motion of complex visual features (line ends, corners). Microstimulation of a neuron located in the V5 affects the perception of motion. For example, if one finds a neuron with preference for upward motion, and then we use an electrode to stimulate it, the monkey becomes more likely to report 'upward' motion.[27]
There is still much controversy over the exact form of the computations carried out in area MT[28] and some research suggests that feature motion is in fact already available at lower levels of the visual system such as V1.[29][30]
Functional organization
MT was shown to be organized in direction columns.[31] DeAngelis argued that MT neurons were also organized based on their tuning for binocular disparity.[32]
See also
- Brodmann area
- Cortical area
- Cortical blindness
- Feature integration theory
- List of regions in the human brain
- Retinotopy
References
- ^ a b Goodale & Milner; Milner, A.David (1992). "Separate pathways for perception and action.". Trends in Neuroscience 15 (1): 20–25. doi:10.1016/0166-2236(92)90344-8. PMID 1374953.
- ^ Ungerleider and Mishkin (1982). Ingle DJ, Goodale MA and Mansfield RJW. ed. Analysis of Visual Behavior. Boston: MIT Press.
- ^ Franz VH, Scharnowski F, Gegenfurtner (2005). "Illusion effects on grasping are temporally constant not dynamic.". J Exp Psychol Hum Percept Perform. 31 (6): 1359–78. doi:10.1037/0096-1523.31.6.1359. PMID 16366795.
- ^ Gazzaniga, Ivry & Mangun: Cognitive neuroscience, 2002
- ^ T. J. Bussey, L. M. Saksida, Hippocampus 17, 898 (2007).
- ^ I. Stepniewska, J. H. Kaas, J. Comp. Neurol. 371, 129 (1996).
- ^ R. Gattas, A. P. Sousa, M. Mishkin, L. G. Ungerleider, Cereb. Cortex 7, 110 (1997).
- ^ J. Hegde, D. C. Van Essen, J. Neurosci. 20, RC61 (2000).
- ^ J. Hegde, D. C. Van Essen, J. Neurophysiol. 92, 3030 (2004).
- ^ a b A. Anzai, X. Peng, D. C. Van Essen, Nat. Neurosci. 10, 1313 (2007).
- ^ I. Maruko et al., J. Neurophysiol. 100, 2486 (2008).
- ^ http://www.sciencemag.org/cgi/content/full/325/5936/87
- ^ Braddick, OJ, O'Brian, JMD, et al (2001). "Brain areas sensitive to visual motion.". Perception 30 (1): 61–72. doi:10.1068/p3048. PMID 11257978.
- ^ Rosa MG, Tweedale R (2000). "Visual areas in lateral and ventral extrastriate cortices of the marmoset monkey". J Comp Neurol 422 (4): 621–51. doi:10.1002/1096-9861(20000710)422:4<621::AID-CNE10>3.0.CO;2-E. PMID 10861530.
- ^ Moran & Desimone. Selective Attention Gates Visual Processing in the Extrastriate Cortex. Science 229(4715), 1985.
- ^ a b Born R, Bradley D (2005). "Structure and function of visual area MT.". Annu Rev Neurosci 28: 157–89. doi:10.1146/annurev.neuro.26.041002.131052. PMID 16022593.
- ^ Felleman D, Van Essen D (1991). "Distributed hierarchical processing in the primate cerebral cortex.". Cereb Cortex 1 (1): 1–47. doi:10.1093/cercor/1.1.1-a. PMID 1822724.
- ^ Ungerleider L, Desimone R (1986). "Cortical connections of visual area MT in the macaque.". J Comp Neurol 248 (2): 190–222. doi:10.1002/cne.902480204. PMID 3722458.
- ^ Sincich L, Park K, Wohlgemuth M, Horton J (2004). "Bypassing V1: a direct geniculate input to area MT.". Nat Neurosci 7 (10): 1123–8. doi:10.1038/nn1318. PMID 15378066.
- ^ Palmer SM, Rosa MG (2006). "A distinct anatomical network of cortical areas for analysis of motion in far peripheral vision.". Eur J Neurosci 24 (8): 2389–405. doi:10.1111/j.1460-9568.2006.05113.x. PMID 17042793.
- ^ Rodman HR, Gross CG, Albright TD (1989). "Afferent basis of visual response properties in area MT of the macaque. I. Effects of striate cortex removal". J Neurosci 9 (6): 2033–50. PMID 2723765.
- ^ Dubner R, Zeki S (1971). "Response properties and receptive fields of cells in an anatomically defined region of the superior temporal sulcus in the monkey.". Brain Res 35 (2): 528–32. doi:10.1016/0006-8993(71)90494-X. PMID 5002708..
- ^ Maunsell J, Van Essen D (1983). "Functional properties of neurons in middle temporal visual area of the macaque monkey. I. Selectivity for stimulus direction, speed, and orientation.". J Neurophysiol 49 (5): 1127–47. PMID 6864242.
- ^ Hess, Baker, Zihl (1989). "The" motion-blind" patient: low-level spatial and temporal filters". Journal of Neuroscience 9 (5): 1628–1640. PMID 2723744.
- ^ Baker, Hess, Zihl (1991). "Residual motion perception in a" motion-blind" patient, assessed with limited-lifetime random dot stimuli". Journal of Neuroscience 11 (2): 454–461. PMID 1992012.
- ^ Movshon, J.A., Adelson, E.H., Gizzi, M.S., & Newsome, W.T. (1985). The analysis of moving visual patterns. In: C. Chagas, R. Gattass, & C. Gross (Eds.), Pattern recognition mechanisms (pp. 117-151), Rome: Vatican Press.
- ^ Britten & Van Wezel 1998
- ^ Wilson, H.R., Ferrera, V.P., & Yo, C. (1992). A psychophysically motivated model for two-dimensional motion perception. Vis Neurosci, 9 (1), 79-97.
- ^ Tinsley, C.J., Webb, B.S., Barraclough, N.E., Vincent, C.J., Parker, A., & Derrington, A.M. (2003). The nature of V1 neural responses to 2D moving patterns depends on receptive-field structure in the marmoset monkey. J Neurophysiol, 90 (2), 930-937.
- ^ Pack & Born, 2003
- ^ Albright T (1984). "Direction and orientation selectivity of neurons in visual area MT of the macaque.". J Neurophysiol 52 (6): 1106–30. PMID 6520628.
- ^ DeAngelis G, Newsome W (1999). "Organization of disparity-selective neurons in macaque area MT.". J Neurosci 19 (4): 1398–415. PMID 9952417.
External links
- The Primary Visual Cortex by Matthew Schmolesky at University of Utah
- Architecture of the Visual Cortex, by David Hubel at Harvard University
- NeuroNames ancil-415 - striate area 17
- NeuroNames ancil-699 - Brodmann area 17 in guenon
- BrainMaps at UCDavis visual%20cortex
- Simulator for computational modeling of visual cortex maps at topographica.org
Sensory system: Visual system and eye movement pathways Visual perception 1° (Bipolar cell of Retina) → 2° (Ganglionic cell) → 3° (Optic nerve → Optic chiasm → Optic tract → LGN of Thalamus) → 4° (Optic radiation → Cuneus and Lingual gyrus of Visual cortex → Blobs → Globs)Muscles of orbit TrackingHorizontal gazeVertical gazePupillary reflex Pupillary dilation1° (Posterior hypothalamus → Ciliospinal center) → 2° (Superior cervical ganglion) → 3° (Sympathetic root of ciliary ganglion → Nasociliary nerve → Long ciliary nerves → Iris dilator muscle)1° (Retina → Optic nerve → Optic chiasm → Optic tract → Visual cortex → Brodmann area 19 → Pretectal area) → 2° (Edinger-Westphal nucleus) → 3° (Short ciliary nerves → Ciliary ganglion → Ciliary muscle)Circadian rhythm M: EYE
anat(g/a/p)/phys/devp/prot
noco/cong/tumr, epon
proc, drug(S1A/1E/1F/1L)


Wikimedia Foundation. 2010.