- Suprachiasmatic nucleus
Infobox Brain
Name = PAGENAME
Latin = nucleus suprachiasmaticus
GraySubject =
GrayPage =

Caption = Suprachiasmatic nucleus is 'SC', at center left, in blue. OC, in black, isoptic chiasm .
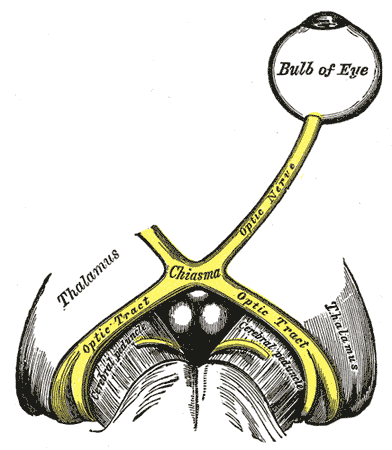
Caption2 = The left optic nerve and the optic tracts. (Suprachiasmatic nucleus not labeled, but diagram illustrates region.)
IsPartOf =
Components =
Artery =
Vein =
Acronym =
BrainInfoType = hier
BrainInfoNumber = 367
MeshName = Suprachiasmatic+Nucleus
MeshNumber = A08.186.211.730.385.357.342.625
DorlandsPre = n_11z
DorlandsSuf = 12583563
The suprachiasmatic nucleus (SCN) is a bilateral region of the brain, located in thehypothalamus , that is responsible for controlling endogenouscircadian rhythms . The neuronal and hormonal activities it generates regulate many different body functions over a 24-hour period.The SCN contains several cell types and several different
peptide s (includingvasopressin andvasoactive intestinal peptide ) andneurotransmitter s, and interacts with many other regions of the brain.Location
The SCN is situated in the anterior
hypothalamus immediately dorsal and superior (hence supra) to theoptic chiasm (CHO) bilateral to (on either side of) thethird ventricle .Circadian effects
The SCN receives inputs from specialized photoreceptive retinal
ganglion cells , via theretinohypothalamic tract .Destruction of the SCN leads to a complete loss of circadian rhythm. Rats with damage to the SCN have no
circadian rhythm s, i.e., they sleep the same total amount, but polyphasically for random lengths at a time.The SCN also controls 'slave oscillators' in the peripheral tissues, which exhibit their own ~24 hour rhythms, but are crucially synchronized by the SCN. [cite journal | last = Bernard | first = Samuel | coauthors = Didier Gonze, Branka Čajavec, Hanspeter Herzel, Achim Kramer | date = 2007-04-13 | year = | month = | title = Synchronization-Induced Rhythmicity of Circadian Oscillators in the Suprachiasmatic Nucleus | journal = PLoS Computational Biology | volume = 3 | issue = 4 | pages =
publisher = | location = | issn = | doi = | bibcode = | oclc = | id = | url = http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pcbi.0030068 | format = | accessdate = 2008-04-02 | quote = ]The importance of entraining our bodies to an exogenous cue, such as daylight, is reflected by several
circadian rhythm sleep disorders , where this process does not function normally.Neurons in the "ventrolateral SCN" (vlSCN) have the ability for light-induced gene expression. If light is turned on at night, the vlSCN relays this information throughout the SCN, in a process called "entrainment".
Neurons in the "dorsomedial SCN" (dmSCN) are believed to make an endogenous 24-hour rhythm that can persist under constant darkness (in humans averaging about 24h 11min).
Melanopsin -containingganglion cell s in theretina have a direct connection to the SCN via the retinohypothalamic tract.The SCN sends information to other hypothalamic nuclei and the
pineal gland to modulate body temperature and production of hormones such ascortisol andmelatonin .Other signals from the retina
The SCN is one of four nuclei that receive nerve signals directly from the retina.
The other three are the
lateral geniculate nucleus (LGN), thesuperior colliculus , and thepretectum :* The "LGN" passes information about color, contrast, shape, and movement on to the
visual cortex and itself signals to the SCN.* The "superior colliculus" controls the movement and orientation of the eyeball.
* The "pretectum" controls the size of the
pupil .Gene expression
The circadian rhythm in the SCN is generated by a
gene expression cycle in individual SCN neurons. This cycle has been well conserved through evolution, and is essentially similar in cells from many widely different organisms that show circadian rhythms.Fruitfly
For example, in the fruitfly "
Drosophila ", the cellular circadian rhythm in neurons is controlled by two interlocked feedback loops.
* In the first loop, the bHLH transcription factors "clock" ("clk") and "cycle" ("cyc") drive the transcription of their own repressors "period" ("per") and "timeless" ("tim"). PER and TIM proteins then accumulate in the cytoplasm, translocate into the nucleus at night, and turn off their own transcription, thereby setting up a 24 hour oscillation of transcription and translation.
* In the second loop, the transcription factors "vrille" ("vri") and "Pdp1" are initiated by CLK/CYC. PDP1 acts positively on Clk transcription and negatively on VRI.These genes encode various
transcription factors that trigger expression of other proteins. The products of "clock" and "cycle", called CLK and CYC, belong to the PAS-containing subfamily of thebasic-helix-loop-helix (bHLH) family oftranscription factors , and form aheterodimer . This heterodimer (CLK-CYC) initiates the transcription of "per" and "tim", whose protein products dimerize and then inhibit their own expression by disrupting CLK-CYC-mediated transcription. This negative feedback mechanism gives a 24-hour rhythm in the expression of theclock genes . Many genes are suspected to be linked to circadian control by "E-box elements" in their promoters, as CLK-CYC and its homologs bind to these elements.The 24-hr rhythm could be reset by light via the protein CRYPTOCHROME (CRY), which is involved in the circadian photoreception in "Drosophila." CRY associates with TIM in a light-dependent manner that leads to the destruction of TIM. Without the presence of TIM for stabilization, PER is eventually destroyed during the day. As a result, the repression of CLK-CYC is reduced and the whole cycle reinitiates again.
Mammals
In mammals, circadian clock genes behave in a similar manner.
CLOCK (circadian locomotor output cycles kaput) was first cloned in mouse andBMAL1 (brain and muscle aryl hydrocarbon receptor nuclear translocator (ARNT)-like 1) is the primary homolog of "Drosophila" CYC.Three homologs of PER (
PER1 ,PER2 , andPER3 ) and two CRY homologs (CRY1 andCRY2 ) have been identified.TIM has been identified in mammals, however, its function is still not determined.
Recent research suggests that, outside the SCN clock, genes may have other important roles as well, including their influence on the effects of drugs of abuse such as
cocaine . [cite journal |author=Yuferov V, Butelman E, Kreek M |title=Biological clock: biological clocks may modulate drug addiction |journal=Eur J Hum Genet |volume=13 |issue=10 |pages=1101–3 |year=2005 |pmid=16094306 |doi=10.1038/sj.ejhg.5201483] [cite journal |author=Manev H, Uz T |title=Clock genes as a link between addiction and obesity |journal=Eur J Hum Genet |volume=14 |issue=1 |pages=5 |year=2006 |pmid=16288309 |doi=10.1038/sj.ejhg.5201524]Electrophysiology
Neurons in the SCN fire
action potential s in a 24-hour rhythm. At mid-day, the firing rate reaches a maximum, and, during the night, it falls again. How the gene expression cycle (so-called the core clock) connects to the neural firing remains unknown.Many SCN neurons are sensitive to light stimulation via the retina, and sustainedly firing action potentials during a light pulse (~30 seconds) in rodents. The photic response is likely linked to effects of light on circadian rhythms. In addition, focal application of melatonin can decrease firing activity of these neurons, suggesting that melatonin receptors present in the SCN mediate phase-shifting effects through the SCN.
Calcium dynamics
Two contradictory reports exist about circadian variation of the cell calcium concentration. However, both reports agree that the resting calcium level is slightly higher during the day than at night.
exuality and the SCN
A member of the American Psychiatric Association, Professor D.F.Swaab carried out research into this particular hypothalamic component whilst trying to find evidence of an organic basis for homosexuality in humans. He found the suprachiasmatic nucleus to be nearly twice the size in male homosexual subjects as in their heterosexual counterparts. [cite journal |author=Swaab DF, Hofman MA |title=An enlarged suprachiasmatic nucleus in homosexual men |journal=Brain Res. |volume=537 |issue=1-2 |pages=141–8 |year=1990 |pmid=2085769 |doi=10.1016/0006-8993(90)90350-K] This research was further confirmed by Laura S. Allen, who found the midsagittal plane of the anterior commissure of the hypothalamus to be one third larger in male homosexual subjects than in male heterosexuals. [cite journal |author=Allen LS, Gorski RA |title=Sexual orientation and the size of the anterior commissure in the human brain |journal=Proc. Natl. Acad. Sci. U.S.A. |volume=89 |issue=15 |pages=7199–202 |year=1992 |pmid=1496013 |doi=10.1073/pnas.89.15.7199] As suggested by the author of the article: "This anatomical difference, which correlates with gender and sexual orientation, may, in part, underlie differences in cognitive function and cerebral lateralization among homosexual men, heterosexual men, and heterosexual women". [cite journal |author=Allen LS, Gorski RA |title=Sexual orientation and the size of the anterior commissure in the human brain |journal=Proc. Natl. Acad. Sci. U.S.A. |volume=89 |issue=15 |pages=7199–202 |year=1992 |pmid=1496013 |doi=10.1073/pnas.89.15.7199] However, as it is put by Jeffrey Satinover, M.D “The specifics of these findings are not as important as realizing that unless group differences are dramatic, individual studies of such differences mean almost nothing. It would take hundreds, perhaps thousands of such studies before meaningful trend emerges”. It is also true that the brain may reconfigure itself in response to certain experiences. Research by G. Gabbard published in the American Journal of Psychiatry 149, no. 8 points that the establishment of new connections between cells, dramatically increase or decreases in the thickness of connections between cells in response to training, trauma and especially traumatic experience altering brain and body of monkeys in measurable ways.
ee also
*
Chronobiology
*Photosensitive ganglion cell
*Sense of time References
External links
* [http://thebrain.mcgill.ca/flash/a/a_02/a_02_cr/a_02_cr_vis/a_02_cr_vis.html Diagram at thebrain.mcgill.ca]
Wikimedia Foundation. 2010.