- Cryptochrome
-
Cryptochrome 1 (photolyase-like) 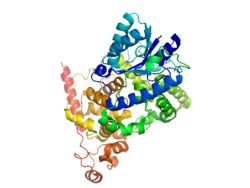
Crystal structure of the PHR domain of cryptochrome 1 from Arabidopsis thaliana.[1]Identifiers Symbols CRY1; PHLL1 External IDs OMIM: 601933 MGI: 1270841 HomoloGene: 7042 GeneCards: CRY1 Gene Gene Ontology Molecular function • nucleotide binding
• DNA binding
• DNA photolyase activity
• receptor activity
• G-protein coupled photoreceptor activity
• lyase activityCellular component • nucleus
• mitochondrionBiological process • DNA repair
• transcription
• regulation of transcription, DNA-dependent
• visual perception
• circadian rhythm
• protein-chromophore linkage
• response to stimulusSources: Amigo / QuickGO RNA expression pattern 
More reference expression data Orthologs Species Human Mouse Entrez 1407 12952 Ensembl ENSG00000008405 ENSMUSG00000020038 UniProt Q16526 P97784 RefSeq (mRNA) NM_004075 NM_007771 RefSeq (protein) NP_004066 NP_031797 Location (UCSC) Chr 12:
105.91 – 106.01 MbChr 10:
84.56 – 84.61 MbPubMed search [1] [2] Cryptochrome 2 (photolyase-like) Identifiers Symbols CRY2; FLJ10332; HCRY2; KIAA0658; PHLL2 External IDs OMIM: 603732 MGI: 1270859 HomoloGene: 56466 GeneCards: CRY2 Gene Gene Ontology Molecular function • nucleotide binding
• DNA photolyase activity
• protein binding
• photoreceptor activityCellular component • nucleus
• cytoplasmBiological process • DNA repair
• regulation of transcription, DNA-dependent
• protein import into nucleus
• circadian rhythm
• protein-chromophore linkage
• response to stimulusSources: Amigo / QuickGO RNA expression pattern 
More reference expression data Orthologs Species Human Mouse Entrez 1408 12953 Ensembl ENSG00000121671 ENSMUSG00000068742 UniProt Q49AN0 Q99JJ1 RefSeq (mRNA) NM_001127457.1 XM_994658 RefSeq (protein) NP_001120929.1 XP_999752 Location (UCSC) Chr 11:
45.87 – 45.9 MbChr 2:
92.24 – 92.27 MbPubMed search [3] [4] Cryptochromes (from the Greek κρυπτό χρώμα, hidden colour) are a class of blue light-sensitive flavoproteins found in plants and animals. Cryptochromes are involved in the circadian rhythms of plants and animals, and in the sensing of magnetic fields in a number of species. The name Cryptochrome was proposed as a pun combining the cryptic nature of the photoreceptor, and the cryptogamic organisms on which many blue light studies were carried out. [2]
The two genes Cry1 and Cry2 code for the two cryptochrome proteins CRY1 and CRY2.[3] In insects and plants, CRY1 regulates the circadian clock in a light-dependent fashion, whereas in mammals, CRY1 and CRY2 act as light-independent inhibitors of CLOCK-BMAL1 components of the circadian clock.[4] In plants, blue light photoreception can be used to cue developmental signals.[1]
Contents
Discovery
Although Charles Darwin first documented plant responses to blue light in the 1800s, it was not until the 1980s that research began to identify the pigment responsible.[5] In 1980, researchers discovered that the HY4 gene of the plant Arabidopsis thaliana was necessary for the plant's blue light sensitivity, and when the gene was sequenced in 1993, it showed high sequence homology with photolyase, a DNA repair protein activated by blue light.[6] By 1995, it became clear that the products of the HY4 gene and its two human homologs did not exhibit photolyase activity and were instead a new class of blue light photoreceptor hypothesized to be circadian photopigments.[7] In 1996 and 1998, Cry homologs were identified in Drosophila and mice, respectively.[8][9]
Evolutionary history and structure
Cryptochromes (CRY1,CRY2) are evolutionarily old and highly conserved proteins that belong to the flavoproteins superfamily that exists in all kingdoms of life.[1] All members of this superfamily have the characteristics of an N-terminal photolyase homology (PHR) domain. The PHR domain can bind to the flavin adenine dinucleotide (FAD) cofactor and a light-harvesting chromophore.[1] Cryptochromes are derived from and closely related to photolyases, which are bacterial enzymes that are activated by light and involved in the repair of UV-induced DNA damage. In eukaryotes, cryptochromes no longer retain this original enzymatic activity.[10]
The structure of cryptochrome involves a fold very similar to that of photolyase, with a single molecule of FAD noncovalently bound to the protein.[1] These proteins have variable lengths and surfaces on the C-terminal end, due to the changes in genome and appearance that result from the lack of DNA repair enzymes. The Ramachandran plot[11] shows that the secondary structure of the CRY1 protein is primarily a right-handed alpha helix with little to no steric overlap.[12] The structure of CRY1 is almost entirely made up of alpha helices, with several loops and few beta sheets. The molecule is arranged as an orthogonal bundle.[1]
Function
Phototropism
In plants, cryptochromes mediate phototropism, or directional growth towards a light source, in response to blue light. This response is now known to have its own set of photoreceptors, the phototropins. Unlike phytochromes and phototropins, cryptochromes are not kinases. Their flavin chromophore is reduced by light and transported into the cell nucleus, where it affects the turgor pressure and causes subsequent stem elongation. Specifically, Cry2 is responsible for blue-light mediated cotyledon and leaf expansion. Cry2 overexpression in transgenic plants increases blue light-stimulated cotyledon expansion, which results in many broad leaves and no flowers, rather than a few primary leaves with a flower.[13] A double loss-of-function mutation in Arabidopsis thaliana Early Flowering 3 (elf3) and Cry2 genes delays flowering under continuous light was shown to accelerates it during long and short days, which suggests that Arabidopsis CRY2 may play a role in accelerating flowering time during continuous light.[14]
Light capture
Despite much research on the topic, cryptochrome photoreception and phototransduction in Drosophila and Arabidopsis thaliana is still poorly understood. Cryptochromes are known to possess two chromophores: pterin (in the form of 5,10-methenyltetrahydrofolic acid (MHF)) and flavin (in the form of FAD).[15] Both may absorb a photon, and in Arabidopsis, pterin appears to absorb at a wavelength of 380 nm and flavin at 450 nm. Past studies have supported a model by which energy captured by pterin is transferred to flavin.[16] Under this model of phototransduction, FAD would then be reduced to FADH, which probably mediates the phosphorylation of a certain domain in cryptochrome. This could then trigger a signal transduction chain, possibly affecting gene regulation in the cell nucleus.
Recent research has indicated that a different mechanism may function in Drosophila. The true ground state of the flavin cofactor in Drosophila CRY is still debated, with some models indicating the FAD is in an oxidized form [17], while others support a model in which the flavin cofactor exists in anion radical form, FAD•-. Recently, researchers have observed that oxidized FAD is readily reduced to FAD•- by light. Furthermore, mutations that blocked photoreduction had no effect on light-induced degradation of CRY, while mutations that altered the stability of FAD•- destroyed CRY photoreceptor function.[18][19]These observations provide support for a ground state of FAD•-. Researchers have also recently proposed a model in which FAD•- is excited to its doublet or quartet state by absorption of a photon, which then leads to a conformational change in the CRY protein.[20]
Circadian rhythm
Studies in animals and plants suggest that cryptochromes play a pivotal role in the generation and maintenance of circadian rhythms.[21] In Drosophila, cryptochrome (dCRY) acts as a blue-light photoreceptor that directly modulates light input into the circadian clock[22], while in mammals, cryptochromes (CRY1 and CRY2) act as transcription repressors within the circadian clockwork.[23] Some insects, including the monarch butterfly, have both a mammal-like and a Drosophila-like version of cryptochrome, providing evidence for an ancestral clock mechanism involving both light sensing and transcriptional repression roles for cryptochrome.[24][25]
Cry mutants have altered circadian rhythms, showing that Cry affects the circadian pacemaker. Drosophila with mutated Cry exhibit little to no mRNA cycling.[26] A point mutation in cryb, which is required for flavin association in CRY protein, results in no PER or TIM protein cycling in either DD or LD.[27] In addition, mice lacking Cry1 or Cry2 genes exhibit differentially altered free running periods, but are still capable of photoentrainment. However, mice that lack both Cry1 and Cry2 are arrhythmic in both LD and DD and always have high Per1 mRNA levels. These results suggest that cryptochromes play a photoreceptive role, as well as acting as negative regulators of Per gene expression in mice.[28]
In Drosophila
In Drosophila, cryptochrome functions as a blue light photoreceptor. Exposure to blue light induces a conformation similar to that of the always active CRY mutant with a C-terminal deletion (CRYΔ).[20] The half-life of this conformation is 15 minutes in the dark and facilitates the binding of CRY to other clock gene products, PER and TIM, in a light-dependent manner.[20][22][4][29] Once bound by dCRY, dTIM is committed to degradation by the ubiquitin-proteasome system.[29][20]
Although light pulses do not entrain, full photoperiod LD cycles can still drive cycling in the ventral-lateral neurons in the Drosophila brain. These data along with other results suggest that CRY is the cell-autonomous photoreceptor for body clocks in Drosophila and may play a role in nonparametric entrainment (entrainment by short discrete light pulses). However, the lateral neurons receive light information through both the blue light CRY pathway and the rhodopsin pathway. Therefore, CRY is involved in light perception and is an input to the circadian clock, but is not required for the operation of the clock because the rhodopsin pathway is sufficient in the absence of the CRY pathway.[30] Recently, it has also been shown that there is a CRY-mediated light response that is independent of the classical circadian CRY-TIM interaction. This mechanism is believed to require a flavin redox-based mechanism that is dependent on potassium channel conductance. This CRY mediated light response has been shown to increase action potential firing within seconds of a light response in opsin-knockout Drosophila.[31]
Cryptochrome, like many genes involved in circadian rhythm, shows circadian cycling in mRNA and protein levels. In Drosophila, Cry mRNA concentrations cycle under a light-dark cycle (LD), with high levels in light and low levels in the dark.[26] This cycling persists in constant darkness (DD), but with decreased amplitude.[26] The transcription of the Cry gene also cycles with a similar trend.[26] CRY protein levels, however, cycle in a different manner than Cry transcription and mRNA levels. In LD, CRY protein has low levels in light and high levels in dark, and in DD, CRY levels increase continuously throughout the subjective day and night.[26] Thus, CRY expression is regulated by the clock at the transcriptional level and by light at the translational and posttranslational level.[26]
Overexpression of Cry also affects circadian light responses. In Drosophila, Cry overexpression increases flies’ sensitivity to low intensity light.[26] This light regulation of CRY protein levels suggests that CRY has a circadian role upstream of other clock genes and components.[26]
In mammals
Cryptochrome is one of the four groups of mammalian clock genes/proteins that generate a transcription-translation negative-feedback loop (TTFL), along with Period (PER), CLOCK, and BMAL1.[32] In this loop, CLOCK and BMAL1 proteins are transcriptional activators, which together bind to the promoters of the Cry and Per genes and activate their transcription.[32] The CRY and PER proteins then bind to each other, enter the nucleus, and inhibit CLOCK-BMAL1 activated transcription.[32]
In mice, Cry1 expression displays circadian rhythms in the suprachiasmatic nucleus, a brain region involved in the generation of circadian rhythms, with mRNA levels peaking during the light phase and reaching a minimum in the dark.[33]These daily oscillations in expression are maintained in constant darkness. [33]
While CRY has been well established as a TIM homolog in mammals, the role of CRY as a photoreceptor in mammals has been controversial. Early papers indicated that CRY has both light-independent and dependent functions. A study in 2000 indicated that mice without rhodopsin but with cryptochrome still respond to light; however, in mice without either rhodopsin or cryptochrome, c-Fos transcription, a mediator of light sensitivity, significantly drops.[34] In recent years, data have supported melanopsin as the main circadian photoreceptor, particularly melanopsin cells which mediate entrainment and communication between the eye and the suprachiasmatic nucleus (SCN).[35] One of the main difficulties in confirming or denying CRY as a mammalian photoreceptor is that when the gene is knocked out the animal goes arrhythmic, so it is hard to measure its capacity as purely a photoreceptor. However, some recent studies indicate that human CRY may mediate light response in peripheral tissues.[36]
Normal mammalian circadian rhythm relies critically on delayed expression of Cry1 following activation of the Cry1 promoter. Whereas rhythms in Per2 promoter activation and Per2 mRNA levels have almost the same phase, Cry1 mRNA production is delayed by approximately four hours relative to Cry1 promoter activation.[37] This delay is independent of CRY1 or CRY2 levels and is mediated by a combination of E/E’-box and D-box elements in the promoter and RevErbA/ROR binding elements (RREs) in the gene’s first intron.[38] Transfection of arrhythmic Cry1-/- Cry2-/- double-knockout cells with only the Cry1 promoter (causing constitutive Cry1 expression) is not sufficient to rescue rhythmicity. Transfection of these cells with both the promoter and the first intron is required for restoration of circadian rhythms in these cells.[38]
Magnetoception
Cryptochromes in the photoreceptor neurons of birds' eyes are involved in magnetic orientation during migration.[39] Cryptochromes are also essential for the light-dependent ability of Drosophila to sense magnetic fields.[40] Furthermore, magnetic fields affect cryptochromes in Arabidopsis thaliana: growth behavior is affected by magnetic fields in the presence of blue ( but not red) light.[41]
According to one model,[42] cryptochrome forms a pair of two radicals with correlated spins when exposed to blue light. The occurrence of such light-generated radical pairs and the correlation of the radical pair state have been confirmed recently in a cryptochrome of Xenopus laevis.[43] However, recent evidence from Arabidopsis thaliana cryptochrome also suggests that radical pairs can be generated by the light-independent dark reoxidation of Flavin protein by molecular oxygen through the formation of a spin-correlated FADH-superoxide radical pairs.[44] Magnetoception is hypothesized to function through the surrounding magnetic fields effect on the correlation (parallel or anti-parallel) of these radicals, which affects the duration that cryptochrome remains activated. Activation of cryptochrome may affect the light-sensitivity of retinal neurons, with the overall result that the animal can "see" the magnetic field.[45]
References
- ^ a b c d e f PDB 1u3c; Brautigam CA, Smith BS, Ma Z, Palnitkar M, Tomchick DR, Machius M, Deisenhofer J (August 2004). "Structure of the photolyase-like domain of cryptochrome 1 from Arabidopsis thaliana". Proc. Natl. Acad. Sci. U.S.A. 101 (33): 12142–7. doi:10.1073/pnas.0404851101. PMC 514401. PMID 15299148. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=514401.
- ^ Gressel J (1979). "BLUE LIGHT PHOTORECEPTION". Photochemistry and Photobiology 30 (3): 749–54. doi:10.1111/j.1751-1097.1979.tb07209.x.
- ^ van der Spek PJ, Kobayashi K, Bootsma D, Takao M, Eker AP, Yasui A (October 1996). "Cloning, tissue expression, and mapping of a human photolyase homolog with similarity to plant blue-light receptors". Genomics 37 (2): 177–82. doi:10.1006/geno.1996.0539. PMID 8921389.
- ^ a b Griffin EA, Staknis D, Weitz CJ (October 1999). "Light-independent role of CRY1 and CRY2 in the mammalian circadian clock". Science 286 (5440): 768–71. doi:10.1126/science.286.5440.768. PMID 10531061.
- ^ Darwin, Charles (1881). The Power of Movement in Plants. New York: D. Appleton and Company.
- ^ Ahmad M, Cashmore AR (November 1993). "HY4 gene of A. thaliana encodes a protein with characteristics of a blue-light photoreceptor". Nature 366 (6451): 162–6. Bibcode 1993Natur.366..162A. doi:10.1038/366162a0. PMID 8232555.
- ^ Thompson CL, Sancar A (2004). "Cryptochrome: Discovery of a Circadian Photopigment". In Lenci F, Horspool WM. CRC handbook of organic photochemistry and photobiology. Boca Raton: CRC Press. pp. 1381–89. ISBN 0-8493-1348-1.
- ^ Todo T, Ryo H, Yamamoto K, Toh H, Inui T, Ayaki H, Nomura T, Ikenaga M (April 1996). "Similarity among the Drosophila (6-4) photolyase, a human photolyase homolog, and the DNA photolyase-blue-light photoreceptor family". Science 272 (5258): 109–12. Bibcode 1996Sci...272..109T. doi:10.1126/science.272.5258.109. PMID 8600518.
- ^ Kobayashi K, Kanno S, Smit B, van der Horst GT, Takao M, Yasui A (November 1998). "Characterization of photolyase/blue-light receptor homologs in mouse and human cells". Nucleic Acids Res. 26 (22): 5086–92. doi:10.1093/nar/26.22.5086. PMC 147960. PMID 9801304. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=147960.
- ^ Weber S (February 2005). "Light-driven enzymatic catalysis of DNA repair: a review of recent biophysical studies on photolyase". Biochim. Biophys. Acta 1707 (1): 1–23. doi:10.1016/j.bbabio.2004.02.010. PMID 15721603.
- ^ "MolProbity Ramachandran analysis,1U3C, model 1" (PDF). www.rcsb.org. http://www.rcsb.org/pdb/images/1U3C_ram_m_500.pdf.
- ^ Nelson DR, Lehninger AL, Cox M (2005). Lehninger Principles of Biochemistry. New York: W.H. Freeman. ISBN 0-7167-4339-6.
- ^ Hsu DS, Zhao X, Zhao S, Kazantsev A, Wang RP, Todo T, Wei YF, Sancar A (Dec 1996). "Putative human blue-light photoreceptors hCRY1 and hCRY2 are flavoproteins". Biochemistry 35 (44): 13871–7. doi:10.1021/bi962209o. PMID 8909283.
- ^ Nefissi R, Natsui Y, Miyata K, Oda A, Hase Y, Nakagawa M, Ghorbel A, Mizoguchi T (February 2011). "Double loss-of-function mutation in EARLY FLOWERING 3 and CRYPTOCHROME 2 genes delays flowering under continuous light but accelerates it under long days and short days: an important role for Arabidopsis CRY2 to accelerate flowering time in continuous light". J. Exp. Bot. 62 (8): 2731–2744. doi:10.1093/jxb/erq450. PMID 21296763.
- ^ Song SH, Dick B, Penzkofer A, Pokorny R, Batschauer A, Essen LO (October 2006). "Absorption and fluorescence spectroscopic characterization of cryptochrome 3 from Arabidopsis thaliana". J. Photochem. Photobiol. B, Biol. 85 (1): 1–16. doi:10.1016/j.jphotobiol.2006.03.007. PMID 16725342.
- ^ Hoang N, Bouly JP, Ahmad M (January 2008). "Evidence of a light-sensing role for folate in Arabidopsis cryptochrome blue-light receptors". Mol Plant 1 (1): 68–74. doi:10.1093/mp/ssm008. PMID 20031915.
- ^ Berndt A, Kottke T, Breitkreuz H et al. (April 2007). "A novel photoreaction mechanism for the circadian blue light photoreceptor Drosophila cryptochrome". J. Biol. Chem. 282 (17): 13011–21. doi:10.1074/jbc.M608872200. PMID 17298948.
- ^ Song SH, Oztürk N, Denaro TR, Arat NO, Kao YT, Zhu H, Zhong D, Reppert SM, Sancar A (June 2007). "Formation and function of flavin anion radical in cryptochrome 1 blue-light photoreceptor of monarch butterfly". J. Biol. Chem. 282 (24): 17608–12. doi:10.1074/jbc.M702874200. PMID 17459876.
- ^ Oztürk N, Song SH, Selby CP, Sancar A (February 2008). "Animal type 1 cryptochromes. Analysis of the redox state of the flavin cofactor by site-directed mutagenesis". J. Biol. Chem. 283 (6): 3256–63. doi:10.1074/jbc.M708612200. PMID 18056988.
- ^ a b c d Ozturk N, Selby CP, Annayev Y, Zhong D, Sancar A (January 2011). "Reaction mechanism of Drosophila cryptochrome". Proc. Natl. Acad. Sci. U.S.A. 108 (2): 516–21. Bibcode 2011PNAS..108..516O. doi:10.1073/pnas.1017093108. PMC 3021015. PMID 21187431. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3021015.
- ^ Klarsfeld A, Malpel S, Michard-Vanhée C, Picot M, Chélot E, Rouyer F (February 2004). "Novel features of cryptochrome-mediated photoreception in the brain circadian clock of Drosophila". J. Neurosci. 24 (6): 1468–77. doi:10.1523/JNEUROSCI.3661-03.2004. PMID 14960620.
- ^ a b Emery P, Stanewsky R, Helfrich-Förster C, Emery-Le M, Hall JC, Rosbash M (May 2000). "Drosophila CRY is a deep brain circadian photoreceptor". Neuron 26 (2): 493–504. doi:10.1016/S0896-6273(00)81181-2. PMID 10839367.
- ^ Reppert SM, Weaver DR (August 2002). "Coordination of circadian timing in mammals". Nature 418 (6901): 935–41. doi:10.1038/nature00965. PMID 12198538.
- ^ Zhu H, Yuan Q, Briscoe AD, Froy O, Casselman A, Reppert SM (December 2005). "The two CRYs of the butterfly". Curr. Biol. 15 (23): R953–54. doi:10.1016/j.cub.2005.11.030. PMID 16332522.
- ^ Zhu H, Sauman I, Yuan Q, Casselman A, Emery-Le M, Emery P, Reppert SM (January 2008). "Cryptochromes define a novel circadian clock mechanism in monarch butterflies that may underlie sun compass navigation". PLoS Biol. 6 (1): e4. doi:10.1371/journal.pbio.0060004. PMC 2174970. PMID 18184036. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2174970.
- ^ a b c d e f g h Emery P, So WV, Kaneko M, Hall JC, Rosbash M (November 1998). "CRY, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity". Cell 95 (5): 669–79. doi:10.1016/S0092-8674(00)81637-2. PMID 9845369.
- ^ Stanewsky R, Kaneko M, Emery P, Beretta B, Wager-Smith K, Kay SA, Rosbash M, Hall JC (November 1998). "The cryb mutation identifies cryptochrome as a circadian photoreceptor in Drosophila". Cell 95 (5): 681–92. doi:10.1016/S0092-8674(00)81638-4. PMID 9845370.
- ^ Vitaterna MH, Selby CP, Todo T, Niwa H, Thompson C, Fruechte EM, Hitomi K, Thresher RJ, Ishikawa T, Miyazaki J, Takahashi JS, Sancar A (October 1999). "Differential regulation of mammalian period genes and circadian rhythmicity by cryptochromes 1 and 2". Proc. Natl. Acad. Sci. U.S.A. 96 (21): 12114–19. Bibcode 1999PNAS...9612114V. doi:10.1073/pnas.96.21.12114. PMC 18421. PMID 10518585. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=18421.
- ^ a b Busza A, Emery-Le M, Rosbash M, Emery P (June 2004). "Roles of the two Drosophila CRYPTOCHROME structural domains in circadian photoreception". Science 304 (5676): 1503–6. Bibcode 2004Sci...304.1503B. doi:10.1126/science.1096973. PMID 15178801.
- ^ Dunlap JC (January 1999). "Molecular bases for circadian clocks". Cell 96 (2): 271–90. doi:10.1016/S0092-8674(00)80566-8. PMID 9988221.
- ^ Fogle KJ, Parson KG, Dahm NA, Holmes TC (March 2011). "CRYPTOCHROME is a blue-light sensor that regulates neuronal firing rate". Science 331 (6023): 1409–13. Bibcode 2011Sci...331.1409F. doi:10.1126/science.1199702. PMID 21385718.
- ^ a b c Sancar A, Lindsey-Boltz LA, Kang TH, Reardon JT, Lee JH, Ozturk N (June 2010). "Circadian clock control of the cellular response to DNA damage". FEBS Lett. 584 (12): 2618–25. doi:10.1016/j.febslet.2010.03.017. PMC 2878924. PMID 20227409. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2878924.
- ^ a b Miyamoto Y, Sancar A (May 1998). "Vitamin B2-based blue-light photoreceptors in the retinohypothalamic tract as the photoactive pigments for setting the circadian clock in mammals". Proc. Natl. Acad. Sci. U.S.A 95 (11): 6097–102. Bibcode 1998PNAS...95.6097M. doi:10.1073/pnas.95.11.6097. PMC 27591. PMID 9600923. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=27591.
- ^ Selby CP, Thompson C, Schmitz TM, Van Gelder RN, Sancar A (December 2000). "Functional redundancy of cryptochromes and classical photoreceptors for nonvisual ocular photoreception in mice". Proc. Natl. Acad. Sci. U.S.A. 97 (26): 14697–702. Bibcode 2000PNAS...9714697S. doi:10.1073/pnas.260498597. PMC 18981. PMID 11114194. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=18981.
- ^ Hattar S, Liao HW, Takao M, Berson DM, Yau KW (February 2002). "Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity". Science 295 (5557): 1065–70. Bibcode 2002Sci...295.1065H. doi:10.1126/science.1069609. PMC 2885915. PMID 11834834. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2885915.
- ^ Hoang N, Schleicher E, Kacprzak S, Bouly JP, Picot M, Wu W, Berndt A, Wolf E, Bittl R, Ahmad M (July 2008). Schibler, Ueli. ed. "Human and Drosophila cryptochromes are light activated by flavin photoreduction in living cells". PLoS Biol. 6 (7): e160. doi:10.1371/journal.pbio.0060160. PMC 2443192. PMID 18597555. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2443192.
- ^ Sato TK, Yamada RG, Ukai H, Baggs JE, Miraglia LJ, Kobayashi TJ, Welsh DK, Kay SA, Ueda HR, Hogenesch JB (12 February 2006). "Feedback repression is required for mammalian circadian clock function". Nature Genetics 38 (3): 312–19. doi:10.1038/ng1745. PMC 1994933. PMID 16474406. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1994933.
- ^ a b Ukai-Tadenuma M, Yamada RG, Xu H, Ripperger JA, Liu AC, Ueda H R (1 January 2011). "Delay in feedback repression by cryptochrome 1 is required for circadian clock function". Cell 144 (2): 268–81. doi:10.1016/j.cell.2010.12.019. PMID 21236481.
- ^ Heyers D, Manns M, Luksch H, Güntürkün O, Mouritsen H (2007). Iwaniuk, Andrew. ed. "A visual pathway links brain structures active during magnetic compass orientation in migratory birds". PLoS ONE 2 (9): e937. Bibcode 2007PLoSO...2..937H. doi:10.1371/journal.pone.0000937. PMC 1976598. PMID 17895978. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1976598.
- ^ Gegear RJ, Casselman A, Waddell S, Reppert SM (August 2008). "Cryptochrome mediates light-dependent magnetosensitivity in Drosophila". Nature 454 (7207): 1014–18. Bibcode 2008Natur.454.1014G. doi:10.1038/nature07183. PMC 2559964. PMID 18641630. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2559964.
- ^ Ahmad M, Galland P, Ritz T, Wiltschko R, Wiltschko W (February 2007). "Magnetic intensity affects cryptochrome-dependent responses in Arabidopsis thaliana". Planta 225 (3): 615–24. doi:10.1007/s00425-006-0383-0. PMID 16955271. Lay summary – Centre national de la recherche scientifique.
- ^ Rodgers CT, Hore PJ (January 2009). "Chemical magnetoreception in birds: the radical pair mechanism". Proc. Natl. Acad. Sci. U.S.A. 106 (2): 353–60. Bibcode 2009PNAS..106..353R. doi:10.1073/pnas.0711968106. PMC 2626707. PMID 19129499. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2626707.
- ^ Biskup T, Schleicher E, Okafuji A, Link G, Hitomi K, Getzoff ED, Weber S (2009). "Direct observation of a photoinduced radical pair in a cryptochrome blue-light photoreceptor". Angew. Chem. Int. Ed. Engl. 48 (2): 404–47. doi:10.1002/anie.200803102. PMID 19058271.
- ^ Muller P, Ahmad M (April 2011). "Light activated cryptochrome reacts with molecular oxygen to form a flavin-superoxide radical pair consistent with magnetoreception". J. Biol. Chem. 286 (24): 21033–21040. doi:10.1074/jbc.M111.228940. PMID 21467031.
- ^ Chandler D, Ilia Solov'yov I, Schulten K. "Cryptochrome and Magnetic Sensing". Beckman Institute for Advanced Science and Technology, University of Illinois Urbana–Champaign. http://www.ks.uiuc.edu/Research/cryptochrome/. Retrieved 2011-04-14.
External links
- MeSH cryptochrome
- Cryptochrome circadian clock in Monarch Butterflies, by Steven M. Reppert, Department of Neurobiology, University of Massachusetts
- Cryptochrome and Magnetic Sensing, Theoretical and Computational Biophysics Group at the University of Illinois at Urbana-Champaign
- 2IJG at the Protein Data Bank; 3-D structure of Arabidopsis cryptochrome 3, obtained by X-ray crystallography.
- Animated model of Murine circadian pathway, including role of Cry
Categories:- Human proteins
- Physiology
- Pigments
- Sensory receptors
Wikimedia Foundation. 2010.