- P2X purinoreceptor
-
ATP P2X receptor Identifiers Symbol P2X_receptor Pfam PF00864 InterPro IPR001429 PROSITE PDOC00932 TCDB 1.A.7 OPM family 220 OPM protein 3h9v Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary P2X receptors are a family of cation-permeable ligand gated ion channels that open in response to the binding of extracellular adenosine 5'-triphosphate (ATP). They belong to a larger family of receptors known as the purinergic receptors. P2X receptors are present in a diverse array of organisms including humans, mouse, rat, rabbit, chicken, zebrafish, bullfrog, fluke, and amoeba.[1]
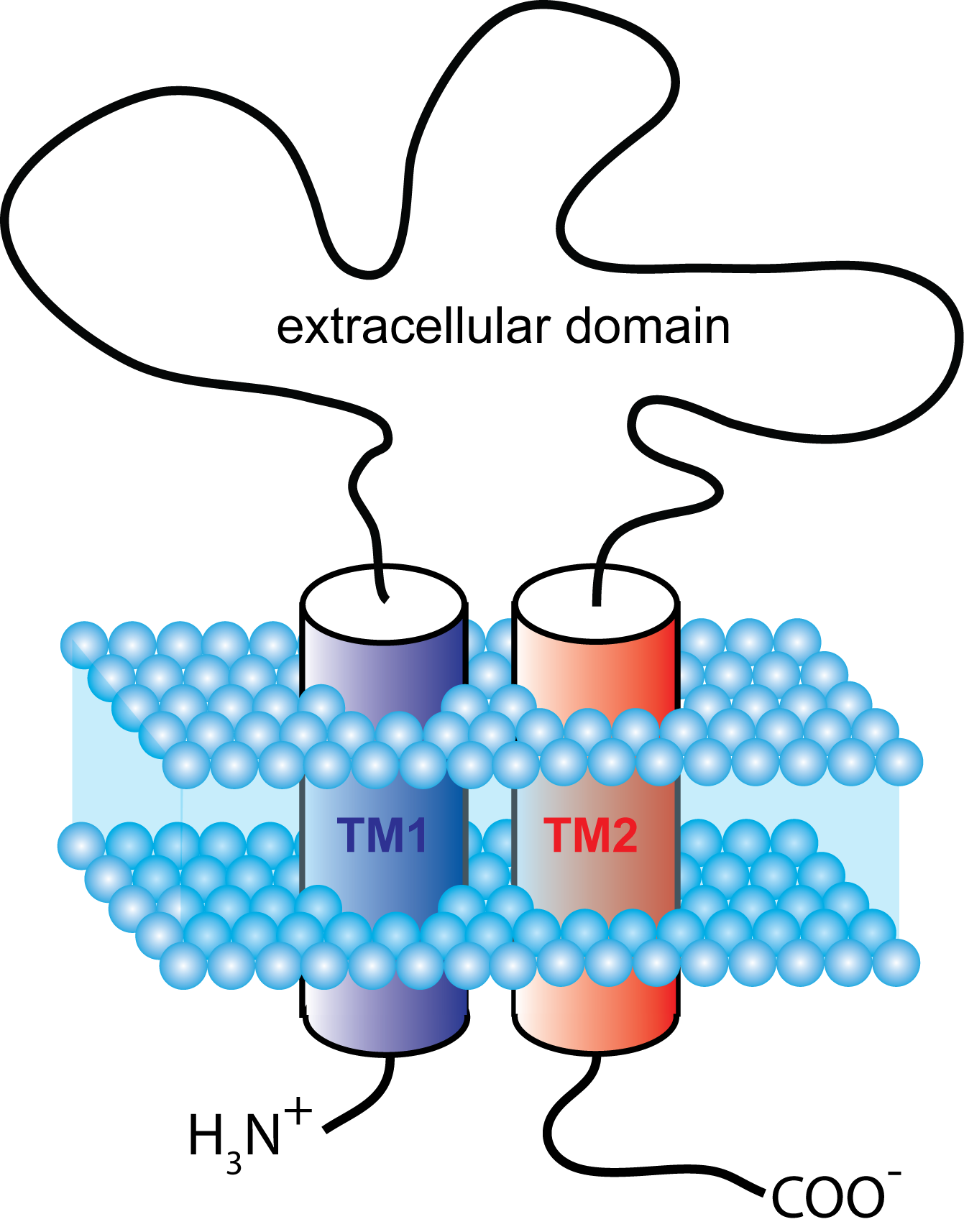
 Figure 1. Schematic representation showing the membrane topology of a typical P2X receptor subunit. First and second transmembrane domains are labeled TM1 and TM2.
Figure 1. Schematic representation showing the membrane topology of a typical P2X receptor subunit. First and second transmembrane domains are labeled TM1 and TM2.
Contents
Basic structure and nomenclature
To date, seven separate genes coding for P2X subunits have been identified, and named to as P2X1 through P2X7.[1][2]
receptor subtype HUGO gene name chromosomal location P2X1 P2RX1 17p13.3 P2X2 P2RX2 12q24.33 P2X3 P2RX3 11q12 P2X4 P2RX4 12q24.32 P2X5 P2RX5 17p13.3 P2X6 P2RX6 22p11.21 P2X7 P2RX7 12q24 The subunits all share a common topology, possessing two plasma membrane spanning domains, a large extracellular loop and intracellular carboxyl and amino termini (Figure 1). With the exception of P2X6, each subunit can readily form a functional homomeric receptor [3]. Receptor nomenclature dictates that naming is determined by the constituent subunits; e.g. a homomeric P2X receptor made up of only P2X1 subunits is called a P2X1 receptor, and a heteromeric receptor containing P2X2 and P2X3 subunits is called a P2X2/3 receptor. The general consensus is that P2X6 cannot form a functional homomeric receptor when expressed alone, but nevertheless can co-assemble with other subunits to form functional heteromeric receptors. Current data suggests that, all of the P2X subunits are capable of forming heteromeric P2X receptors with at least one other subunit type.
Evidence from early molecular biological and functional studies has strongly indicated that the functional P2X receptor protein is a trimer, with the three peptide subunits arranged around an ion-permeable channel pore.[4] This view was recently confirmed by the use of X-ray crystallography to resolve the three-dimensional structure of the zebrafish P2X4 receptor [5](Figure 2).
The relationship between the structure and function of P2X receptors has been the subject of considerable research, and key protein domains responsible for regulating ATP binding, ion permeation, pore dilation and desensitization have been identified.[6][7]
Pharmacology
The pharmacology of a given P2X receptor is largely determined by its subunit makeup.[2] Different subunits exhibit different sensitivities to purinergic agonists such as ATP, α,β-meATP and BzATP; and antagonists such as pyridoxalphosphate-6-azophenyl-2',4'-disulphonic acid (PPADS) and suramin.[1] Of continuing interest is the fact that some P2X receptors (P2X2, P2X4, human P2X5, and P2X7) exhibit multiple open states in response to ATP, characterized by a time-dependent increase in the permeabilities of large organic ions such as N-methyl-D-glucamine (NMDG+) and nucleotide binding dyes such as propidium iodide (YO-PRO-1). Whether this change in permeability is due to a widening of the P2X receptor channel pore itself or the opening of a separate ion-permeable pore is the subject of continued investigation.
Tissue distribution
P2X receptors are expressed in cells from a wide variety of animal tissues. On presynaptic and postsynaptic nerve terminals throughout the central, peripheral and autonomic nervous systems, P2X receptors have been shown to modulate synaptic transmission.[1][8] Furthermore, P2X receptors are able to initiate contraction in cells of the heart muscle, skeletal muscle, and various smooth muscle tissues, including that of the vasculature, vas deferens and urinary bladder. P2X receptors are also expressed on leukocytes, including lymphocytes and macrophages, and are present on blood platelets. There is some degree of subtype specificity as to which P2X receptor subtypes are expressed on specific cell types, with P2X1 receptors being particularly prominent in smooth muscle cells, and P2X2 being widespread throughout the autonomic nervous system. However, such trends are very general and there is considerable overlap in subunit distribution, with most cell types expressing more than one subunits. For example, P2X2 and P2X3 subunits are commonly found co-expressed in sensory neurons, where they often co-assemble into functional P2X2/3 receptors.
Physiological roles
In keeping with their wide distribution throughout the body, P2X receptors are involved in a variety of physiological processes,[1][9] including:
- Modulation of cardiac rhythm and contractility[10]
- Modulation of vascular tone[1]
- Mediation of nociception[11] - e.g. hypersensitivity to innocuous stimuli following upregulation of P2X4 in the spinal cord
- Contraction of the vas deferens during ejaculation[1]
Activation and channel opening
ATP binds to the extracellular loop of the P2X receptor, whereupon it evokes a conformational change in the structure of the ion channel that results in the opening of the ion-permeable pore. This allows cations such as Na+ and Ca2+ to enter the cell, leading to depolarization of the cell membrane and the activation of various Ca2+-sensitive intracellular processes. The channel opening time is dependent upon the subunit makeup of the receptor. For example, P2X1 and P2X3 receptors desensitize rapidly (a few hundred milliseconds) in the continued presence of ATP, whereas the P2X2 receptor channel remains open for as long as ATP is bound to it. Three ATP molecules are thought to be required to activate a P2X receptor, suggesting that ATP needs to bind to each of the three subunits in order to open the channel pore, though recent evidence suggests that ATP binds at the three subunit interfaces.[12] The precise mechanism by which the binding of ATP leads to the opening of the P2X receptor channel pore is not well understood, but is currently under investigation.[6]
Allosteric modulation
The sensitivity of P2X receptors to ATP is strongly modulated by changes in extracellular pH and by the presence of heavy metals (e.g. zinc and cadmium). For example, the ATP sensitivity of P2X1, P2X3 and P2X4 receptors is attenuated when the extracellular pH<7, whereas the ATP sensitivity of P2X2 is significantly increased. On the other hand, zinc potentiates ATP-gated currents through P2X2, P2X3 and P2X4, and inhibits currents through P2X1. The allosteric modulation of P2X receptors by pH and metals appears to be conferred by the presence of histidine side chains in the extracellular domain.[1] In contrast to the other members of the P2X receptor family, P2X4 receptors are also very sensitive to modulation by the macrocyclic lactone, ivermectin.[13] Ivermectin potentiates ATP-gated currents through P2X4 receptors by increasing the open probability of the channel in the presence of ATP, which it appears to do by interacting with the transmembrane domains from within the lipid bilayer.[14]
Subfamilies
- P2RX1 IPR003044
- P2RX2 IPR003045
- P2RX3 IPR003046
- P2RX4 IPR003047
- P2RX5 IPR003048
- P2RX6 IPR003049
- P2RX7 IPR003050
Human proteins containing this domain
P2RX1; P2RX2; P2RX3; P2RX4; P2RX5; P2RX7; P2RXL1; TAX1BP3
See also
References
- ^ a b c d e f g h North RA (2002). "Molecular physiology of P2X receptors". Physiol. Rev. 82 (4): 1013–67. doi:10.1152/physrev.00015.2002 (inactive 2010-08-02). PMID 12270951.
- ^ a b Gever JR, Cockayne DA, Dillon MP, Burnstock G, Ford AP (2006). "Pharmacology of P2X channels". Pflugers Arch. 452 (5): 513–37. doi:10.1007/s00424-006-0070-9. PMID 16649055.
- ^ Torres GE, Egan TM, Voigt MM (1999). "Hetero-oligomeric assembly of P2X receptor subunits. Specificities exist with regard to possible partners.". J. Biol. Chem. 274 (10): 6653–9. doi:10.1074/jbc.274.10.6653. PMID 10037762.
- ^ Nicke A, Baumert HG, Rettinger J, Eichele A, Lambrecht G, Mutschler E, Schmalzing G (1998). "P2X1 and P2X3 receptors form stable trimers: a novel structural motif of ligand-gated ion channels". EMBO J. 17 (11): 3016–28. doi:10.1093/emboj/17.11.3016. PMC 1170641. PMID 9606184. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1170641.
- ^ Kawate T, Michel JC, Birdsong WT, Gouaux E. (2009). "Crystal structure of the ATP-gated P2X4 ion channel in the closed state". Nature 460 (7255): 592–598. doi:10.1038/nature08198. PMC 2720809. PMID 19641588. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2720809.
- ^ a b Egan TM, Samways DS, Li Z (2006). "Biophysics of P2X receptors". Pflugers Arch. 452 (5): 501–12. doi:10.1007/s00424-006-0078-1. PMID 16708237.
- ^ Roberts JA, Vial C, Digby HR, Agboh KC, Wen H, Atterbury-Thomas A, Evans RJ (2006). "Molecular properties of P2X receptors". Pflugers Arch. 452 (5): 486–500. doi:10.1007/s00424-006-0073-6. PMID 16607539.
- ^ Burnstock G (2000). "P2X receptors in sensory neurones". Br J Anaesth 84 (4): 476–88. PMID 10823099. http://bja.oxfordjournals.org/cgi/content/abstract/84/4/476.
- ^ Khakh BS, North RA (2006). "P2X receptors as cell-surface ATP sensors in health and disease". Nature 442 (7102): 527–32. doi:10.1038/nature04886. PMID 16885977.
- ^ Vassort G (2001). "Adenosine 5'-triphosphate: a P2-purinergic agonist in the myocardium". Physiol. Rev. 81 (2): 767–806. PMID 11274344.
- ^ Chizh BA, Illes P (2001). "P2X receptors and nociception". Pharmacol. Rev. 53 (4): 553–68. PMID 11734618. http://pharmrev.aspetjournals.org/cgi/content/abstract/53/4/553.
- ^ Evans RJ (2008). "Orthosteric and allosteric binding sites of P2X receptors". Eur. Biophys. J. 38 (3): 319–27. doi:10.1007/s00249-008-0275-2. PMID 18247022.
- ^ Khakh BS, Proctor W, Dunwiddie TV, Labarca C, Lester HA (1999). "Allosteric control of gating and kinetics at P2X(4) receptor channels". J. Neurosci. 19 (17): 7289–99. PMID 10460235.
- ^ Priel A, Silberberg SD (2004). "Mechanism of ivermectin facilitation of human P2X4 receptor channels". J. Gen. Physiol. 123 (3): 281–93. doi:10.1085/jgp.200308986. PMC 2217454. PMID 14769846. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2217454.
External links
- Ivar von Kügelgen: Pharmacology of mammalian P2X- and P2Y-receptors, BIOTREND Reviews No. 03, September 2008,© 2008 BIOTREND Chemicals AG
- Ligand-gated ion channel Database (European Bioinformatics Institute)
Ion channel, cell surface receptor: ligand-gated ion channels Cys-loop receptors monomers: α1 · α2 · α3 · α4 · α5 · α6 · α7 · α9 · α10 · β1 · β2 · β3 · β4 · δ · ε
pentamers: (α3)2(β4)3 · (α4)2(β2)3 · (α7)5 · (α1)2(β4)3 - Ganglion type · (α1)2β1δε - Muscle typeZincZinc-activatedIonotropic glutamates Ligand-gated onlyVoltage- and ligand-gated‘Orphan’ATP-gated channels Categories:- Ion channels
- Ionotropic receptors
- Cell signaling
- Molecular neuroscience

Wikimedia Foundation. 2010.