- Cycad
-
Cycadophyta
Temporal range: Early Permian–Recent
Cycas rumphii with old and new male cones. Scientific classification Kingdom: Plantae Division: Cycadophyta
Bessey 1907: 321.[1]Class: Cycadopsida
Brongn.[2]Order: Cycadales
Dumort.Families Cycadaceae cycas family
Stangeriaceae stangeria family
Zamiaceae zamia familyCycads /ˈsaɪkædz/ are seed plants typically characterized by a stout and woody (ligneous) trunk with a crown of large, hard and stiff, evergreen leaves. They usually have pinnate leaves. The individual plants are either all male or all female (dioecious). Cycads vary in size from having a trunk that is only a few centimeters tall to trunks up to several meters tall. They typically grow very slowly and live very long, with some specimens known to be as much as 1,000 years old. Because of their superficial resemblance, they are sometimes confused with and mistaken for palms or ferns, but are only distantly related to either.
Cycads are found across much of the subtropical and tropical parts of the world. They are found in South and Central America (where the greatest diversity occurs), Mexico, the Antilles, southeastern United States, Australia, Melanesia, Micronesia, Japan, China, Southeast Asia, India, Sri Lanka, Madagascar, and southern and tropical Africa, where at least 65 species occur. Some can survive in harsh semidesert climates (xerophytic), others in wet rain forest conditions, and some in both.[citation needed] Some can grow in sand or even on rock, some in oxygen-poor swampy bog-like soils rich in organic material, and some in both.[citation needed] Some are able to grow in full sun, some in full shade, and some in both.[citation needed] Some are salt tolerant (halophytes).
Cycads belong to the biological division Cycadophyta. There are three extant families of cycads, Cycadaceae, Stangeriaceae, and Zamiaceae. Though they are a minor component of the plant kingdom today, during the Jurassic period they were extremely common. They have changed little since the Jurassic, compared to some major evolutionary changes in other plant divisions.
Cycads are gymnosperms (naked seeded), meaning that their unfertilized seeds (ovules) are open to the air to be directly fertilized by pollination, as contrasted with angiosperms, who have enclosed seeds with more complex fertilization arrangements. Cycads have very specialized pollinators, usually a specific kind of beetle. They have been reported to fix nitrogen in association with a cyanobacterium living in the roots. These blue-green algae produce a neurotoxin called BMAA that is found in the seeds of cycads. A neurotoxin cycling through the food chain from the cyanobacterium through cycads to its seeds, to bats eating the seeds, to humans consuming the bats, is hypothesized to be a source for some neurological diseases in humans, such as mad cow disease.[3]
Contents
Origins
The cycad fossil record dates to the early Permian, 280 mya (million years ago). There is controversy over older cycad fossils that date to the late Carboniferous period, 300–325 mya. One of the first colonizers of terrestrial habitats, this clade probably diversified extensively within its first few million years, although the extent to which it radiated is unknown because relatively few fossil specimens have been found. The regions to which cycads are restricted probably indicate their former distribution in the Pangea before the supercontinents Laurasia and Gondwana separated (Hermsen et al. 2006). Recent studies have indicated that the common perception of existing cycad species as living fossils is largely misplaced, with only Bowenia dating to the Cretaceous or earlier. Although the cycad lineage itself is ancient, most extant species have evolved in the last 12 million years.[4]
The family Stangeriaceae (named for Dr. William Stanger, 1812(?)–1854), consisting of only three extant species, is thought to be of Gondwanan origin, as fossils have been found in Lower Cretaceous deposits in Argentina, dating to 70–135 mya. The family Zamiaceae is more diverse, with a fossil record extending from the middle Triassic to the Eocene (54–200 mya) in North and South America, Europe, Australia, and Antarctica, implying that the family was present before the break-up of Pangea. The family Cycadaceae is thought to be an early offshoot from other cycads, with fossils from Eocene deposits (38–54 mya) in Japan and China, indicating that this family originated in Laurasia. Cycas is the only genus in the family and contains 99 species, the most of any cycad genus. Molecular data have recently shown that Cycas species in Australasia and the east coast of Africa are recent arrivals, suggesting adaptive radiation may have occurred. The current distribution of cycads may be due to radiations from a few ancestral types sequestered on Laurasia and Gondwana, or could be explained by genetic drift following the separation of already evolved genera. Both explanations account for the strict endemism across present continental lines.
 Leaves and cone of Encephalartos sclavoi
Leaves and cone of Encephalartos sclavoi
 The fossil Cycad Zamites feneonis
The fossil Cycad Zamites feneonisTaxonomy
There are about 305 described species, in 10–12 genera and 2–3 families of cycads (depending on taxonomic viewpoint). The classification below, proposed by Dennis Stevenson in 1992, is based upon a hierarchical structure based on cladistic analyses of morphological, anatomical, karyological, physiological and phytochemical data.
The number of species in the clade is low compared to the number of species in most other plant phyla. However, paleobotanical and molecular research indicates that diversity was greater in the history of the phylum. Fossil evidence shows that structural diversity in Mesozoic cycad pollen "considerably exceeds that seen in surviving genera today". The impacts of extinction on diversity are highlighted below. The disparity in molecular sequences is very high between the three main lineages of cycads, implying that genetic diversity in the clade was once high, but this fact has led to major disagreements about the divisions within the Cycadales.
The number of described cycad species has doubled in the past 25 years, mostly due to improved sampling and further exploration. Experts infer there may still be about 100 undescribed species, based on the rate of discovery. These are likely to be in Asia and South America, where areas of endemism are highest. Diversity hotspots also occur in Australia, South Africa, Mexico, China and Vietnam, which together account for more than 70% of the world’s cycad species. The taxonomy of the Cycadophyta is, however, now stabilizing.
Cycad systematists reject the biological/isolation species concept, because some clearly defined cycad species can interbreed and produce fertile offspring; this character is thus not disproportionately weighted when determining species barriers. The phenetic species concept, which states that a species is defined based on overall similarities with other individuals of the same species combined with a significant gap in variation with other species, is also rejected. Most cycad taxonomists agree on a modified version of the evolutionary species concept, The classification below is taken from Stevenson (1992).
- Suborder Cycadineae
- Family Cycadaceae
- Subfamily Cycadoideae
-
-
- Cycas. About 105 species in the Old World from Africa east to southern Japan, Australia and the western Pacific Ocean islands; type: C. circinalis L.; see also C. pruinosa and C. revoluta
-
-
- Subfamily Cycadoideae
- Suborder Zamiineae
- Family Stangeriaceae
- Subfamily Stangerioideae
-
-
- Stangeria. One species in southern Africa; type: S. eriopus (Kunze) Baillon
-
-
- Subfamily Bowenioideae
-
-
- Bowenia. Two species in Queensland, Australia; type: B. spectabilis Hook. ex Hook. f.
-
-
- Subfamily Stangerioideae
- Family Zamiaceae
- Subfamily Encephalartoideae
- Tribe Diooeae
- Tribe Encephalarteae
- Subtribe Encephalartinae
- Encephalartos. About 66 species in southeast Africa; type: E. friderici-guilielmi Lehmann, E. transvenosus (Modjadji cycad)
- Subtribe Macrozamiinae
- Macrozamia. About 41 species in Australia; type: M. riedlei (Fischer ex Gaudichaud) C.A. Gardner
- Lepidozamia. Two species in eastern Australia; type: L. peroffskyana Regel
- Subtribe Encephalartinae
- Subfamily Zamioideae
- Tribe Ceratozamieae
-
- Ceratozamia. 26 species in southern Mexico and Central America; type: C. mexicana Brongn.
-
- Tribe Zamieae
- Subtribe Microcycadinae
- Microcycas. One species in Cuba; type: M. calocoma (Miquel) A. DC.
- Subtribe Zamiinae
- Subtribe Microcycadinae
- Tribe Ceratozamieae
- Subfamily Encephalartoideae
Cycads are most closely related to the extinct Bennettitales, and are also relatively close relatives to the Ginkgoales, as shown in the following phylogeny (Crepet 2000):
Cycads
Ginkgo
Conifers
Anthophytes Bennettitales
Gnetales
Angiosperms
Angiosperms
Cycads
Bennettitales
Ginkgo
Conifers
Gnetales
Traditional view Modern view Identification
Cycads have a cylindrical trunk which usually does not branch. Leaves grow directly from the trunk, and typically fall when older, leaving a crown of leaves at the top. The leaves grow in a rosette form, with new foliage emerging from the top and center of the crown. The trunk may be buried, so the leaves appear to be emerging from the ground, so the plant appears to be a basal rosette. The leaves are generally large in proportion to the trunk size, and sometimes even larger than the trunk.
The leaves are pinnate (in the form of a birds feather, pinnae), with a central leaf stalk from which parallel "ribs" emerge from each side of the stalk, perpendicular to it. The leaves are typically either compound (the leaf stalk has leaflets emerging from it as "ribs"), or have edges (margins) that are so deeply cut (incised) as to appear compound. Some species have leaves that are bipinnate, which means that the leaflets each have their own sub-leaflets, growing in the same form on the leaflet as the leaflets grow on the stalk of the leaf (self similar geometry).
The three families can be identified by looking at the central stalk of the leaf. Each family has at least one vein running up the leaf stalk from bottom to top (longitudinal). In Cycadaceae, there is only one vein in the center of the leaf stalk (central vein), but there are no veins on the stalklets of the leaflet (no lateral veins). In Stangeriaceae, there is also only one central vein, but there are also lateral veins. In Zamiaceae, there are more than one central veins, and they are parallel to each other.
History
Modern knowledge about cycads began in the 9th century with the recording by two Arab naturalists that the genus Cycas was used as a source of flour in India. Later, in the 16th century, Antonio Pigafetta, Fernao Lopez de Castanheda and Francis Drake found Cycas plants in the Moluccas, where the seeds were eaten. The first report of cycads in the New World was by Giovanni Lerio in his 1576 trip to Brazil, where he observed a plant named ayrius by the indigenous people; this species is now classified in the genus Zamia.
Cycads belonging to the genus Encephalartos were first described by Johann Georg Christian Lehmann in 1834. The name is derived from the Greek articles "en", meaning "in", "cephale", meaning "head", and "artos", meaning "bread".
Throughout the 18th-19th centuries, discoveries of several species were reported by numerous naturalist researchers and discoverers traveling throughout the world. One of the most notable researchers of cycads was American botanist C.J. Chamberlain whose work is noteworthy for the quantity of data and the novelty of his approach to studying cycads. His 15 years of travel throughout Africa, the Americas and Australia to observe cycads in their natural habitat resulted in his 1919 publication of The Living Cycads which remains current in its synthesis of taxonomy, morphology and reproductive biology of cycads, most of which was obtained from his original research. His 1940s monograph on the Cycadales, though never published (most likely because of his death) was never used by botanists. The most recent complete work on the cycads is the book by Norstog and Nicholls entitled "the Biology of the Cycads" published in 1998.
Uses

The starch obtained from the stems of certain species is still used as food by some indigenous tribes. Tribal people also grind and soak the seeds to remove the nerve toxins that may be present, making the food source generally safe to eat, although often not all the toxin is removed. In addition, consumers of bush meat may face a health threat as the meat comes from game which may have eaten cycad seeds and carry traces of the toxin in body fat.
Cycad, known as sotetsu (Jap. ソテツ, Kanji: 蘇鉄) in Japanese, was traditionally a famine food in Okinawa - a last resort to turn to for sustenance during particularly difficult times.[5] A period of particularly devastating poverty and famine in the 1920s, caused in large part by Japanese economic policies in the island prefecture, is known as "cycad hell" or sotetsu jigoku.[6]
Cycad meal known as Eenthu in Malayalam is a common food in Kerala. Traditionally, the seeds were sliced and kept in direct sunlight or near the hearth during rainy season to promote drying. The drying process is carried out to reduce the toxin levels and as a means of preservation. The outer shell is subsequently removed and inner portion is ground into a flour. Properly dried cycad seed flour may be stored for several years without deterioration.
Food items like Puttu,Eenthu kanji,Eenthu payasam etc are made out of cycad seed power.These food items are particularly prepared in heavy rainy seasons in Kerala.
Cycad leaves are used to decorate venues during festivals, marriages and other community celebrations.
There is some indication that the regular consumption of starch derived from cycads is a factor in the development of Lytico-Bodig disease, a neurological disease with symptoms similar to those of Parkinson's disease and ALS. Lytico-Bodic and its potential connection to cycasin ingestion is one of the subjects explored in Oliver Sacks' 1997 book Island of the Colourblind. Cattle that graze in pastures containing cycads may ingest the leaves and seeds and develop the neurologic syndrome of cycad toxicosis known as zamia staggers.
In Vanuatu, where the cycad is known by the Bislama name namele, the tree has deep customary and spiritual significance. A single cycad leaf may be used as a taboo sign, while a pair of crossed cycad leaves is a peace sign and appears on the Vanuatu flag. The breaking off of fronds from a cycad leaf is used in traditional contexts as an aid to counting.
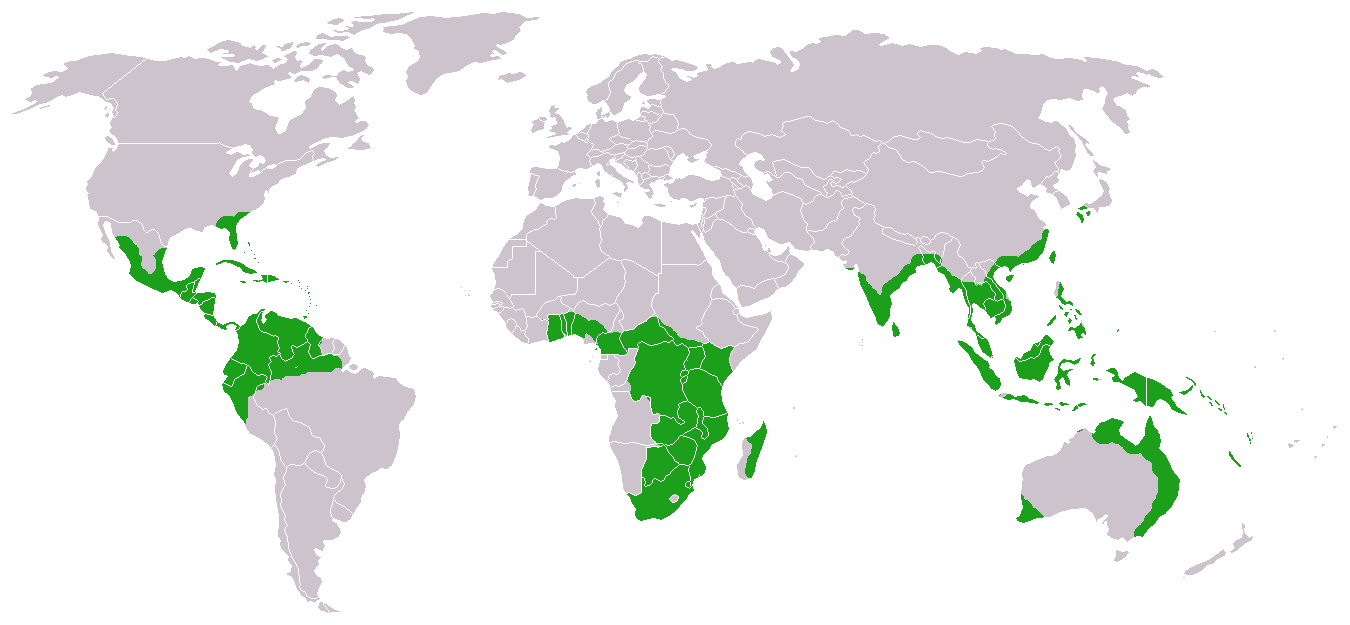
Distribution

Approximate world distribution of living Cycadales Overall species diversity peaks at 17˚ 15"N and 28˚ 12"S, with a minor peak at the equator. There is therefore not a latitudinal diversity gradient towards the equator but towards the tropics. However, the peak in the northern tropics is largely due to Cycas in Asia and Zamia in the New World, whereas the peak in the southern tropics is due to Cycas again, and also to the diverse genus Encephalartos in southern and central Africa and Macrozamia in Australia. Thus, the distribution pattern of cycad species with latitude appears to be an artifact of the geographical isolation of cycad genera, and is dependent on the remaining species in each genus that did not follow the extinction pattern of their ancestors. Cycas is the only genus that has a broad geographical range and can thus be used to infer that cycads tend to live in the upper and lower tropics. This is probably because these areas have a drier climate with relatively cool winters; while cycads require some rainfall, they appear to be partly xerophytic. Potted specimens are found and thrive in global locations such as Canada, Russia, Finland and Chile.
Speciation
There are no documented cases of sympatric speciation in cycads and allopatry appears to be the most common form of speciation in the group. This is difficult to study. as they are long-lived plants, so natural experiments have been investigated. One example is Cycas seemannii, which occurs only in Fiji, New Caledonia, Tonga and Vanuatu. Genetic diversity within populations was found to be significantly lower than between islands, suggesting that genetic drift is a likely mechanism for speciation, and is probably currently occurring between the isolated populations. Allopatry has also been proposed as the mechanism of speciation in Dioon, which predominantly occurs in Mexico. The many rivers that have shaped the region, and repeated glaciation and consequent disjunction, are thought to have been important in reproductive isolation not only in Dioon but in many other plant and animal taxa. Parapatric speciation may also have occurred, especially as cycads are pollinated by insects rather than by wind (Stevenson et al. 1998). As the range of the species grows, the individuals furthest apart are prevented from interbreeding, as insects have relatively small ranges and will not pollinate between these plants. If sympatric speciation has occurred in cycads, this would most likely be because of a host shift in pollinators, due to the very fact that cycads are uniformly dioecious.
Extinction
The probable former range of cycads can be inferred from their global distribution. For example, the family Stangeriaceae only contains three extant species in Africa and Australia. Diverse fossils of this family have been dated to 135 mya, indicating that diversity may have been much greater before the Jurassic and late Triassic mass extinction events. However, the cycad fossil record is generally poor and little can be deduced about the effects of each mass extinction event on their diversity.
Instead, correlations can be made between the number of extant gymnosperms and angiosperms. It is likely that cycad diversity was affected more by the great angiosperm radiation in the mid-Cretaceous than by extinctions. Very slow cambial growth was first used to define cycads, and because of this characteristic the group could not compete with the rapidly growing, relatively short-lived angiosperms, which now number over 250,000 species, compared to the 947 remaining gymnosperms. It is surprising that the cycads are still extant, having been faced with extreme competition and five major extinctions. The ability of cycads to survive in relatively dry environments where plant diversity is generally lower, and their great longevity, may explain their long persistence.
Conservation

In recent years, many cycads have been dwindling in numbers and may face risk of extinction because of theft and unscrupulous collection from their natural habitats, as well as from habitat destruction.
About 23% of the 305 extant cycad species are either critically endangered or endangered, and 15% are vulnerable. Thus, 38% of cycads are on the IUCN Red List (2004), and the other 62% are in the Least concern or Near Threatened category (i.e. not actually on the Red List), or are data deficient. This value has changed dramatically within the past few years; 46% of cycads were on the 1978 Red List, and this rose to 82% in 1997. This was largely due to the recent discovery of over 150 new species, disagreements about classification, and uncertainty. This has not been helpful for conservation planning for the group.
Zamia in the New World, Cycas in Asia and Encephalartos in Africa are the most threatened genera. This pattern reflects the pressures on species in these regions. At least two species, Encephalartos woodii and Encephalartos relictus (both from Africa), are confirmed extinct in the wild. Cycads are long-lived with infrequent reproduction, and most populations are small, putting them at risk of extinction from habitat destruction and stochastic environmental events. Regionally, Australian cycads are the least at risk, as they are locally common and habitat fragmentation is low. However, land management with fire is thought to be a threat to Australian species. African cycads are rare and are thought to be naturally decreasing due to small population sizes, and there is controversy over whether to let natural extinction processes act on these cycads.
All cycads are in the CITES appendix appearing under the heading Plant Kingdom and under three family names, Cycadaceae, Stangeriaceae and Zamiaceae.
All cycads are CITES APPENDIX II except the following, in APPENDIX I:
- Cycas beddomei
- Stangeria eriopus
- All Ceratozamia
- All Chigua
- All Encephalartos
- Microcycas calocoma
Cycad seeds from species on APPENDIX II are not CITES regulated. APPENDIX I seeds are treated the same as the plants.
As of 2008, only two known specimens were known to be extant, both males. Cycad expert Loran Whitelock, author of The Cycads, was doing experiments on cloning with one of the specimens, to try to preserve this species.[citation needed]
Horticulture
 A Sago Cycad (Cycas revoluta) growing in England as a houseplant
A Sago Cycad (Cycas revoluta) growing in England as a houseplantCycads can be cut into pieces to make new plants, or by direct planting of the seeds. Propagation by seeds is the preferred method of growth, and two unique risks to their germination exist. One is that the seeds have no dormancy, so the embryo is biologically required to maintain growth and development, which means if the seed dries out, it dies. The second is that the emerging radicle and embryo can be very susceptible to fungal diseases in its early stages, when in unhygienic or excessively wet conditions. Thus, many cycad growers pregerminate the seeds in moist, sterile media such as vermiculite or perlite. However pregermination is not necessary, and many report success by directly planting the seeds in regular potting soil. As with many plants, a combination of well-drained soil, sunlight, water and nutrients will help it to prosper. Although, because of their hardy nature, cycads do not necessarily require the most tender or careful treatment, they can grow in almost any medium, including soilless ones. One of the most common causes of cycad death is from rotting stems and roots due to over-watering.
Some insects, particularly scale insects, some weevils and chewing insects can damage cycads, though the pests are susceptible to insecticides such as the horticulture soluble oil white oil. Sometimes bacterial preparations may be used to control insect infestation on cycads. When some of the mature plants prepare for reproduction, though, the presence of weevils has been shown to help accomplish pollination.
While the cycads have a reputation of slow growth, it is not always well-founded, and some actually grow quite fast, achieving reproductive maturity in 2–3 years (as with some Zamia species), while others in 15 years (as with some Cycas, Australian Macrozamia and Lepidozamia).
See also
References
- ^ Bessey, C.E. (1907). "A synopsis of plant phyla". Nebraska Univ. Stud. 7: 275–373.
- ^ Brongniart, A. (1843). Énumération des genres de plantes cultivées au Musée d'histoire naturelle de Paris.
- ^ The Tangle, Jonathan Weiner, Annals of Medicine, The New Yorker, 4-11-2005, [1]
- ^ N. S. Nagalingum et al., Science, [2]
- ^ Kerr, George. Okinawa: The History of an Island People. Revised Edition. Boston: Tuttle Publishing, 2000. pp406-7.
- ^ Smits, Gregory. "Okinawa in Postwar Japanese Politics and the Economy." East-Asian-History.net. Accessed 5 December 2009.
- Chamberlain, C.J. (1965) [1919] The Living Cycads, 1st ed. reprinted, New York : Hafner, 172 p., LCCN 65-027079
- Chaw, S.-M., Parkinson, C.L., Cheng, Y., Vincent, T.M., & Palmer, J.D. (2000) "Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifer", Proceedings of the National Academy of Sciences, 97 (8), p. 4086-4091.
- Chaw, S.-M., Walters, T.W., Chang, C.-C., Hu, S.-H., & Chen, S.-H. (2005) "A phylogeny of cycads (Cycadales) inferred from chloroplast matK gene, trnK intron, and nuclear rDNA ITS region", Molecular Phylogenetics and Evolution, 37 (1), p. 214-234. doi:10.1016/j.ympev.2005.01.006
- Chaw, S.-M., Zharkikh, A., Sung, H.-M., Luu, T.-C., & Li, W.-H. (1997) "Molecular phylogeny of extant gymnosperms and seed plant evolution: Analysis of nuclear 18s rRNA sequences", Molecular Biology and Evolution, 14 (1), p. 56-68.
- Crepet, W.L. (2000) "Progress in understanding angiosperm history, success, and relationships: Darwin's abominably 'perplexing phenomenon'", Proceedings of the National Academy of Sciences, 97 (24), p. 12939-12941, doi:10.1073/pnas.97.24.12939
- De Luca, Paulo (1990) "A Historical Perspective on Cycads from Antiquity to the Present", In: Stevenson, D. (Ed.) The Biology, Structure, and Systematics of the Cycadales, Memoirs of the New York Botanical Garden, 57, p. 1-7, ISBN 0-89327-350-3
- Donaldson, J.S. (2003) "Chapter 3: Regional Overview: Africa", In: Donaldson, J.S. (ed.), Cycads. Status Survey and Conservation Action Plan, IUCN/SSC Cycad Specialist Group, Gland; Cambridge : UKIUCN, ISBN 2-8317-0699-8, p. 9-19
- Donaldson, J. (2004) "Saving ghosts? The implications of taxonomic uncertainty and shifting infrageneric concepts in the cycadales for red listing and conservation planning", In: Walters,T. & Osborne, R. (eds), Cycad Classification: Concepts and Recommendations, Wallingford, UK : CABI, ISBN 0-85199-741-4, p. 13-22.
- Donaldson, J., Hill, K.D., & Stevenson, D.W. (2003a) "Chapter 2: Cycads of the World: An Overview", In: Donaldson, J.S. (ed.), Cycads. Status Survey and Conservation Action Plan, IUCN/SSC Cycad Specialist Group, Gland; Cambridge : UKIUCN, ISBN 2-8317-0699-8, p. 3-8
- Donaldson, J.S., Dehgan, A.P., Vovides, A.P., & Tang, W. (2003b) "Chapter 7: Cycads in trade and sustainable use of cycad populations", In: Donaldson, J.S. (ed.), Cycads. Status Survey and Conservation Action Plan, IUCN/SSC Cycad Specialist Group, Gland; Cambridge : UKIUCN, ISBN 2-8317-0699-8, p. 39-47
- Golding, J.S. & Hurter, P.J.H. (2003) "A Red List account of Africa’s cycads and implications of considering life-history and threats", Biodiversity and Conservation, 12 (3), p. 507–528, doi:10.1023/A:1022472801638
- Gonzàlez-Astorga, J., Vovides, A.P., Ferrer, M.M., & Iglesias, C. (2003a) "Population genetics of Dioon edule Lindl. (Zamiaceae, Cycadales): biogeographical and evolutionary implications", Botanical Journal of the Linnean Society, 80 (3), p. 457-467, doi:10.1046/j.1095-8312.2003.00257.x
- Gonzàlez-Astorga, J., Vovides, A.P., & Iglesias, C. (2003b) "Morphological and geographic variation in Dioon edule Lindl. (Zamiaceae): ecological and evolutionary implications", Botanical Journal of the Linnean Society, 141 (4), p. 465-470, doi:10.1046/j.1095-8339.2003.00155.x
- Gregory, T.J. & Chemnick, J. (2004) Hypotheses of the relationship between biogeography and speciation in Dioon (Zamiaceae). In: Walters,T. & Osborne, R. (eds), Cycad Classification: Concepts and Recommendations, Wallingford, UK : CABI, ISBN 0-85199-741-4, p. 137-148.
- Hermsen, E., E. Taylor, T. Taylor, & D. Stevenson. 2006. Cataphylls of the middle Triassic cycad Antarcticycas schopfii and new insights into cycad evolution. American Journal of Botany 93: 724–738. 2006.
- Hill, C.R. (1990) "Ultrastructure of in situ fossil cycad pollen from the English Jurassic, with a description of the male cone Androstrobus balmei sp. nov.", Review of Palaeobotany and Palynology, 65 (1-4), p. 165-193, doi:10.1016/0034-6667(90)90067-S
- Hill, K.D. (2003) "Chapter 4: Regional Overview: Australia", In: Donaldson, J.S. (ed.), Cycads. Status Survey and Conservation Action Plan, IUCN/SSC Cycad Specialist Group, Gland; Cambridge : UKIUCN, ISBN 2-8317-0699-8, p. 20-24
- Hill, K.D. (2004) "Character evolution, species recognition and classification concepts in the cycadaceae", In: Walters,T. & Osborne, R. (eds), Cycad Classification: Concepts and Recommendations, Wallingford, UK : CABI, ISBN 0-85199-741-4, p. 23-44. CABI, Oxford.
- Hill, K. & Stevenson, D. 1998–present. The Cycad Pages. http://plantnet.rbgsyd.nsw.gov.au/PlantNet/cycad/
- Hill, K.D., Stevenson, D.W., & Obsorne, R. (2004) "The World List of Cycads", Botanical Review, 70 (2), p. 274-298. doi:10.1663/0006-8101(2004)070[0274:TWLOC]2.0.CO;2
- Jones, David L. (1993) Cycads of the World, Washington, D.C. : Smithsonian Institution Press, ISBN 1-56098-220-9. 2nd ed. republished (2002) as: Cycads of the World: ancient plants in today's landscape, Sydney : Reed New Holland, ISBN 1-876334-69-X
- Keppel, G., Lee, S.W., & Hodgskiss, P.D. (2002) "Evidence for Long Isolation Among Populations of a Pacific Cycad: Genetic Diversity and Differentiation in Cycas seemannii A.Br. (Cycadaceae)", Journal of Heredity, 93 (2), p. 133-139. doi:10.1093/jhered/93.2.133
- Norstog, K.J. & Nicholls, T.J. (1998) Biology of Cycads, Ithaca : Cornell University Press, ISBN 0-80143-033-X
- Stevenson, D. 1992. A formal classification of the extant cycads. Brittonia 44: 220-223.
- Stevenson, D., K. Norstog, & P. Fawcett. 1998. Pollination biology of cycads. In: Reproductive Biology: In systematics, conservation, and economic botany. Eds. S. Owens & P. Rudall. pp. 277–294. Royal Botanic Gardens, Kew.
Walters, T., Osborne, R., & Decker, D. (2004) "We hold these truths…", In: Walters,T. & Osborne, R. (eds), Cycad Classification: Concepts and Recommendations, Wallingford, UK : CABI, ISBN 0-85199-741-4, p. 1-11
- Whitelock, L.M. (2002) The Cycads, Portland, OR : Timber Press, ISBN 0-88192-522-5
External links
- Palm Trees, Small Palms, Cycads, Bromeliads and tropical plants site with thousands of large, high quality photos of cycads and associated flora. Includes information on habitat and cultivation.
- Hill KD (1998–2004) The Cycad Pages, Royal Botanic Gardens Sydney. http://plantnet.rbgsyd.nsw.gov.au/PlantNet/cycad/index.html
- Gymnosperm Database: Cycads
- Fairchild Tropical Botanic Garden- one of the largest collection of cycads in the world in Florida, U.S.A.
- Palm and Cycad Societies of Australia (PACSOA)
- The Cycad Society of South Africa
- Cycad nitrogen fixation
- Cycad toxicity
- Cycads - Foto
- The Cult of the Cycads, New York Times Magazine article on cycad collectorship and cycad smuggling
- Cycads An annotated link directory
Classification of Archaeplastida / Plantae sensu lato Rhodophyta Cyanidiophyceae · Porphyridiophyceae · Compsopogonophyceae · Stylonematophyceae · Rhodellophyceae · Bangiophyceae · Florideophyceae (Hildenbrandiales, Acrochaetiales, Nemaliales, Batrachospermales, Corallinales, Gelidiales, Gracilariales, Ceramiales)Glaucocystophyceae Glaucocystis · Cyanophora · GloeochaeteViridiplantae/
Plantae
sensu strictoEuphyllophytaMoniliformopses (Equisetopsida, Filicopsida, Psilotopsida)
Spermatophyta: Gymnosperm (Pinophyta, Cycadophyta, Ginkgophyta, Gnetophyta) · MagnoliophytaCategories:- Cycads
- Living fossils
- Dioecious plants






Wikimedia Foundation. 2010.