- Quaternary extinction event
-
 Late Pleistocene landscape of northern Eurasia
Late Pleistocene landscape of northern Eurasia
The Quaternary period saw the extinctions of numerous predominantly larger, especially megafaunal, species, many of which occurred during the transition from the Pleistocene to the Holocene epoch. However, the extinction wave did not stop at the end of the Pleistocene, but continued especially on isolated Islands in Holocene extinctions. Among the main causes hypothesized by paleontologists are natural climate change and overkill by humans, who appeared during the Middle Pleistocene and invaded many previously uninhabited regions of the world during the Late Pleistocene and Holocene. A variant of the latter possibility is the second-order predation hypothesis, which focuses more on the indirect damage caused by overcompetition with nonhuman predators. The spread of disease is also discussed as a possible reason.
Contents
The Pleistocene or Ice Age extinction event
The Late Pleistocene extinction event saw the extinction of many mammals weighing more than 40 kg.
- In North America around 33 of 45 genera of large mammals became extinct.
- In South America 46 of 58
- In Australia 15 of 16
- In Europe 7 of 23
- In Subsaharan Africa only 2 of 44
The extinctions in the Americas entailed the elimination of all the larger (over 100 kg) mammalian species of South American origin, including those that had migrated north in the Great American Interchange. Only in North America, South America and Australia did the extinction occur at family taxonomic levels or higher.
There are two main hypotheses concerning the Pleistocene extinction:
- The animals died off due to climate change: the retreat of the polar ice cap.
- The animals were exterminated by humans: the "prehistoric overkill hypothesis" (Martin, 1967).
There are some inconsistencies between the current available data and the prehistoric overkill hypothesis. For instance, there are ambiguities around the timing of sudden extinctions of Australian megafauna. Biologists note that comparable extinctions have not occurred in Africa and South or Southeast Asia, where the fauna evolved with hominids. Post-glacial megafaunal extinctions in Africa have been spaced over a longer interval.
Evidence supporting the prehistoric overkill hypothesis includes the persistence of certain island megafauna for several millennia past the disappearance of their continental cousins. Ground sloths survived on the Antilles long after North and South American ground sloths were extinct. The later disappearance of the island species correlates with the later colonization of these islands by humans. Similarly, woolly mammoths died out on remote Wrangel Island 7000 years after their mainland extinction. Steller's sea cows also persisted off the isolated and uninhabited Commander Islands for thousands of years after they vanished from continental shores of the north Pacific.[1]
Controversial alternative hypotheses to the theory of human responsibility include the Younger Dryas event and Tollmann's hypothetical bolide, which claim that the extinctions resulted from bolide impact(s). However, such hypotheses would predict an instantaneous, regional extinction(s), and thus cannot account for the planet-wide species losses that occurred over an interval of thousands of years.
Africa and Asia
The Old World tropics have been relatively spared by Pleistocene extinctions. Africa and Asia are the only regions that have megamammals weighing over 1000 kg today. However, during the early, middle and late Pleistocene some large animal forms disappeared from these regions without being replaced by comparable successor species. Climate change has been cited as most likely causing the extinctions in Southeast Asia.[2]
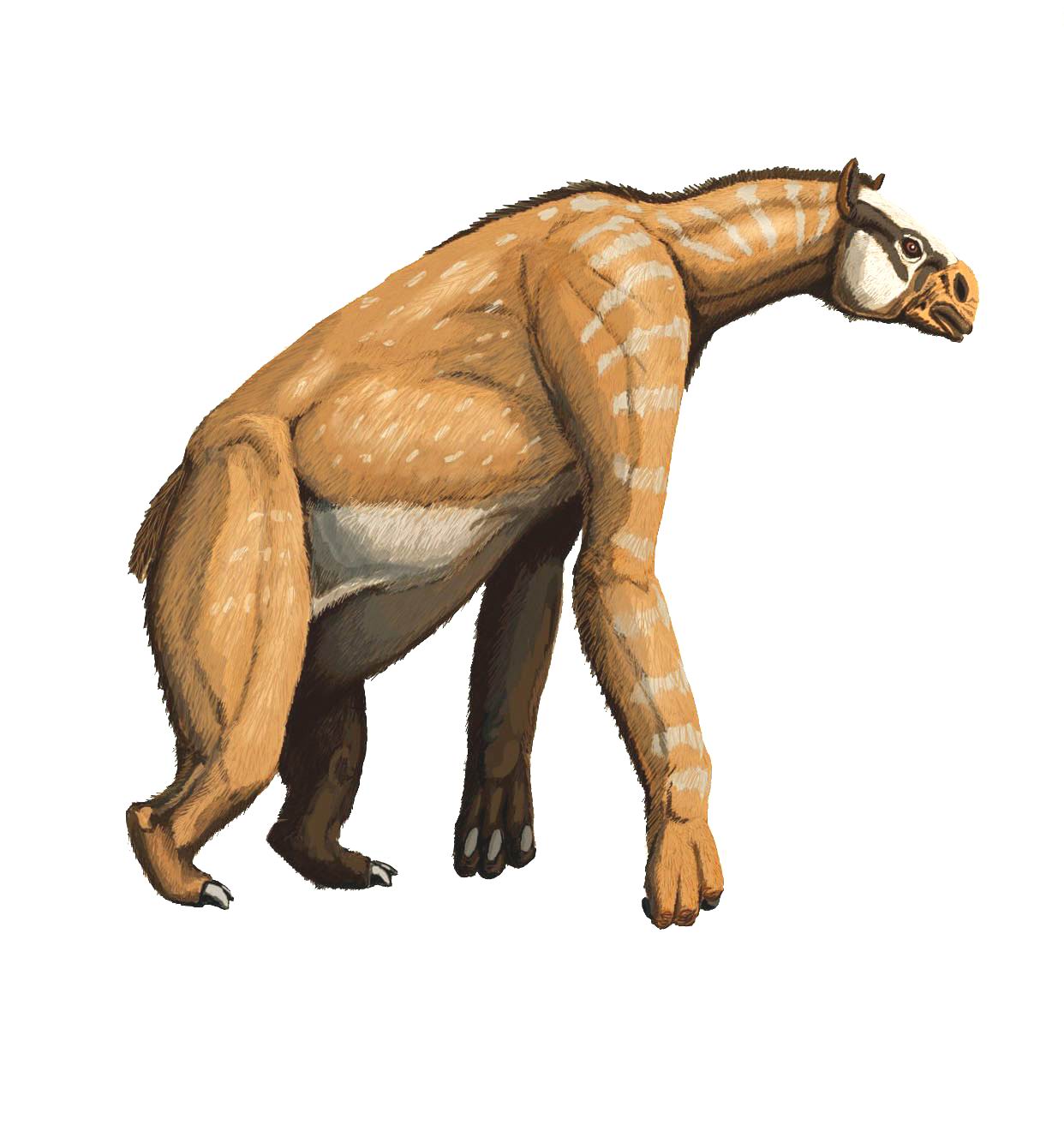
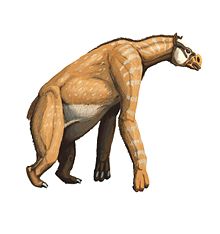 The chalicotheres vanished in the early Pleistocene
The chalicotheres vanished in the early PleistoceneLarge animals,which disappeared in Africa or Asia during the Early and Middle and Late Pleistocene:
- Giraffes, including Sivatherium.
- A species of wolf (Xenocyon lycaonoides)
- A few species of warthog such as Metridiochoerus
- Chalicotheres
- Deinotherium, Anancus and Mammuthus subplanifrons, relatives of the elephant
- Hippopotamus gorgops (a giant hippopotamus)
- False saber-toothed cats like Dinofelis
- Saber-toothed cats like Megantereon and Homotherium
- Giant tapir Megatapirus
- Giant ape Gigantopithecus
- Giant hyena Pachycrocuta
Large animals, which disappeared in parts of Africa and Asia during the Late Pleistocene:
- Giant Long-horned Buffalo (Pelorovis)
- Giant hartebeest Megalotragus
- Elephas recki (a species of elephant)
- Loxodonta adaurora (a species of African elephant)
Australia and New Guinea
See also: Australian megafauna Diprotodon became extinct around 50,000 years ago.
Diprotodon became extinct around 50,000 years ago.The sudden spate of extinctions occurred earlier than in the Americas. Most evidence points to the period immediately after the first arrival of humans—thought to be a little under 50,000 years ago—but scientific argument continues as to the exact date range. The Australian extinctions included:
- Diprotodon (giant relatives of the wombats)
- Zygomaturus (a "marsupial rhino")
- Hulitherium (a large marsupial herbivore)
- Phascolonus (a giant wombat)
- Palorchestes azael (a marsupial "tapir")
- Macropus titan (a giant kangaroo)
- Procoptodon goliah (a hoof-toed giant short-faced kangaroo)
- Sthenurus (a giant kangaroo)
- Simosthenurus (a giant kangaroo)
- Protemnodon (a giant kangaroo)
- Propleopus oscillans (an omnivorous kangaroo)
- Wonambi (a five-to-six-metre-long Australian constrictor snake)
- Thylacoleo carnifex (a lioness-sized marsupial carnivore)
- Megalania prisca (a giant monitor lizard)
- Dromornithidae (The entire family of giant birds. Though around the size of emus, they are actually more related to ducks and other waterfowl.)
Some extinct megafauna, such as the bunyip-like Diprotodon, may be the sources of ancient cryptozoological legends.
Northern Eurasia
 The Woolly Mammoth became extinct around 12,000 years ago – except on Wrangel Island and St. Paul Island, which humans did not colonize until much later.
The Woolly Mammoth became extinct around 12,000 years ago – except on Wrangel Island and St. Paul Island, which humans did not colonize until much later. A Collared Peccary, surviving relative of the extinct Giant Peccary.
A Collared Peccary, surviving relative of the extinct Giant Peccary.(80,000–10,000 years ago)
- Woolly mammoth (Mammuthus primigenius)
- Woolly rhinoceros (Coelodonta antiquitatis)
- Irish Elk (Megaloceros giganteus)
- Scimitar cat (Homotherium sp.)
- Cave lion (Panthera leo spelaea)
- Cave bear (Ursus spelaeus)
- Cave hyena (Crocuta crocuta spelaea)
- Steppe Wisent (Bison priscus)
- Elephants (Palaeoloxodon)
- Merk´s rhinoceros (Stephanorhinus kirchbergensis)
- Neanderthals (Homo neanderthalensis), survived until about 24,000 years ago in the Iberian peninsula.[3]
North America
 The size of a short faced bear compared with a human.
The size of a short faced bear compared with a human.During the last 50,000 years, including the end of the last glacial period, approximately 33 genera of large mammals have become extinct in North America. Of these, 15 genera extinctions can be reliably attributed to a brief interval of 11,500 to 10,000 radiocarbon years before present, shortly following the arrival of the Clovis people in North America. Most other extinctions are poorly constrained in time, though some definitely occurred outside of this narrow interval.[4] In contrast, only about half a dozen small mammals disappeared during this time. Previous North American extinction pulses had occurred at the end of glaciations, but not with such an imbalance between large mammals and small ones. (Moreover, previous extinction pulses were not comparable to the Quaternary extinction event; they involved primarily species replacements within ecological niches, while the latter event resulted in many ecological niches being left unoccupied.) The megafaunal extinctions include twelve genera of edible herbivores (H), and five large, dangerous carnivores (C). North American extinctions included:
- American horses, (Equus, three to five species) (H)
- Yukon wild ass (Equus lambei) (H)
- Western camel (Camelops hesternus) (H)
- North American llamas (Hemiauchenia)(H)
- Deer (Cervidae, two genera) (H)
- Pronghorn (Antilocapridae, two genera, one survived) (H)
- Stag-moose (Cervalces scotti) (H)
- Shrub-ox and Harlan's muskox (the Arctic Muskox survived) (H)
- Ancient Bison (Bison antiquus) (H)
- Long-horned bison / Giant bison (Bison latifrons) (H)
- Giant Beaver (Castoroides, two species) (H)
- Saiga (Saiga tatarica) (H)
- Teratorns (C)
- California tapirs (Tapirus californicus and Tapirus merriami) (H)
- Pinckney's Capybara (Neochoerus pinckneyi) (H)
- Eremotherium and Nothrotheriops, megatheriid ground sloths (H)
- Megalonyx, a megalonychid ground sloth (H)
- Paramylodon, a mylodontid ground sloth (H)
- Glyptodonts (H)
- Pampatheres (H)
- Short-faced Bear (Arctodus simus, larger than the present Grizzly Bear) (C)
- Florida Cave Bear (Tremarctos floridanus) (C)
- Saber-toothed cat (Smilodon fatalis) (C)
- Scimitar cat (Homotherium serum) (C)
- Giant polar bear (Ursus maritimus tyrannus, it is possible that this giant relative of the modern polar bear lived in North America) (C)
- American lion (Panthera leo atrox, larger than the current African Lion but probably a fairly recent immigrant through Beringia) (C)
- American cheetah (Miracinonyx, not a true cheetah, possibly ancestral) (C)
- Dire wolf (Canis dirus) (C)
- Mammoth (Mammuthus), several species (H)
- American mastodon (Mammut americanum) (H)
- Flat-headed (Platygonus) & Long-nosed (Mylohyus) peccaries (H)
- California (Tapirus californicus) & Vero (Tapirus veroensis) tapirs (H)
The survivors are in some ways as significant as the losses: bison, moose (a late Pleistocene immigrant through Beringia), elk, caribou, deer, pronghorn, muskox, bighorn sheep, and mountain goat. All save the pronghorns descended from Asian ancestors that had evolved with human predators.[5] Pronghorns are the second fastest land mammal (after the cheetah), which may have helped them elude hunters. More difficult to explain in the context of overkill is the survival of bison, since these animals first appeared in North America less than 240,000 years ago [6][7][8] and so were geographically removed from human predators for a sizeable period of time. Because ancient bison evolved into living bison,[9][10] there was no continent-wide extinction of bison at the end of the Pleistocene (although the genus was regionally extirpated in many areas). The survival of bison into the Holocene and recent times is therefore inconsistent with the overkill scenario. By the end of the Pleistocene, when humans first entered North America, these large animals had been geographically separated from human hunters for more than 200,000 years. Given this enormous span of geologic time, bison would almost certainly have been very nearly as naive as native North American large mammals.
The culture that has been connected with the wave of extinctions in North America is the paleo-Indian culture associated with the Clovis people (q.v.), who were thought to use spear throwers to kill large animals. The chief criticism of the "prehistoric overkill hypothesis" has been that the human population at the time was too small and/or not sufficiently widespread geographically to have been capable of such ecologically significant impacts. This criticism does not mean that climate change scenarios explaining the extinction are automatically to be preferred by default, however, any more than weaknesses in climate change arguments can be taken as supporting overkill. Some form of a combination of both factors could be plausible, although overkill would have the lion's share for the blame. It is easier to believe overkill would be a lot easier to achieve large-scale extinction with an already dying population due to climate change.[citation needed]
Lack of tameable megafauna was perhaps one of the reasons why Amerindian civilizations evolved differently than Old World ones.[11] Critics have disputed this by arguing that llamas, alpacas, and bison were domesticated.[12]
South America
 An illustration of Megatherium.
An illustration of Megatherium.At the Pleistocene-Holocene transition South America, which had remained largely unglaciated except for increased mountain glaciation in the Andes, saw an extinction wave, which carried off many large species. Unfortunately of those were the last of the Notoungulata (Toxodon, Macrauchenia) a group which have ruled South America for 57 million years and may have been common throughout the world before the rise of the modern ungulates. South America being an island continent it still had the primitive forms that have been replaced elsewhere, though most of them went extinct during the Great American Interchange. Of the megafauna survivors of the interchange, none survived the Quaternary one so only their smaller relatives remain (Anteaters, tree sloths, armadillos; New World Marsupials: Opposums, Shrew opossums, and Monito del Monte (actually more related to Australian marsupials, yet Australian marsupials evolved from a South American ancestor that may have been similar to Monito del Monte)). Of those that survived the interchange but not the Quaternary were the Ground sloths, Glyptodonts, Pampatheres, and the previously mentioned Notoungulatas; and of these all but the Notoungulatas migrated to North America, only to go extinct there too. Today the largest land mammals remaining in South America are the wild Lamini camels: Guanacos and Vicuñas and their domestic counterparts Llamas and Alpacas. Other notable surviving megafauna or large fauna are tapirs, rheas, Jaguars, Boa Constrictors, Anacondas, Caymans, and giant rodents like Capybaras.
- Smilodon fatalis and populator
- Arctodus
- Ground sloths of three families
- Glyptodonts
- Pampatheres
- Hippidion
- Horse (Equus)
- Toxodon
- Macrauchenia
- Cuvieronius
- Stegomastodon
Later extinctions
Mediterranean Islands
- Pygmy hippos of Cyprus (Phanourios minutus), Crete (Hippopotamus creutzburgi), Malta (H. melitensis) and Sicily (H. pentlandi)
- the Balearic Islands cave goat (Myotragus balearicus) of Majorca and Minorca
- Dwarf elephants of Cyprus (Elephas cypriotes), Sicily, Malta (E. falconeri) and many other islands
- Giant swan (Cygnus falconeri) of Malta
- Giant dormice: Minorcan Giant Dormouse, Majorcan Giant Dormouse
New Zealand
c. AD 1500, several species became extinct after Polynesian settlers arrived, including:
- Moa, giant flightless ratite birds, ten species
- Haast's Eagle (Harpagornis moorei)
- Adzebills, giant flightless predatory birds, two species
- Finsch's Duck, a large, flightless duck
Pacific, including Hawaii
Recent research, based on archaeological and paleontological digs on 70 different islands, has shown that numerous species went extinct as people moved across the Pacific, starting 30,000 years ago in the Bismarck Archipelago and Solomon Islands (Steadman & Martin 2003). It is currently estimated that among the bird species of the Pacific some 2000 species have gone extinct since the arrival of humans (Steadman 1995). Among the extinctions were:
- Moa-nalos, giant grazing Hawaiian ducks
- Nēnē-nui, or Woodwalking Goose, a large species of goose that once inhabited island of Maui
- Sylviornis, a giant galliform bird on New Caledonia
- Mekosuchine crocodiles on New Caledonia, Fiji and Samoa
- Meiolaniid turtles on Lord Howe Island and New Caledonia
Madagascar
Starting with the arrival of humans c. 2000 years ago, nearly all of the island's megafauna became extinct, including:
- Eight or more species of Elephant birds, giant flightless ratites in the genera Aepyornis and Mullerornis.
- 17 species of subfossil lemur, including:
- Giant aye-aye (Daubentonia robusta)
- Sloth lemurs, including chimpanzee-sized Palaeopropithecus and gorilla-sized Archaeoindris
- Pachylemur, a larger, more robust relative of the ruffed lemurs
- Monkey lemurs (Archaeolemuridae, the most terrestrial of all the known lemurs, often compared to baboons)
- Megaladapis, an orangutan-sized arboreal lemur similar to a koala
- Giant tortoise
- Malagasy Hippopotamus, three species
- Plesiorycteropus (an aardvark-like mammal, two species)
- Giant Fossa (Cryptoprocta spelea, a cougar-sized relative of the Fossa)
- Malagasy Crowned Eagle (Stephanoaetus mahery, a giant bird of prey)
Indian Ocean Islands
Starting c. 500 years ago, a number of species became extinct upon human settlement of the islands, including:
- Several species of giant tortoise on the Seychelles and Mascarene Islands
- Many species of birds on the Mascarene Islands, including the Dodo, the Rodrigues Solitaire, and the unrelated Réunion Solitaire.
Caribbean
Hunting hypothesis
The hunting hypothesis suggests that humans hunted megaherbivores to extinction. As a result, carnivores and scavengers that depended upon those animals became extinct from lack of prey.[13][14][15] Therefore this hypothesis holds Pleistocene humans responsible for the megafaunal extinction. One variant, often referred to as blitzkrieg, portrays humans as hunting the megafauna to extinction within a relatively short period of time. Some of the direct evidence for this includes: fossils of some megafauna found in conjunction with human remains, embedded arrows and tool cut marks found in megafaunal bones, and European cave paintings that depict such hunting. Biogeographical evidence is also suggestive; the areas of the world where humans evolved currently have more of their Pleistocene megafaunal diversity (the elephants and rhinos of Asia and Africa) compared to other areas such as Australia, the Americas, Madagascar and New Zealand, areas where early humans were non-existent. In addition, where animals have not been hunted for several years they become naive. Based on this evidence, a picture arises of the megafauna of Asia and Africa evolving with humans, learning to be wary of them, and in other parts of the world the wildlife appearing ecologically naive and easier to hunt.[citation needed] This is particularly true of island fauna, which display a dangerous lack of fear of humans. Of course, it is impossible to demonstrate that continental mammals were possessed of a similar naïveté.
Circumstantially, the close correlation in time between the appearance of humans in an area and extinction there provides weight for this scenario. This is perhaps the strongest evidence, as it is almost impossible that it could be coincidental when science has so many data points. For example, the woolly mammoth survived on islands despite changing climatic conditions for thousands of years after the end of the last glaciation, but they died out when humans arrived around 1700 BC. The megafaunal extinctions covered a vast period of time and highly variable climatic situations. The earliest extinctions in Australia were complete approximately 50,000 BP, well before the last glacial maximum and before rises in temperature. The most recent extinction in New Zealand was complete no earlier than 500 BP and during a period of cooling. In between these extremes megafaunal extinctions have occurred progressively in such places as North America, South America and Madagascar with no climatic commonality. The only common factor that can be ascertained is the arrival of humans.[16][17] Even within regions, this phenomenon appears to be true. The mammal extinction wave in Australia about 50,000 years ago coincides not with known climatic changes, but with the arrival of humans. In addition, large mammal species like the giant kangaroo Protemnodon appear to have survived in Tasmania longer than on the Australian mainland. Tasmania was colonised by humans a few thousand years after mainland Australia, which argues for the hunting hypothesis.[18][19]
 Extinctions occur in relation to human artifacts regardless of climate change in Australia, but appear linked to climate change in North America[citation needed]
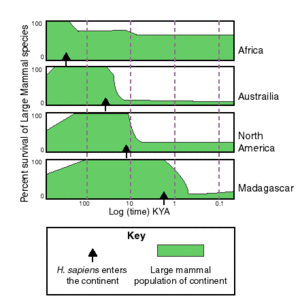
Extinctions occur in relation to human artifacts regardless of climate change in Australia, but appear linked to climate change in North America[citation needed]World wide extinctions seem to follow the migration of humans and to be most severe where humans arrived most recently and least severe where humans were originally—Africa (see figure at left). This suggests that in Africa, where humans evolved, prey animals and human hunting ability evolved together, so the animals evolved avoidance techniques. As humans migrated throughout the world and became more and more proficient at hunting, they encountered animals that had evolved without the presence of humans. Lacking the fear of humans that African animals had developed, animals outside of Africa were easy prey for human hunting techniques. It also suggests that this is independent of climate change (see figure at left).
Extinction through human hunting has been supported by archaeological finds of mammoths with projectile points embedded in their skeletons, by observations of modern naïve animals allowing hunters to approach easily[20][21][22] and by computer models by Mosimann and Martin,[23] and Whittington and Dyke,[24] and most recently by Alroy.[25]
Overkill hypothesis
 The timing of extinctions follows the "March of Man"
The timing of extinctions follows the "March of Man"The overkill hypothesis, a variant of the hunting hypothesis, was proposed 40 years ago by Paul S. Martin, Professor of Geosciences Emeritus at the Desert Laboratory of the University of Arizona. It sparked debate which continues today. It explains why the megafaunal extinctions occurred within a relatively short period of time. The most convincing evidence of his theory is that 80% of the North American large mammal species disappeared within 1000 years of the arrival of humans in the Americas.
Arguments regarding the hunting hypothesis
The major objections to the theory are as follows:
- In predator-prey models it is unlikely that predators could over-hunt their prey since predators need their prey as food to sustain life and reproduce.[26] This criticism has been rejected by many ecologists because humans have the widest dietary choice of any predator and are perfectly capable of switching to alternative prey or even plant foods when any prey species becomes rare. Humans have indisputably hunted numerous species to extinction, which renders any argument that human predators can never hunt prey to extinction immediately invalid.
- There is no archeological evidence that megafauna other than mammoths, mastodons, gomphotheres and bison were hunted. (Meltzer) Overkill proponents, however, say this is due to chance and the low probability of animals with low populations to be preserved. (Martin) Additionally, biochemical analyses have shown that Clovis tools were used in butchering horses and camels.[27]
- A small number of animals that were hunted, such as a single species of bison, did not go extinct. This cannot be explained by proposing that surviving bison in North America were recent Eurasian immigrants that were familiar with human hunting practices, since Bison first appeared in North America approximately 240,000 years ago [6][7][8] and then evolved into living bison.[9][28] Bison at the end of the Pleistocene were thus likely to have been almost as naive as their native North American megafaunal companions.
- The dwarfing of animals is not explained by overkill. Numerous authors[who?], however, have pointed out that dwarfing of animals is perfectly well explained by humans selectively harvesting the largest animals, and have provided proof that even within the 20th century numerous animal populations have reduced in average size due to human hunting.
- Eurasian Pleistocene megafauna went extinct in roughly same time period despite having a much longer time to adapt to hunting pressure by humans. However, the extinction of the Eurasian megafauna can be viewed as a result of a different process than that of the American megafauna. This makes the theory less parsimonious since another mechanism is required. The latter case occurred after the sudden appearance of modern human hunters on a land mass they had never previously inhabited, while the former case was the culmination of the gradual northward movement of human hunters over thousands of years as their technology for enduring extreme cold and bringing down big game improved. Thus, while the hunting hypothesis does not necessarily predict the rough simultaneity of the north Eurasian and American megafaunal extinctions, this simultaneity cannot be regarded as evidence against it.
- Eugene S. Hunn, President of the Society of Ethnobiology, offers a dissenting view. He points out that the birthrate in hunter-gatherer societies is generally too low, that too much effort is involved in the bringing down of a large animal by a hunting party, and that in order for hunter-gatherers to have brought about the extinction of megafauna simply by hunting them to death, an extraordinary amount of meat would have had to have been wasted.[29] It is possible that those who advocate the overkill hypothesis simply have not considered the differences in outlook between typical forager (hunter-gatherer) cultures and the present-day industrial cultures which exist in modernized human societies; waste may be tolerated and even encouraged in the latter, but is not so much in the former. It may be noted that in relatively recent human history, for instance, the Lakota of North America were known to take only as much bison as they could use, and they used virtually the whole animal—this despite having access to herds numbering in the millions.[30] Conversely, "buffalo jumps"[31] featured indiscriminate killing of a herd. However, Hunn's comments are in reference to a hunter-prey equilibrium state reached after thousands of years of coexistence, and are not relevant to hunters newly arrived on a virgin land mass full of easily taken big game. The well-established practice of industrial-scale moa butchering by the early Maori, involving enormous wastage of less choice portions of the meat, indicates that these arguments are incorrect.[20]
- The hypothesis that the Clovis culture represented the first humans to arrive in the New World has been disputed recently. (See Models of migration to the New World.)
Climate change hypothesis
At the end of the 19th and beginning of the 20th centuries, when scientists first realized that there had been glacial and interglacial ages, and that they were somehow associated with the prevalence or disappearance of certain animals, they surmised that the termination of the Pleistocene ice age might be an explanation for the extinctions.
Critics object that since there were multiple glacial advances and withdrawals in the evolutionary history of many of the megafauna, it is rather implausible that only after the last glacial would there be such extinctions. However, this criticism is rejected by a recent study indicating that terminal Pleistocene megafaunal community composition may have differed markedly from faunas present during earlier interglacials, particularly with respect to the great abundance and geographic extent of Pleistocene Bison at the end of the epoch.[32] This suggests that the survival of megafaunal populations during earlier interglacials is essentially irrelevant to the terminal Pleistocene extinction event, because bison were not present in similar abundance during any of the earlier interglacials.
Some evidence weighs against climate change as a valid hypothesis as applied to Australia. It has been shown that the prevailing climate at the time of extinction (40,000–50,000 BP) was similar to that of today, and that the extinct animals were strongly adapted to an arid climate. The evidence indicates that all of the extinctions took place in the same short time period, which was the time when humans entered the landscape. The main mechanism for extinction was likely fire (started by humans) in a then much less fire-adapted landscape. Isotopic evidence shows sudden changes in the diet of surviving species, which could correspond to the stress they experienced before extinction.[33][34][35]
Some evidence obtained from analysis of the tusks of mastodons from the American Great Lakes region appears inconsistent with the climate change hypothesis. Over a span of several thousand years prior to their extinction in the area, the mastodons show a trend of declining age at maturation. This is the opposite of what one would expect if they were experiencing stresses from deteriorating environmental conditions, but is consistent with a reduction in intraspecific competition that would result from a population being reduced by human hunting.[36]
Increased temperature
The most obvious change associated with the termination of an ice age is the increase in temperature. Between 15,000 BP and 10,000 BP, a 6°C increase in global mean annual temperatures occurred. This was generally thought to be the cause of the extinctions.
According to this hypothesis, a temperature increase sufficient to melt the Wisconsin ice sheet could have placed enough thermal stress on cold-adapted mammals to cause them to die. Their heavy fur, which helps conserve body heat in the glacial cold, might have prevented the dumping of excess heat, causing the mammals to die of heat exhaustion. Large mammals, with their reduced surface area-to-volume ratio, would have fared worse than small mammals.
Arguments against the temperature hypothesis
More recent research has demonstrated that the annual mean temperature of the current interglacial that we have seen for the last 10,000 years is no higher than that of previous interglacials, yet some of the same large mammals survived similar temperature increases. Therefore warmer temperatures alone may not be a sufficient explanation.[37][38][39][40][41][42] However, a recent study has demonstrated that Pleistocene bison were more abundant and more widely distributed in North America at the end of the Pleistocene than at any previous time during the epoch, and in many continental regions was among the most common large herbivores.[32] The documented increase in abundance of bison in midcontinent North America preceded the extinction event, and so cannot have been a result of the extinction.[32] Although it is premature to suggest that the increasing abundance of bison may have driven the extinction, these data nevertheless demonstrate that terminal Pleistocene megafaunal community composition was significantly different during the end-Pleistocene warming phase than that from any earlier warming periods.
In addition, numerous species such as mammoths on Wrangel Island[43] and St. Paul Island survived in human-free refugia despite changes in climate. This would not be expected if climate change were responsible. Under normal ecological assumptions island populations should be more vulnerable to extinction due to climate change because of small populations and an inability to migrate to more favorable climes.
Increased continentality affects vegetation in time or space
Other scientists have proposed that increasingly extreme weather—hotter summers and colder winters—referred to as "continentality", or related changes in rainfall caused the extinctions. The various hypotheses are outlined below.
Vegetation changes: geographic
It has been shown that vegetation changed from mixed woodland-parkland to separate prairie and woodland.[39][40][42] This may have affected the kinds of food available. If so, herbivores might not have found the plants with which they had evolved and thus would have fallen prey to the anti-herbivory toxins in the plants that remained available. Shorter growing seasons may have caused the extinction of large herbivores and the dwarfing of many others. In this case, as observed, bison and other large ruminants would have fared better than horses, elephants and other monogastrics, because ruminants are able to extract more nutrition from limited quantities of high-fiber food and better able to deal with anti-herbivory toxins.[44][45][46] So, in general, when vegetation becomes more specialized, herbivores with less diet flexibility may be less able to find the mix of vegetation they need to sustain life and reproduce, within a given area.
Rainfall changes: time
Increased continentality resulted in reduced and less predictable rainfall limiting the availability of plants necessary for energy and nutrition.[47][48][49] Axelrod[50] and Slaughter[51] have suggested that this change in rainfall restricted the amount of time favorable for reproduction. This could disproportionately harm large animals, since they have longer, more inflexible mating periods, and so may have produced young at unfavorable seasons (i.e., when sufficient food, water, or shelter was unavailable because of shifts in the growing season). In contrast, small mammals, with their shorter life cycles, shorter reproductive cycles, and shorter gestation periods, could have adjusted to the increased unpredictability of the climate, both as individuals and as species which allowed them to synchronize their reproductive efforts with conditions favorable for offspring survival. If so, smaller mammals would have lost fewer offspring and would have been better able to repeat the reproductive effort when circumstances once more favored offspring survival.[52]
Arguments against the continentality hypotheses
Critics have identified a number of problems with the continentality hypotheses.
- Megaherbivores have prospered at other times of continental climate. For example, megaherbivores thrived in Pleistocene Siberia, which had and has a more continental climate than Pleistocene or modern (post-Pleistocene, interglacial) North America.[53][54][55]
- The animals that went extinct actually should have prospered during the shift from mixed woodland-parkland to prairie, because their primary food source, grass, was increasing rather than decreasing.[56][55][57] Although the vegetation did become more spatially specialized, the amount of prairie and grass available increased, which would have been good for horses and for mammoths, and yet they went extinct. This criticism ignores the increased abundance and broad geographic extent of Pleistocene Bison at the end of the Pleistocene, which would have increased competition for these resources in a manner not seen in any earlier interglacials.[32]
- Although horses went extinct in the New World, they were successfully reintroduced by the Spanish in the 16th century—into a modern post-Pleistocene, interglacial climate. Today there are feral horses still living in those same environments. They find a sufficient mix of food to avoid toxins, they extract enough nutrition from forage to reproduce effectively and the timing of their gestation is not an issue. Of course, this criticism ignores the obvious fact that present-day horses are not competing for resources with ground sloths, mammoths, mastodons, camels, llamas, and bison. Similarly, mammoths survived the Pleistocene Holocene transition on isolated, uninhabited islands in the Mediterranean Sea[58] and on Wrangel Island in the Siberian Arctic[59] until 4,000 to 7,000 years ago.
- Large mammals should have been able to migrate, permanently or seasonally, if they found the temperature too extreme, the breeding season too short, or the rainfall too sparse or unpredictable.[60] Seasons vary geographically. By migrating away from the equator, herbivores could have found areas with growing seasons more favorable for finding food and breeding successfully. Modern-day African elephants migrate during periods of drought to places where there is apt to be water.[61]
- Large animals store more fat in their bodies than do medium-sized animals[62] and this should have allowed them to compensate for extreme seasonal fluctuations in food availability.
The extinction of the megafauna could have caused the disappearance of the mammoth steppe. Alaska now has low nutrient soil unable to support bison, mammoths, and horses. R. Dale Guthrie has claimed this as a cause of the extinction of the megafauna there; however, he may be interpreting it backwards. The loss of large herbivores to break up the permafrost allows the cold soils that are unable to support large herbivores today. Today, in the arctic, where trucks have broken the permafrost grasses and diverse flora and fauna can be supported.[63][64] In addition, Chapin (Chapin 1980) showed that simply adding fertilizer to the soil in Alaska could make grasses grow again like they did in the era of the mammoth steppe. Possibly, the extinction of the megafauna and the corresponding loss of dung is what led to low nutrient levels in modern day soil and therefore is why the landscape can no longer support megafauna.
Arguments against both climate change and overkill
Detractors have asserted that neither the overkill nor climate change hypotheses can explain several observations.[citation needed] Among these are the observation that browsers, mixed feeders and non-ruminant grazer species suffered most, while ruminant grazers generally survive[citation needed] However a broader variation of the overkill hypothesis may predict this, because changes in vegetation wrought by anthropogenic fire preferentially selects against browse species.[citation needed] Because of perceived shortcomings of the overkill or climate change hypotheses alone, some scientists support a combination of climate change and overkill.[citation needed]
Hyperdisease hypothesis
Theory
The Hyperdisease Hypothesis attributes the extinction of large mammals during the late Pleistocene to indirect effects of the newly arrived aboriginal humans.[65][66][67] The Hyperdisease Hypothesis proposes that humans or animals traveling with them (e.g., chickens or domestic dogs) introduced one or more highly virulent diseases into vulnerable populations of native mammals, eventually causing extinctions. The extinction was biased toward larger-sized species because smaller species have greater resilience because of their life history traits (e.g., shorter gestation time, greater population sizes, etc.). Humans are thought to be the cause because other earlier immigrations of mammals into North America from Eurasia did not cause extinctions.[65]
Diseases imported by people have been responsible for extinctions in the recent past; for example, bringing avian malaria to Hawaii has had a major impact on the isolated birds of the island.
If a disease was indeed responsible for the end-Pleistocene extinctions, then there are several criteria it must satisfy (see Table 7.3 in MacPhee & Marx 1997). First, the pathogen must have a stable carrier state in a reservoir species. That is, it must be able to sustain itself in the environment when there are no susceptible hosts available to infect. Second, the pathogen must have a high infection rate, such that it is able to infect virtually all individuals of all ages and sexes encountered. Third, it must be extremely lethal, with a mortality rate of c. 50–75%. Finally, it must have the ability to infect multiple host species without posing a serious threat to humans. Humans may be infected, but the disease must not be highly lethal or able to cause an epidemic.
One suggestion is that pathogens were transmitted by the expanding humans via the domesticated dogs they brought with them.[68] Unfortunately for such a theory it can not account for several major extinction events, notably Australia and North America. Dogs did not arrive in Australia until approximately 35,000 years after the first humans arrived and approximately 30,000 years after the megafaunal extinction was complete and as such can not be implicated. In contrast numerous species including wolves, mammoths, camelids and horses had emigrated continually between Asia and North America over the past 100,000 years. For the disease hypothesis to be applicable in the case of the Americas it would require that the population remain immunologically naive despite this constant transmission of genetic and pathogenic material.
Arguments against the hyperdisease hypothesis
- No evidence of disease has been found.
- Generally speaking, disease has to be very virulent to kill off all the individuals in a genus or species. Even such a virulent disease as West Nile Virus is unlikely to have caused extinction.[69]
- The disease would need to be implausibly selective while being simultaneously implausibly broad. Such a disease needs to be capable of killing of wolves such as Canis dirus or goats such as Oreamnos harringtoni while leaving other very similar species (Canis lupus and Oreamnos americanus, respectively) unaffected. It would need to be capable of killing off flightless birds while leaving closely related flighted species unaffected. Yet while remaining sufficiently selective to afflict only individual species within genera it must be capable of fatally infecting across such clades as birds, marsupials, placentals, testudines, and crocodilians. No disease with such a broad scope of fatal infectivity is known, much less one that remains simultaneously incapable of infecting numerous closely related species within those disparate clades.
Second-order predation
 Combination Hypotheses: Climate Change, Overkill + Climate Change, Second-Order Predation + Climate Change
Combination Hypotheses: Climate Change, Overkill + Climate Change, Second-Order Predation + Climate Change Overkill Hypothesis and Second-Order Predation
Overkill Hypothesis and Second-Order PredationScenario
The Second-Order Predation Hypothesis says that as humans entered the New World they continued their policy of killing predators, which had been successful in the Old World but because they were more efficient and because the fauna, both herbivores and carnivores, were more naive they killed off enough carnivores to upset the ecological balance of the continent, causing overpopulation, environmental exhaustion, and environmental collapse. The hypothesis accounts for changes in animal, plant, and, human populations.
The scenario is as follows:
- After the arrival of H. sapiens in the New World, existing predators must share the prey populations with this new predator. Because of this competition, populations of original, or first-order, predators cannot find enough food they are in direct competition with humans.
- Second-order predation begins as humans begin to kill predators.
- Prey populations are no longer well controlled by predation. Killing of nonhuman predators by H. sapiens reduces their numbers to a point where these predators no longer regulate the size of the prey populations.
- Lack of regulation by first-order predators triggers boom-and-bust cycles in prey populations. Prey populations expand and consequently overgraze and over-browse the land. Soon the environment is no longer able to support them. As a result, many herbivores starve. Species that rely on the slowest recruiting food become extinct, followed by species that cannot extract the maximum benefit from every bit of their food.
- Boom-bust cycles in herbivore populations change the nature of the vegetative environment, with consequent climatic impacts on relative humidity and continentality. Through overgrazing and overbrowsing, mixed parkland becomes grassland, and climatic continentality increases.
Support
This has been supported by a computer model, the Pleistocene Extinction Model (PEM), which, using the same assumptions and values for all variables (herbivore population, herbivore recruitment rates, food needed per human, herbivore hunting rates, etc.) other than those for hunting of predators. It compares the Overkill hypothesis (predator hunting = 0) with Second-Order Predation (predator hunting varied between 0.01 and 0.05 for different runs). The findings are that Second Order-Predation is more consistent with extinction than is Overkill[70][71] (results graph at left).
The PEM is the only test of multiple hypotheses and is the only model to specifically test combination hypotheses by artificially introducing sufficient climate change to cause extinction. When Overkill and Climate Change are combined they balance each other out. Climate Change reduces the number of plants, Overkill removes animals, therefore fewer plants are eaten. Second-Order Predation combined with Climate Change exacerbates the effect of Climate Change.[72] (results graph at right).
The second-order predation hypothesis is supported by the observation above that there was a massive increase in bison populations.(Scott)[73]
Second-order predation and other theories
- Climate Change: Second-Order Predation accounts for the changes in vegetation, which in turn may account for the increase in continentality. Since the extinction is due to destruction of habitat it accounts for the loss of animals not hunted by humans. Second-Order Predation accounts for the dwarfing of animals as well as extinctions since animals that could survive and reproduce on less food would be selectively favored.
- Hyperdisease: The reduction of carnivores could have been from distemper or other carnivore disease carried by domestic dogs.
- Overkill: The observation that extinctions follow the arrival of humans is supported by the Second-Order Predation hypothesis.
Arguments against the second-order predation hypothesis
- No evidence of humans hunting predators has been found in the New World (though it has been found in Siberia).[74]
- The model specifically assumes high extinction rates in grasslands, but most extinct species ranged across numerous vegetation zones. Historical population densities of ungulates were very high in the Great Plains; savanna environments support high ungulate diversity throughout Africa, and extinction intensity was equally severe in forested environments.
- It is unable to explain why large herbivore populations were not regulated by surviving carnivores such as grizzly bears, wolves, pumas, and jaguars whose populations would have increased rapidly in response to the loss of competitors.
- It does not explain why almost all extinct carnivores were large herbivore specialists such as sabre toothed cats and short faced bears, but most hypocarnivores and generalized carnivores survived.
- There is no historical evidence of boom and bust cycles causing even local extinctions in regions where large mammal predators have been driven extinct by hunting. The recent hunting out of remaining predators throughout most of the United States has not caused massive vegetational change or dramatic boom and bust cycles in ungulates.
- It is not spatially explicit and does not track predator and prey species separately, whereas the multispecies overkill model does both.
- The multispecies model produces a mass extinction through indirect competition between herbivore species: small species with high reproductive rates subsidize predation on large species with low reproductive rates.[25] All prey species are lumped in the Pleistocene Extinction Model.
- Everything explained by the Pleistocene Extinction Model also is explained by the multispecies model, but with fewer assumptions, so the Pleistocene Extinction Model appears less parsimonious. However, the multispecies model does not explain shifts in vegetation, nor is it able to simulate alternative hypotheses. The multispecies model therefore necessitates additional assumptions and hence is less parsimonious.
Arguments against the second-order predation plus climate hypothesis
- It assumes decreases in vegetation due to climate change, but deglaciation doubled the habitable area of North America.
- Any vegetational changes that did occur failed to cause almost any extinctions of small vertebrates, and they are more narrowly distributed on average.
Comet hypothesis
First publicly presented at the Spring 2007 joint assembly of the American Geophysical Union in Acapulco, Mexico, the comet hypothesis suggests that the mass extinction was caused by a swarm of comets 12,900 years ago. Using photomicrograph analysis, research published in January 2009 has found evidence of nanodiamonds in the soil from six sites across North America including Arizona, Minnesota, Oklahoma, South Carolina and two Canadian sites. Similar research found nanodiamonds in the Greenland ice sheet.[75][76][77]
Arguments against the comet hypothesis
Debate around this hypothesis has included, among other things, the lack of an impact crater, relatively small increased level of iridium in the soil, and the highly improbable nature of such an event.
- There is a lack of evidence for a population decline among the Paleoindians at 12,900 ± 100 calBP as might be expected.[78][79]
- There is evidence that the megafaunal extinctions that occurred across northern Eurasia, North America and South America at the end of the Pleistocene were not synchronous as the bolide theory would predict. The extinctions in South America appear to have occurred at least 400 years after those in North America.[80][81][82]
- Additionally, some island megafaunal populations survived thousands of years longer than populations of the same or related species on nearby continents; examples include the survival of woolly mammoths on Wrangel Island until 3700 BP,[80][81] and the survival of ground sloths in the Antilles until 4700 cal BP.[80][81][82]
See also
References
- ^ Anderson, Paul K. (July 1995). "Competition, Predation, and the Evolution and Extinction of Steller’s Sea Cow, Hydrodamalis Gigas". Marine Mammal Science (Society for Marine Mammalogy) 11 (3): 391–4. doi:10.1111/j.1748-7692.1995.tb00294.x. http://www3.interscience.wiley.com/journal/119963340/abstract.
- ^ Julien Louys, Darren Curnoe, Haowen Tong. (2007). "Characteristics of Pleistocene megafauna extinctions in Southeast Asia". Palaeogeography, Palaeoclimatology, Palaeoecology 243: 152–173. doi:10.1016/j.palaeo.2006.07.011.
- ^ Rincon, Paul (13 September 2006). "Neanderthals' 'last rock refuge'". BBC News. http://news.bbc.co.uk/1/hi/5343266.stm?lsm. Retrieved 2009-10-11.
- ^ Anthony D. Barnosky, Paul L. Koch, Robert S. Feranec, Scott L. Wing, Alan B. Shabel (2004). "Assessing the Causes of Late Pleistocene Extinctions on the Continents". Science 306 (5693): 70–75. Bibcode 2004Sci...306...70B. doi:10.1126/science.1101476. PMID 15459379.
- ^ MacPhee, RDE (1999). Extinctions in Near Time: Causes, Contexts, and Consequences. Kluwer Academic Publishers. ISBN 0306460920.
- ^ a b Bell, C.J., et al. (2004). "The Blancan, Irvingtonian, and Rancholabrean mammal ages". In Woodburne, M.O.. Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology. New York: Columbia Univ. Press. pp. 232–314. ISBN 0231130406.
- ^ a b Scott, E., Cox, S.M. (2008). "Late Pleistocene distribution of Bison (Mammalia; Artiodactyla) in the Mojave Desert of Southern California and Nevada". In Wang, X., Barnes, L.G.. Geology and Vertebrate Paleontology of Western and Southern North America. Los Angeles: Natural History Museum of Los Angeles County. pp. 359–382.
- ^ a b Sanders, A.E., R.E. Weems, and L.B. Albright III (2009). "Formalization of the mid-Pleistocene "Ten Mile Hill beds" in South Carolina with evidence for placement of the Irvingtonian–Rancholabrean boundary". In Albright III, L.B.. Papers on Geology, Vertebrate Paleontology, and Biostratigraphy in Honor of Michael O. Woodburne. Flagstaff: Museum of Northern Arizona. pp. 369–375.
- ^ a b Shapiro, B., et al. (2004). "Rise and Fall of the Beringian Steppe Bison". Science 306 (5701): 1561–1565. Bibcode 2004Sci...306.1561S. doi:10.1126/science.1101074. PMID 15567864.
- ^ Wilson, M.C., L.V. Hills, and B. Shapiro (2008). "Late Pleistocene northward-dispersing Bison antiquus from the Bighill Creek Formation, Gallelli Gravel Pit, Alberta, Canada, and the fate of Bison occidentalis". Canadian Journal of Earth Sciences 45 (7): 827–859. Bibcode 2008CaJES..45..827W. doi:10.1139/E08-027.
- ^ Diamond, J. (1997). Guns, Germs, and Steel: The Fates of Human Societies. WW Norton. ISBN 978-0393061314.
- ^ Pielou, EC (1992). After the Ice Age: the return of life to glaciated North America. University of Chicago Press. ISBN 978-0226668123.
- ^ Martin P. S. (1963). The last 10,000 years: A fossil pollen record of the American Southwest. Tucson, AZ: Univ. Ariz. Press. ISBN 0-8165-1759-2.
- ^ Martin P. S. (1967). "Prehistoric overkill". In Martin, P.S., Wright, H.E.. Pleistocene extinctions: The search for a cause. New Haven: Yale Univ. Press. ISBN 0-300-00755-8.
- ^ Martin P. S. (1989). "Prehistoric overkill: A global model". In Martin, P.S., Klein, R.G.. Quaternary extinctions: A prehistoric revolution. Tucson, AZ: Univ. Arizona Press. pp. 354–404. ISBN 0-8165-1100-4.
- ^ Martin, P. S. (2005). Twilight of the Mammoths: Ice Age Extinctions and the Rewilding of America. University of California Press. ISBN 0520231414. http://books.google.com/?id=eThoCsL1hRAC.
- ^ Burney, D. A.; Flannery, T. F. (July 2005). "Fifty millennia of catastrophic extinctions after human contact" (PDF). Trends in Ecology & Evolution (Elsevier) 20 (7): 395–401. doi:10.1016/j.tree.2005.04.022. PMID 16701402. http://www.anthropology.hawaii.edu/Fieldschools/Kauai/Publications/Publication%204.pdf.
- ^ Diamond J (2008). "Palaeontology: The last giant kangaroo". Nature 454 (7206): 835–6. Bibcode 2008Natur.454..835D. doi:10.1038/454835a. PMID 18704074.
- ^ Turneya CSM, Flannery TF, Roberts RG, Reid C, Fifield LK, Higham TFG, Jacobs Z, Kemp N, Colhoun EA, Kalin RM, Ogle N (26 August 2008). "Late-surviving megafauna in Tasmania, Australia, implicate human involvement in their extinction". Proc. Natl. Acad. Sci. U.S.A. 105 (34): 12150–3. Bibcode 2008PNAS..10512150T. doi:10.1073/pnas.0801360105. PMC 2527880. PMID 18719103. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2527880.
- ^ a b Flannery, Tim (2002-10-16). The future eaters: an ecological history of the Australasian lands and people. New York: Grove/Atlantic, Inc.. ISBN 0802139434. OCLC 32745413. http://google.com/books?id=eIW5aktgo0IC&printsec=frontcover.
- ^ Diamond, J. (1984). "Historic extinctions: a Rosetta stone for understanding prehistoric extinctions". In Martin, P.S., Klein, R.G.. Quaternary extinctions: A prehistoric revolution. Tucson, AZ: Univ. Arizona Press. pp. 824–62. ISBN 0-8165-1100-4.
- ^ Diamond, J. (1997). Guns, germs, and steel; the fates of human societies. New York: Norton. ISBN 0-393-31755-2.
- ^ Mossiman, J. E., and Martin, P. S. (1975). "Simulating Overkill by Paleoindians". American Scientist 63: 304–13. Bibcode 1975AmSci..63..304M.
- ^ Whittington, S. L. & Dyke, B. (1984). "Simulating overkill: experiment with the Mossiman and Martin model". In Martin, P.S., Klein, R.G.. Quaternary extinctions: A prehistoric revolution. Tucson, AZ: Univ. Arizona Press. pp. 451–66. ISBN 0-8165-1100-4.
- ^ a b Alroy, J. (2001). "A multispecies overkill simulation of the end-Pleistocene megafaunal mass extinction". Science 292 (5523): 1893–6. Bibcode 2001Sci...292.1893A. doi:10.1126/science.1059342. PMID 11397940. http://www.nceas.ucsb.edu/~alroy/pdfs/2001-Science-292-1893.pdf.
- ^ May, R. M. (2001). Stability and complexity in model ecosystems. Princeton: Princeton Univ. Press. ISBN 0-691-08861-6.
- ^ Scott, J. (2009-02-26). "Camel-butchering in Boulder, 13,000 years ago". Colorado Arts and Sciences Magazine. University of Colorado at Boulder. http://artsandsciences.colorado.edu/magazine/2009/02/camel-butchering-in-boulder-13000-years-ago/. Retrieved 2009-05-01.
- ^ Wilsom, M.C., L.V. Hills, and B. Shapiro (2008). "Late Pleistocene northward-dispersing Bison antiquus from the Bighill Creek Formation, Gallelli Gravel Pit, Alberta, Canada, and the fate of Bison occidentalis". Canadian Journal of Earth Sciences 45 (7): 827–859. Bibcode 2008CaJES..45..827W. doi:10.1139/E08-027.
- ^ Nadasdy, Paul (2006). "Transcending the Debate over the Ecologically Noble Indian: Indigenous Peoples and Environmentalism". Ethnohistory 52 (2). http://ethnohistory.dukejournals.org/cgi/content/refs/52/2/291.
- ^ Svenson, Sally. "Bison." South Dakota Department of Game, Fish and Parks, 1995.
- ^ "Buffalo Jump." Wikipedia.
- ^ a b c d Scott, E. (2010). "Extinctions, scenarios, and assumptions: Changes in latest Pleistocene large herbivore abundance and distribution in western North America". Quat. Int. 217: 225. Bibcode 2010QuInt.217..225S. doi:10.1016/j.quaint.2009.11.003.
- ^ "Thylacoleo — The Beast of the Nullarbor". Willis, Paul; Bryce, Clay; Searle, Mike. Catalyst. Australian Broadcasting Commission. 17 August 2006. Transcript.
- ^ Prideaux GJ, Long JA, Ayliffe LK, et al. (January 2007). "An arid-adapted middle Pleistocene vertebrate fauna from south-central Australia". Nature 445 (7126): 422–5. Bibcode 2007Natur.445..422P. doi:10.1038/nature05471. PMID 17251978. http://www.nature.com/nature/journal/v445/n7126/abs/nature05471.html.
- ^ Roberts RG, Flannery TF, Ayliffe LK, et al. (June 2001). "New ages for the last Australian megafauna: continent-wide extinction about 46,000 years ago". Science 292 (5523): 1888–92. Bibcode 2001Sci...292.1888R. doi:10.1126/science.1060264. PMID 11397939. http://www.sciencemag.org/cgi/pmidlookup?view=long&pmid=11397939.
- ^ Fisher, Daniel C. (2009). "Paleobiology and Extinction of Proboscideans in the Great Lakes Region of North America". In Haynes, Gary. American Megafaunal Extinctions at the End of the Pleistocene. Springer. pp. 55–75. doi:10.1007/978-1-4020-8793-6_4. ISBN 978-1-4020-8792-9. http://www.springerlink.com/content/m39544m241500322/fulltext.pdf.
- ^ Andersen, S. T (1973). "The differential pollen productivity of trees and its significance for the interpretation of a pollen diagram from a forested region". In Birks, H.J.B., West, R.G.. Quaternary plant ecology: the 14thsymposium of the British Ecological society, University of Cambridge, 28–30 March 1972. Oxford: Blackwell Scientific. ISBN 0-632-09120-7.
- ^ Ashworth, C.A. (1980). "Environmental implications of a beetle assemblage from the Gervais formation (Early Wisconsinian?), Minnesota". Quat. Res. 13 (2): 200–12. Bibcode 1980QuRes..13..200A. doi:10.1016/0033-5894(80)90029-0.
- ^ a b Birks, H.H. (1973). "Modern macrofossil assemblages in lake sediments in Minnesota". In Birks, H.J.B., West, R.G.. Quaternary plant ecology: the 14thsymposium of the British Ecological Society, University of Cambridge, 28–30 March 1972. Oxford: Blackwell Scientific. ISBN 0-632-09120-7.
- ^ a b Birks, H.J.B., Birks, H.H. (1980). Quaternary paleoecology. Baltimore: Univ. Park Press. ISBN 1-930665-56-3.
- ^ Bradley, R. S. (1985). Quaternary Paleoclimatology: Methods of Paleoclimatic Reconstruction. Winchester, MA: Allen & Unwin. ISBN 0-04-551068-7.
- ^ a b Davis, M. B. (1976). "Pleistocene biogeography of temperate deciduous forests". Geoscience and man: Ecology of the Pleistocene. 13. Baton Rouge: School of Geoscience, Louisiana State Univ..
- ^ Vartanyan, S.L., Arslanov, K.A., Tertychnaya, T.V. & Chernov, S.B. (1995). "Radiocarbon dating evidence for mammoths on Wrangel Island, Arctic Ocean, until 2000 BC". Radiocarbon 37: 1–6. http://www.radiocarbon.org/Journal/v37n1/vartanyan.html.
- ^ Guthrie, R. D. (1988). Frozen Fauna of the Mammoth Steppe: The Story of Blue Babe. University Of Chicago Press. ISBN 0-226-31122-8.
- ^ Guthrie, R. D. (1989). "Mosaics, allochemics, and nutrients: an ecological theory of Late Pleistocene megafaunal extinctions". In Martin, P.S., Klein, R.G.. Quaternary extinctions: A prehistoric revolution. Tucson, AZ: Univ. Arizona Press. pp. 259–99. ISBN 0-8165-1100-4.
- ^ Hoppe, P.P. (1978). "Rumen fermentation in African ruminants". Proceedings of the 13th Annual Congress of Game Biologists. Atlanta.
- ^ Bryson, R.A., Baerreis, D.A., Wendland, W.M. (1970). "The character of late-glacial and post-glacial climatic changes". In Dort Jr., W., Jones, Jr., J.K.. Pleistocene and recent environments of the central Great Plains. Lawrence: Univ. Press Kan. ISBN 0-7006-0063-9. Univ. Kan. Spec. Pub. 3.
- ^ Graham, R.W., Lundelius, E.L. (1989). "Coevolutionary disequilibrium and Pleistocene extinctions". In Martin, P.S., Klein, R.G.. Quaternary extinctions: A prehistoric revolution. Tucson, AZ: Univ. Arizona Press. pp. 354–404. ISBN 0-8165-1100-4.
- ^ King, J.E., Saunders, J.J. (1989). "Environmental insularity and the extinction of the American mastodont". In Martin, P.S., Klein R.G.. Quaternary extinctions: A prehistoric revolution. Tucson, AZ: Univ. Arizona Press. pp. 354–404. ISBN 0-8165-1100-4.
- ^ Axelrod, D. I. (1967). "Quaternary extinctions of large mammals". University of California Publications in Geological Sciences 74: 1–42. ASIN B0006BX8LG.
- ^ Slaughter, B. H. (1967). "Animal ranges as a clue to late-Pleistocene extinction". In Martin, P.S., Wright H.E.. Pleistocene extinctions: The search for a cause. New Haven: Yale Univ. Press. ISBN 0-300-00755-8.
- ^ Kilti, R. A. (1988). "Seasonality, gestation time, and large mammal extinctions". In Martin, P.S., Klein R.G.. Quaternary extinctions: A prehistoric revolution. Tucson, AZ: Univ. Arizona Press. pp. 354–404. ISBN 0-8165-1100-4.
- ^ Flereov, C.C. (1967). "On the origin of the mammalian fauna of Canada". In Hopkins, D.M.. The Bering Land Bridge. Palo Alto: Stanford Univ. Press. pp. 271–80. ISBN 0-8047-0272-1.
- ^ Frenzel, B. (1968). "The Pleistocene vegetation of northern Eurasia". Science 161 (3842): 637–49. Bibcode 1968Sci...161..637F. doi:10.1126/science.161.3842.637. PMID 17801456.
- ^ a b McDonald, J. (1989). "The reordered North American selection regime and late Quaternary megafaunal extinctions". In Martin, P.S., Klein, R.G.. Quaternary extinctions: A prehistoric revolution. Tucson, AZ: Univ. Arizona Press. pp. 354–404. ISBN 0-8165-1100-4.
- ^ Birks, H.J.B., West, R.G. (1973). Quaternary plant ecology: the 14th symposium of the British Ecological society, University of Cambridge, 28–30 March 1972. Oxford: Blackwell Scientific. ISBN 0-632-09120-7.
- ^ McDonald, J. (1981). North American Bison: Their classification and evolution.. Berkeley: Univ. Calif. Press. ISBN 0-520-04002-3.
- ^ Burney, D. A. (1993). "Recent animal extinctions: recipes for disaster". American Scientist 81 (6): 530–41. Bibcode 1993AmSci..81..530B.
- ^ Vartanyan, S.L., Garutt, V. E. and Sher, A.V. (1993). "Holocene dwarf mammoths from Wangel Island in the Siberian Arctic". Nature 362 (6418): 337–40. Bibcode 1993Natur.362..337V. doi:10.1038/362337a0.
- ^ Pennycuick, C.J. (1979). "Energy costs of locomotion and the concept of "Foraging radius"". In Sinclair A.R.E., Norton-Griffiths M.. Serengetti: Dynamics of an Ecosystem. Chicago: Univ. Chicago Press. pp. 164–85. ISBN 0-226-76029-4.
- ^ Wing, L.D., Buss, I.O. (1970). "Elephants and Forests". Wildl. Mong. (19).
- ^ Owen-Smith, R.N. (1992). Megaherbivores: The influence of very large body size on ecology. Cambridge studies in ecology.. Cambridge: Cambridge Univ. Press. ISBN 0-521-42637-5.
- ^ Kershaw, G.P. (1984). "Tundra plant communities of the Mackenzie mountains, Northwest Territories; floristic characteristics of long term surface disturbances". In Olson, R., Hastings, R., Geddes, F.. Northern Ecology and Resource Management: Memorial Essays honoring Don Gill. Edmonton, Canada: Univ. Alberta Press. pp. 239–311. ISBN 0888640471.
- ^ Webber, P.J., Miller, P.C., Chapin, F.S. III, MacCown, B.H. (1980). "The vegetation: pattern and succession". In Brown, J., Miller, P.C., Tieszen, L.L., Bunnell, F.L.. An Arctic ecosystem: the coastal tundra at Barrow, Alaska. US/IBP Synthesis id=12. Stroudsburg, PA: Dowden Hutchinson & Ross. pp. 186–219.
- ^ a b MacFee, R.D.E. & Marx, P.A. (1997). "Humans, hyperdisease and first-contact extinctions". In Goodman, S., Patterson, B.D.. Natural Change and Human Impact in Madagascar. Washington D.C.: Smithsonian Press. pp. 169–217. ISBN 1-56098-683-2.
- ^ MacFee, R.D.E. & Marx, P.A. (1998). "Lightning Strikes Twice: Blitzkrieg, Hyperdisease, and Global Explanations of the Late Quaternary Catastrophic Extinctions". American Museum of Natural History. http://www.amnh.biz/science/biodiversity/extinction/Day1/bytes/MacPheePres.html.
- ^ MacPhee, Ross D.E.; Preston Marx (1997). "The 40,000-year Plague: Humans, Hyperdisease, and First-Contact Extinctions". Natural Change and Human Impact in Madagascar. Washington, D.C.: Smithsonian Institution Press. pp. 169–217.
- ^ Fiedel, S (2005). "Man's best friend: mammoth's worst enemy?". World Archaeology 37: 11–35.
- ^ Lyons, K, Smith, F.A., Wagner, P.J., White, E.P., and Brown, J.,H. (2004). "Was a 'hyperdisease' responsible for the late Pleistocene megafaunal extinction?". Ecology 7: 859–68. http://biology.unm.edu/fasmith/Web_Page_PDFs/Lyons_et_al_2004_WN.pdf.
- ^ Whitney-Smith, E. (2004). "Late Pleistocene extinctions through second-order predation". In Barton, C.M., Clark, G.A., Yesner, D.R.. Settlement of the American Continents: A Multidisciplinary Approach to Human Biogeography. Tucson, AZ: University of Arizona Press. ISBN 0-8165-2323-1.
- ^ Whitney-Smith, E. (2009). The Second-Order Predation Hypothesis of Pleistocene Extinctions: A System Dynamics Model. Saarbruken, Germany: VDM Verlag. ISBN 978-3639115796.
- ^ Whitney-Smith, E. (2006). Clovis and Extinctions – Overkill, Second Order Predation, Environmental Degradation in a Non-equilibrium Ecosystem "Clovis Age Continent". University of New Mexico Press.
- ^ Scott, E. (2010). "Extinctions, scenarios, and assumptions: Changes in latest Pleistocene large herbivore abundance and distribution in western North America". Quat. Int.
- ^ Soffer, O. (1985). The Upper Paleolithic of the Central Russian Plain. Orlando, Florida: Academic Press. ISBN 0-12-654270-8.
- ^ Study links mammoth extinction, comets, USA Today, Updated 1/2/2009 7:43 AM, retrieved 4 Apr 2009
- ^ The Extinction Debate, by Evan Hadingham, NOVA, date retrieved 4 Apr 2009
- ^ Last Extinction, NOVA, TV Program Description,Original PBS Broadcast Date: March 31, 2009, date retrieved 4 Apr 2009
- ^ Buchanan B, Collard M, Edinborough K (19 August 2008). "Paleoindian demography and the extraterrestrial impact hypothesis". Proc. Natl. Acad. Sci. U.S.A. 105 (33): 11651–4. Bibcode 2008PNAS..10511651B. doi:10.1073/pnas.0803762105. PMC 2575318. PMID 18697936. http://www.pnas.org/content/105/33/11651.abstract.
- ^ Haynes, Gary (2009). American megafaunal extinctions at the end of the Pleistocene. Springer. p. 125. ISBN 9781402087929. http://books.google.com/books?id=iq6qZXUkWo0C&pg=PA125.
- ^ a b c Haynes, Gary (2009). "Introduction to the Volume". In Haynes, Gary. American Megafaunal Extinctions at the End of the Pleistocene. Springer. pp. 1–20. doi:10.1007/978-1-4020-8793-6_1. ISBN 978-1-4020-8792-9. http://www.springerlink.com/content/w314m76738r91g35/?p=5af1eb7387d443a2b514b284c646efa7&pi=0.
- ^ a b c Fiedel, Stuart (2009). "Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction". In Haynes, Gary. American Megafaunal Extinctions at the End of the Pleistocene. Springer. pp. 21–37. doi:10.1007/978-1-4020-8793-6_2. ISBN 978-1-4020-8792-9. http://www.springerlink.com/content/l225628681672725/?p=5af1eb7387d443a2b514b284c646efa7&pi=1.
- ^ a b Vergano, Dan (2009-01-02). "Study links mammoth extinction, comets". USA Today.Com. Gannett Company. http://www.usatoday.com/tech/science/2009-01-01-mammothimpact_N.htm. Retrieved 2009-05-10.
External links
Hyperdisease hypothesis
- "American Museum of Natural History". http://www.amnh.biz/science/biodiversity/extinction/Day1/bytes/MacPheePres.html.
- J.H. Brown (University of New Mexico). "Was a hyperdisease responsible?". http://biology.unm.edu/JHBrown/Published/WasAHyperdiseaseResponsible.x.pdf.
Second-order predation
- Elin Whitney-Smith. "Quaternary.Net". http://quaternary.net.
Other links
- "Ice Age Bay Area". http://www.kqed.org/quest/television/ice-age-bay-area2.
- "The Extinct Late Pleistocene Mammals of North America". http://www.pbs.org/wgbh/nova/stoneage/mega-list.html.
- "End of the Big Beasts". http://www.pbs.org/wgbh/nova/beta/evolution/end-big-beasts.html.
- "Of mice, mastodons and men: human-mediated extinctions on four continents". http://biology.unm.edu/fasmith/Web_Page_PDFs/Lyons_et_al_2004_EER.pdf.
- "Return to the Ice Age: The La Brea Exploration Guide". http://www.tarpits.org/education/guide/index.html.
Quaternary Pleistocene Holocene Early | Middle | Late Preboreal | Boreal |
Atlantic | Subboreal | SubatlanticPreceded by Proterozoic Eon 542 Ma - Phanerozoic Eon - Present 542 Ma - Paleozoic Era - 251 Ma 251 Ma - Mesozoic Era - 65 Ma 65 Ma - Cenozoic Era - Present Cambrian Ordovician Silurian Devonian Carboniferous Permian Triassic Jurassic Cretaceous Paleogene Neogene Quaternary Categories:- Extinction events
- Pleistocene







Wikimedia Foundation. 2010.