- Factor H
-
Complement factor H 
PDB rendering based on 1haq.Available structures PDB 1FHC, 1HAQ, 1HCC, 1HFH, 1HFI, 1KOV, 2BZM, 2G7I, 2IC4, 2JGW, 2JGX, 2KMS, 2QFG, 2QFH, 2RLP, 2RLQ, 2UWN, 2V8E, 2W80, 2W81, 2WII, 3GAU, 3GAV, 3GAW, 3KXV, 3KZJ Identifiers Symbols CFH; AHUS1; AMBP1; ARMD4; ARMS1; CFHL3; FH; FHL1; HF; HF1; HF2; HUS; MGC88246 External IDs OMIM: 134370 MGI: 88385 HomoloGene: 20086 GeneCards: CFH Gene Gene Ontology Cellular component • extracellular region
• extracellular spaceBiological process • complement activation
• complement activation, alternative pathwaySources: Amigo / QuickGO RNA expression pattern 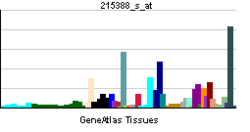

More reference expression data Orthologs Species Human Mouse Entrez 3075 12628 Ensembl ENSG00000000971 ENSMUSG00000026365 UniProt P08603 n/a RefSeq (mRNA) NM_000186.3 NM_009888.3 RefSeq (protein) NP_000177.2 NP_034018.2 Location (UCSC) Chr 1:
196.62 – 196.72 MbChr 1:
141.98 – 142.08 MbPubMed search [2] [3] Factor H is a member of the regulators of complement activation family and is a complement control protein. It is a large (155 kilodaltons), soluble glycoprotein that circulates in human plasma (at a concentration of 500–800 micrograms per milliliter). Its principal function is to regulate the Alternative Pathway of the complement system, ensuring that the complement system is directed towards pathogens and does not damage host tissue. Factor H regulates complement activation on self cells by possessing both cofactor activity for the Factor I mediated C3b cleavage, and decay accelerating activity against the alternative pathway C3 convertase, C3bBb. Factor H protects self cells from complement activation but not bacteria/viruses, in that it binds to glycosaminoglycans (GAGs) that are present on host cells but not pathogen cell surfaces.[1][2]
Contents
Structure
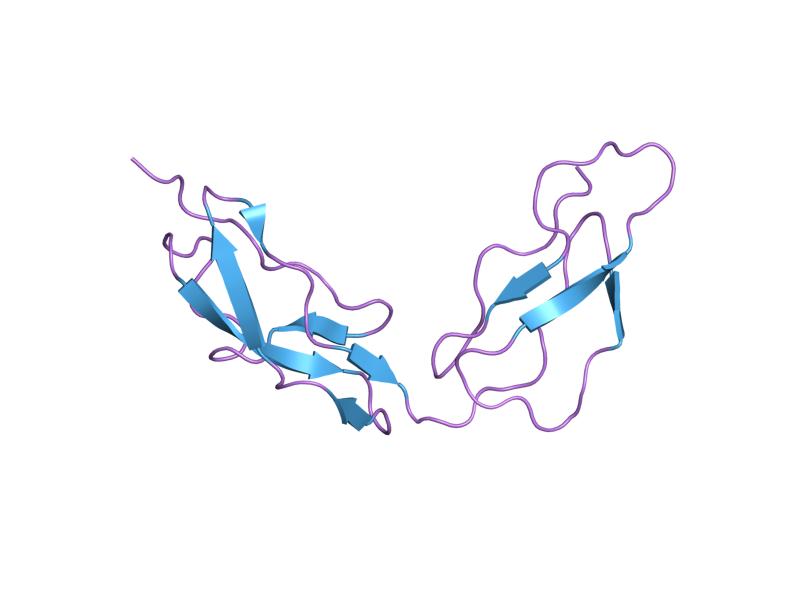
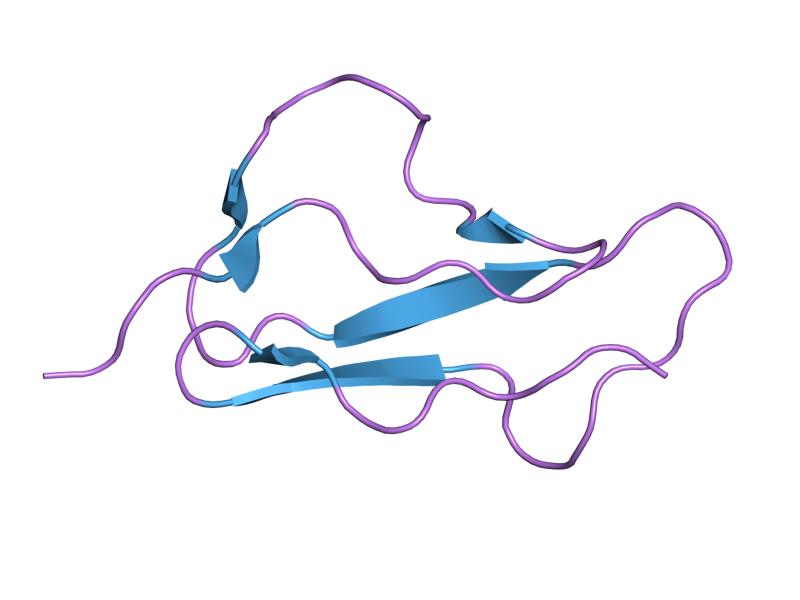
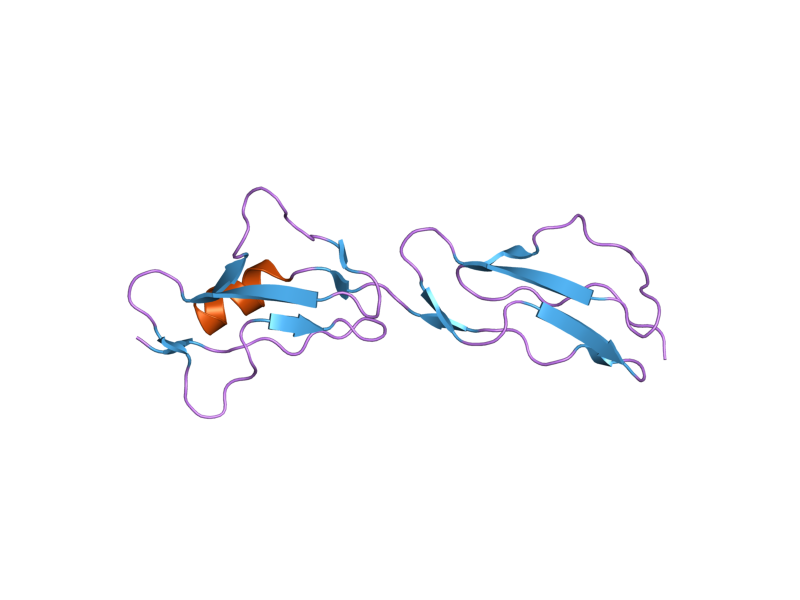
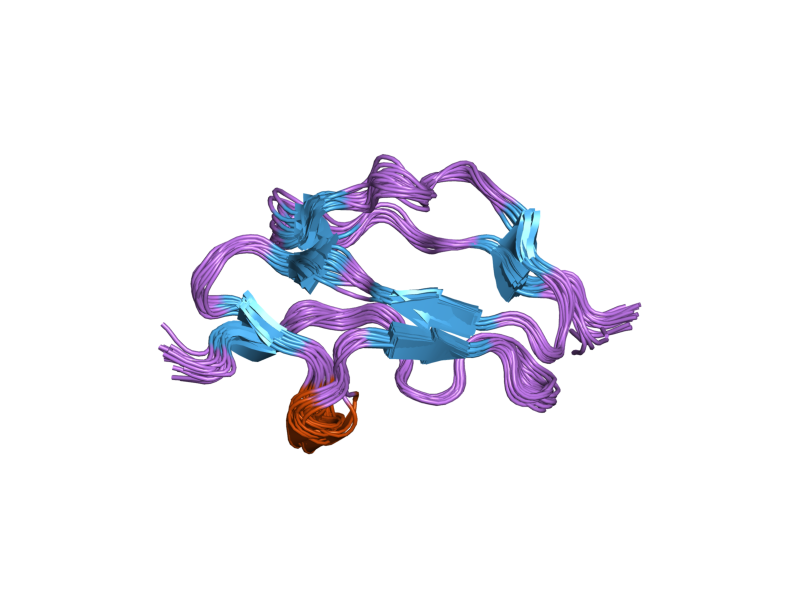
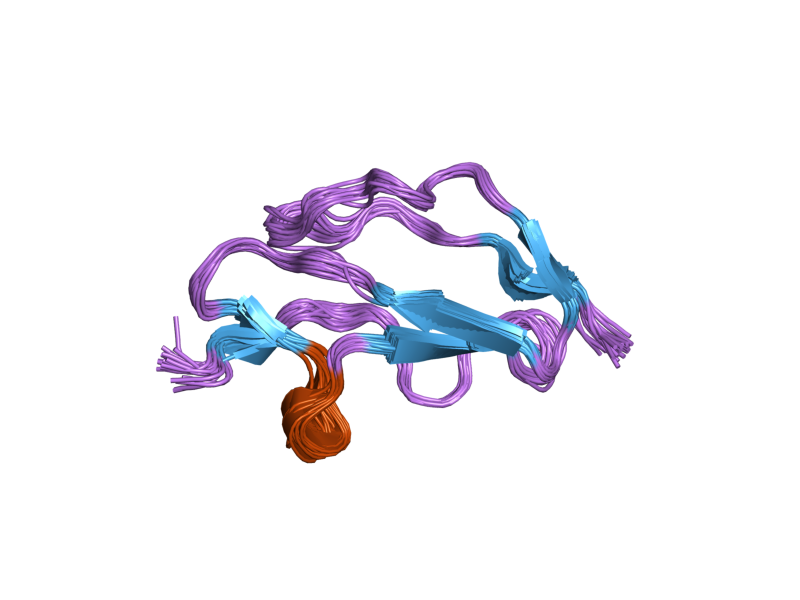
The molecule is made up of 20 complement control protein (CCP) modules (also referred to as Short Consensus Repeats or sushi domains) arranged head to tail. Each of the CCP modules consists of around 60 amino acids with four cysteine residues disulfide bonded in a 1-3 2-4 arrangement, and a hydrophobic core built around an almost invariant tryptophan residue. To date atomic structures have been determined for CCPs 1-3[3], CCP 5,[4] CCP 7 (both 402H & 402Y)[5], CCPs 12-13,[6] CCP 15, CCP 16,[7] CCPs 15-16,[8] and CCPs 19-20.[9][10] The atomic structure for CCPs 6-8 (402H) bound to the GAG mimic sucrose octasulfate[11] and CCPs 1-4 in complex with C3b[12] have also been determined. Although an atomic resolution structure for intact factor H has not yet been determined, low resolution techniques indicate that it may be bent back in solution.[13] Information available to date indicates that CCP modules 1-4 is responsible for the cofactor and decay acceleration activities of factor H, whereas self/non-self discrimination occurs predominantly through GAG binding to CCP modules 7 and/or 19-20.[13][14]
Clinical significance
Due to the central role that factor H plays in the regulation of complement, there are a number of clinical implications arrising from aberrant factor H activity. Overactive factor H may result in reduced complement activity on pathogenic cells - increasing susceptibility to microbial infections. Underactive factor H may result in increased complement activity on healthy host cells - resulting in autoimmune diseases. It is not surprising therefore that mutations or single nucleotide polymorphisms (SNPs) in factor H often result in pathologies. Moreover the complement inhibitory activities of factor H, and other complement regulators, are often used by pathogens to increase virulence.
Recently it was discovered that about 35% of individuals carry at an at-risk SNP in one or both copies of their factor H gene. Homozygous individuals have an approximately sevenfold increased chance of developing age-related macular degeneration, while heterozygotes have a two-to-threefold increased likelihood of developing the disease. This SNP, located in CCP module 7 of factor H, has been shown to affect the interaction between factor H and heparin indicating a causal relationship between the SNP and disease.[15][5] Deletion of two adjacent genes with a high degree of homology to complement factor H, named complement factor H-related 3 and complement factor H-related 1, protects against age-related macular degeneration because of reduced competition for binding of CFH to vascular surface binding sites.[16] [17]
Atypical haemolytic uraemic syndrome
Haemolytic uraemic syndrome (HUS) is a disease associated with microangiopathic haemolytic anemia, thrombocytopenia and acute renal failure. A rare subset of this disease (referred to as atypical haemolytic uraemic syndrome, aHUS), has been strongly linked to mutations in genes of the complement system (including factor H, factor I and membrane cofactor protein), with the factor H mutations being the most numerous. These factor H mutations tend to congregate towards the C-terminus of factor H—a region responsible for discriminating self from non-self and have been shown to disrupt heparin and C3d binding.[18]
Recruitment by pathogens
Given the central role of factor H in protecting cells from complement, it is not surprising that several important human pathogens have evolved mechanisms for recruiting factor H. This recruitment of factor H by pathogens provides significant resistance to complement attack, and therefore increased virulence. Pathogens that have been shown to recruit factor H include: Aspergillus spp.; Borrelia burgdorferi; B. duttonii; B. recurrentis; Candida albicans; Francisella tularensis; Haemophilus influenzae; Neisseria meningitidis; and Streptococcus pyogenes.
Interactions
Factor H has been shown to interact with Complement component 3.[19][20]
Recombinant production
Biologically active Factor H has been produced by Ralf Reski and coworkers in the moss bioreactor [21], in a process called molecular farming.
References
- ^ Pangburn MK (August 2000). "Host recognition and target differentiation by factor H, a regulator of the alternative pathway of complement". Immunopharmacology 49 (1–2): 149–57. doi:10.1016/S0162-3109(00)80300-8. PMID 10904114.
- ^ Rodríguez de Córdoba S, Esparza-Gordillo J, Goicoechea de Jorge E, Lopez-Trascasa M, Sánchez-Corral P (June 2004). "The human complement factor H: functional roles, genetic variations and disease associations". Mol. Immunol. 41 (4): 355–67. doi:10.1016/j.molimm.2004.02.005. PMID 15163532.
- ^ Hocking HG, Herbert AP, Kavanagh D, Soares DC, Ferreira VP, Pangburn MK, Uhrín D, Barlow PN (April 2008). "Structure of the N-terminal Region of Complement Factor H and Conformational Implications of Disease-linked Sequence Variations". J. Biol. Chem. 283 (14): 9475–87. doi:10.1074/jbc.M709587200. PMC 2276370. PMID 18252712. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2276370.
- ^ Barlow PN, Norman DG, Steinkasserer A, Horne TJ, Pearce J, Driscoll PC, Sim RB, Campbell ID. (1992). "Solution structure of the fifth repeat of factor H: a second example of the complement control protein module". Biochemistry. 31 (14): 3626–34. doi:10.1021/bi00129a011. PMID 1533152.
- ^ a b Herbert AP, Deakin JA, Schmidt CQ, Blaum BS, Egan C, Ferreira VP, Pangburn MK, Lyon M, Uhrín D, Barlow PN (June 2007). "Structure shows that a glycosaminoglycan and protein recognition site in factor H is perturbed by age-related macular degeneration-linked single nucleotide polymorphism". J. Biol. Chem. 282 (26): 18960–8. doi:10.1074/jbc.M609636200. PMID 17360715.
- ^ Schmidt CQ, Herbert AP, Mertens HD, Guariento M, Soares DC, Uhrin D, Rowe AJ, Svergun DI, Barlow PN. (2010). "The Central Portion of Factor H (Modules 10–15) Is Compact and Contains a Structurally Deviant CCP Module". J Mol Biol. 395 (1): 105–22. doi:10.1016/j.jmb.2009.10.010. PMC 2806952. PMID 19835885. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2806952.
- ^ Norman DG, Barlow PN, Baron M, Day AJ, Sim RB, Campbell ID. (1991). "Three-dimensional structure of a complement control protein module in solution". J Mol Biol. 219 (4): 717–25. doi:10.1016/0022-2836(91)90666-T. PMID 1829116.
- ^ Barlow PN, Steinkasserer A, Norman DG, Kieffer B, Wiles AP, Sim RB, Campbell ID. (1993). "Solution structure of a pair of complement modules by nuclear magnetic resonance". J Mol Biol. 232 (1): 268–84. doi:10.1006/jmbi.1993.1381. PMID 8331663.
- ^ Herbert AP, Uhrín D, Lyon M, Pangburn MK, Barlow PN. (march 2006). "Disease-associated sequence variations congregate in a polyanion recognition patch on human factor H revealed in three-dimensional structure". J Biol Chem. 281 (24): 16512–20. doi:10.1074/jbc.M513611200. PMID 16533809.
- ^ Jokiranta TS, Jaakola VP, Lehtinen MJ, Pärepalo M, Meri S, Goldman A. (April 2006). "Structure of complement factor H carboxyl-terminus reveals molecular basis of atypical haemolytic uremic syndrome". EMBO J. 25 (8): 1784–94. doi:10.1038/sj.emboj.7601052. PMC 1440827. PMID 16601698. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1440827.
- ^ Prosser BE, Johnson S, Roversi P, Herbert AP, Blaum BS, Tyrrell J, Jowitt TA, Clark SJ, Tarelli E, Uhrín D, Barlow PN, Sim RB, Day AJ, Lea SM. (September 2007). "Structural basis for complement factor H–linked age-related macular degeneration". J Exp Med. 204 (10): 2277–83. doi:10.1084/jem.20071069. PMC 2118454. PMID 17893204. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2118454.
- ^ Wu J, Wu YQ, Ricklin D, Janssen BJ, Lambris JD, Gros P. (2009). "Structure of C3b-factor H and implications for host protection by complement regulators". Nat Immunol. 10 (7): 728–33. doi:10.1038/ni.1755. PMC 2713992. PMID 19503104. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2713992.
- ^ a b Aslam M, Perkins SJ. (June 2001). "Folded-back solution structure of monomeric factor H of human complement by synchrotron X-ray and neutron scattering, analytical ultracentrifugation and constrained molecular modelling". J Mol Biol. 309 (25): 1117–1138. doi:10.1006/jmbi.2001.4720. PMID 11399083.
- ^ Kirkitadze MD, Barlow PN (April 2001). "Structure and flexibility of the multiple domain proteins that regulate complement activation". Immunol. Rev. 180: 146–61. doi:10.1034/j.1600-065X.2001.1800113.x. PMID 11414356.
- ^ Hageman GS, Anderson DH, Johnson LV, Hancox LS, Taiber AJ, Hardisty LI, Hageman JL, Stockman HA, Borchardt JD, Gehrs KM, Smith RJ, Silvestri G, Russell SR, Klaver CC, Barbazetto I, Chang S, Yannuzzi LA, Barile GR, Merriam JC, Smith RT, Olsh AK, Bergeron J, Zernant J, Merriam JE, Gold B, Dean M, Allikmets R (May 2005). "A common haplotype in the complement regulatory gene factor H (HF1/CFH) predisposes individuals to age-related macular degeneration". Proc. Natl. Acad. Sci. U.S.A. 102 (20): 7227–32. doi:10.1073/pnas.0501536102. PMC 1088171. PMID 15870199. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1088171.
- ^ Hughes, Anne E; Orr, Nick; Esfandiary, Hossein; Diaz-Torres, Martha; Goodship, Timothy; Chakravarthy, Usha (2006). "A common CFH haplotype, with deletion of CFHR1 and CFHR3, is associated with lower risk of age-related macular degeneration". Nature Genetics 38 (10): 1173–1177. doi:10.1038/ng1890. PMID 16998489.
- ^ Fritsche, L. G.; Lauer, N.; Hartmann, A.; Stippa, S.; Keilhauer, C. N.; Oppermann, M.; Pandey, M. K.; Kohl, J. et al. (2010). "An imbalance of human complement regulatory proteins CFHR1, CFHR3 and factor H influences risk for age-related macular degeneration (AMD)". Human Molecular Genetics 19 (23): 4694–4704. doi:10.1093/hmg/ddq399. PMID 20843825.
- ^ Atkinson JP, Goodship TH (June 2007). "Complement factor H and the hemolytic uremic syndrome". J. Exp. Med. 204 (6): 1245–8. doi:10.1084/jem.20070664. PMC 2118604. PMID 17548524. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2118604.
- ^ Soames, C J; Sim R B (Sep. 1997). "Interactions between human complement components factor H, factor I and C3b". Biochem. J. (ENGLAND) 326 ( Pt 2) (Pt 2): 553–61. ISSN 0264-6021. PMC 1218704. PMID 9291131. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1218704.
- ^ Jokiranta, T S; Westin J, Nilsson U R, Nilsson B, Hellwage J, Löfås S, Gordon D L, Ekdahl K N, Meri S (Mar. 2001). "Complement C3b interactions studied with surface plasmon resonance technique". Int. Immunopharmacol. (Netherlands) 1 (3): 495–506. doi:10.1016/S1567-5769(00)00042-4. ISSN 1567-5769. PMID 11367533.
- ^ Büttner-Mainik, A., J. Parsons, H. Jérome, A. Hartmann, S. Lamer, A. Schaaf, A. Schlosser, P.F. Zipfel, R. Reski, E.L. Decker (2011): Production of biologically active recombinant human factor H in Physcomitrella. Plant Biotechnology Journal, doi: 10.1111/j.1467-7652.2010.00552.x. [1]
Further reading
- Bradley, D T; Zipfel, P F; Hughes, A E (2011). "Complement in age-related macular degeneration: a focus on function". Eye 25 (6): 683–93. doi:10.1038/eye.2011.37. PMC 3178140. PMID 21394116. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3178140.
- Kardys, I. et al. (2006). "A common polymorphism in the complement factor h gene is associated with increased risk of myocardial infarction the rotterdam study". J. Am. Coll. Cardiol. 47 (8): 1568–75. doi:10.1016/j.jacc.2005.11.076. PMID 16630992.
- Pío R, Elsasser TH, Martínez A, Cuttitta F (2002). "Identification, characterization, and physiological actions of factor H as an adrenomedullin binding protein present in human plasma". Microsc. Res. Tech. 57 (1): 23–7. doi:10.1002/jemt.10047. PMID 11921353.
- Walport MJ; Rosen, Fred S.; Walport, Mark J. (2001). "Complement. First of two parts". N Engl J Med. 344 (14): 1058–66. doi:10.1056/NEJM200104053441406. PMID 11287977.
- Walport MJ; Rosen, Fred S.; Walport, Mark J. (2001). "Complement. Second of two parts". N Engl J Med. 344 (15): 1140–4. doi:10.1056/NEJM200104123441506. PMID 11297706.
PDB gallery  1hcc: THREE-DIMENSIONAL STRUCTURE OF A COMPLEMENT CONTROL PROTEIN MODULE IN SOLUTION
1hcc: THREE-DIMENSIONAL STRUCTURE OF A COMPLEMENT CONTROL PROTEIN MODULE IN SOLUTION 1hfh: SOLUTION STRUCTURE OF A PAIR OF COMPLEMENT MODULES BY NUCLEAR MAGNETIC RESONANCE
1hfh: SOLUTION STRUCTURE OF A PAIR OF COMPLEMENT MODULES BY NUCLEAR MAGNETIC RESONANCE 1hfi: SOLUTION STRUCTURE OF A PAIR OF COMPLEMENT MODULES BY NUCLEAR MAGNETIC RESONANCE
1hfi: SOLUTION STRUCTURE OF A PAIR OF COMPLEMENT MODULES BY NUCLEAR MAGNETIC RESONANCE 2bzm: SOLUTION STRUCTURE OF THE PRIMARY HOST RECOGNITION REGION OF COMPLEMENT FACTOR H
2bzm: SOLUTION STRUCTURE OF THE PRIMARY HOST RECOGNITION REGION OF COMPLEMENT FACTOR H 2g7i: Structure of Human Complement Factor H Carboxyl Terminal Domains 19-20: a Basis for Atypical Hemolytic Uremic Syndrome
2g7i: Structure of Human Complement Factor H Carboxyl Terminal Domains 19-20: a Basis for Atypical Hemolytic Uremic Syndrome 2jgw: STRUCTURE OF CCP MODULE 7 OF COMPLEMENT FACTOR H - THE AMD AT RISK VARIENT (402H)
2jgw: STRUCTURE OF CCP MODULE 7 OF COMPLEMENT FACTOR H - THE AMD AT RISK VARIENT (402H) 2jgx: STRUCTURE OF CCP MODULE 7 OF COMPLEMENT FACTOR H - THE AMD NOT AT RISK VARIENT (402Y)
2jgx: STRUCTURE OF CCP MODULE 7 OF COMPLEMENT FACTOR H - THE AMD NOT AT RISK VARIENT (402Y)External links
- GeneReviews/NCBI/NIH/UW entry on Atypical Hemolytic-Uremic Syndrome
- GeneReviews/NCBI/NIH/UW entry on Dense Deposit Disease/Membranoproliferative Glomerulonephritis Type II
- OMIM entries on Atypical Hemolytic-Uremic Syndrome
- MeSH Complement+Factor+H
Proteins: complement system (C, L, A) Activators/enzymes EarlyMiddleLateInhibitors CLA: C1-inhibitor - Decay-accelerating factor/CD59 - Factor I
CL: C4BP
A: Factor HComplement receptors Proteins: Globular proteins Serum globulins serpins: alpha-1 (Alpha 1-antichymotrypsin, Alpha 1-antitrypsin) · alpha-2 (Alpha 2-antiplasmin) · Antithrombin (Heparin cofactor II)
carrier proteins: alpha-1 (Transcortin) · alpha-2 (Ceruloplasmin) · Retinol binding protein
other: alpha-1 (Orosomucoid) · alpha-2 (alpha-2-Macroglobulin, Haptoglobin)carrier proteins: Sex hormone-binding globulin · Transferrin
other: Angiostatin · Hemopexin · Beta-2 microglobulin · Factor H · Plasminogen · ProperdinOtherOther globulins Albumins Othersee also disorders of globin and globulin proteins
B proteins: BY STRUCTURE: membrane, globular (en, ca, an), fibrousCategories:- Human proteins
- Complement system

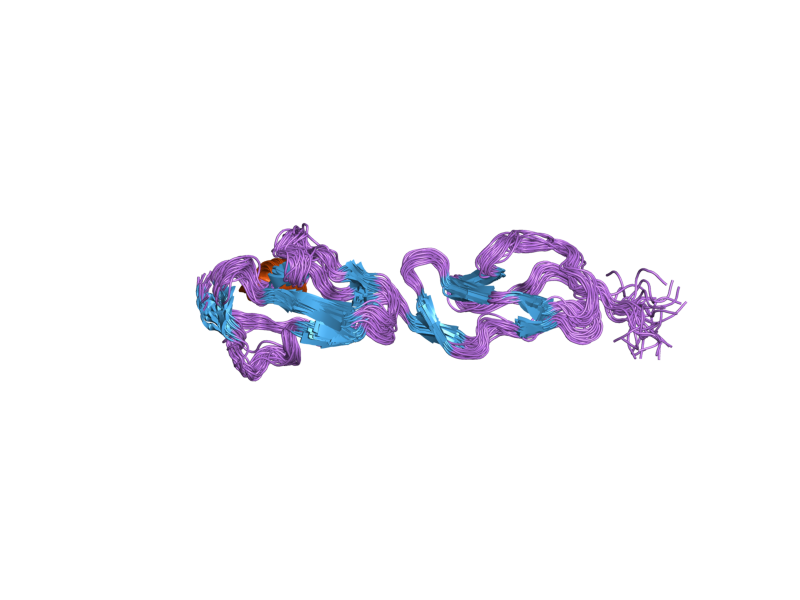
Wikimedia Foundation. 2010.