- Neural development
-
Neural development comprises the processes that generate, shape, and reshape the nervous system, from the earliest stages of embryogenesis to the final years of life. The study of neural development aims to describe the cellular basis of brain development and to address the underlying mechanisms. The field draws on both neuroscience and developmental biology to provide insight into the cellular and molecular mechanisms by which complex nervous systems develop. Defects in neural development can lead to cognitive, motor, and intellectual disability, as well as neurological disorders such as autism, Rett syndrome, and mental retardation.
Contents
Overview of brain development
The brain emerges during embryonic development from the neural tube, an early embryonic structure. The most anterior part of the neural tube is called the telencephalon, which expands rapidly due to cell proliferation, and eventually gives rise to the brain. Gradually some of the cells stop dividing and differentiate into neurons and glial cells, which are the main cellular components of the brain. The newly generated neurons migrate to different parts of the developing brain to self-organize into different brain structures. Once the neurons have reached their regional positions, they extend axons and dendrites, which allow them to communicate with other neurons via synapses. Synaptic communication between neurons leads to the establishment of functional neural circuits that mediate sensory and motor processing, and underlie behavior. The brain does most of its development within the first 20 years of life.
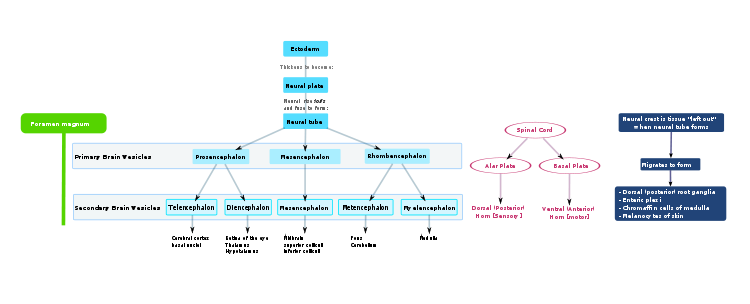 Flowchart of human brain development.
Flowchart of human brain development.
Aspects of neural development
Some landmarks of neural development include the birth and differentiation of neurons from stem cell precursors, the migration of immature neurons from their birthplaces in the embryo to their final positions, outgrowth of axons and dendrites from neurons, guidance of the motile growth cone through the embryo towards postsynaptic partners, the generation of synapses between these axons and their postsynaptic partners, and finally the lifelong changes in synapses, which are thought to underlie learning and memory.
Typically, these neurodevelopmental processes can be broadly divided into two classes: activity-independent mechanisms and activity-dependent mechanisms. Activity-independent mechanisms are generally believed to occur as hardwired processes determined by genetic programs played out within individual neurons. These include differentiation, migration and axon guidance to their initial target areas. These processes are thought of as being independent of neural activity and sensory experience. Once axons reach their target areas, activity-dependent mechanisms come into play. Although synapse formation is an activity-independent event, modification of synapses and synapse elimination requires neural activity.
Developmental neuroscience uses a variety of animal models including mice Mus musculus , the fruit fly Drosophila melanogaster , the zebrafish Danio rerio, Xenopus laevis tadpoles and the worm Caenorhabditis elegans, among others.
Neural induction
During early embryonic development the ectoderm becomes specified to give rise to the epidermis (skin) and the neural plate. The conversion of undifferentiated ectoderm to neuro-ectoderm requires signals from the mesoderm. At the onset of gastrulation presumptive mesodermal cells move through the dorsal blastopore lip and form a layer in between the endoderm and the ectoderm. These mesodermal cells that migrate along the dorsal midline give rise to a structure called the notochord. Ectodermal cells overlying the notochord develop into the neural plate in response to a diffusible signal produced by the notochord. The remainder of the ectoderm gives rise to the epidermis (skin). The ability of the mesoderm to convert the overlying ectoderm into neural tissue is called Neural Induction.
The neural plate folds outwards during the third week of gestation to form the neural groove. Beginning in the future neck region, the neural folds of this groove close to create the neural tube. The formation of the neural tube from the ectoderm is called Neurulation. The ventral part of the neural tube is called the basal plate; the dorsal part is called the alar plate. The hollow interior is called the neural canal. By the end of the fourth week of gestation, the open ends of the neural tube (the neuropores) close off.[1]
Identification of neural inducers
A transplanted blastopore lip can convert ectoderm into neural tissue and is said to have an inductive effect. Neural Inducers are molecules that can induce the expression of neural genes in ectoderm explants without inducing mesodermal genes as well. Neural induction is often studied in Xenopus embryos since they have a simple body pattern and there are good markers to distinguish between neural and non-neural tissue. Examples of Neural Inducers are the molecules Noggin and Chordin.
When embryonic ectodermal cells are cultured at low density in the absence of mesodermal cells they undergo neural differentiation (express neural genes), suggesting that neural differentiation is the default fate of ectodermal cells. In explant cultures (which allow direct cell-cell interactions) the same cells differentiate into epidermis. This is due to the action of BMP4 (a TGF-β family protein) that induces ectodermal cultures to differentiate into epidermis. During neural induction, Noggin and Chordin are produced by the dorsal mesoderm (notochord) and diffuse into the overlying ectoderm to inhibit the activity of BMP4. This inhibition of BMP4 causes the cells to differentiate into neural cells.
Regionalization
Late in the fourth week, the superior part of the neural tube flexes at the level of the future midbrain—the mesencephalon. Above the mesencephalon is the prosencephalon (future forebrain) and beneath it is the rhombencephalon (future hindbrain).
The optical vesicle (which will eventually become the optic nerve, retina and iris) forms at the basal plate of the prosencephalon. The alar plate of the prosencephalon expands to form the cerebral hemispheres (the telencephalon) whilst its basal plate becomes the diencephalon. Finally, the optic vesicle grows to form an optic outgrowth.
Patterning of the nervous system
In chordates, dorsal ectoderm forms all neural tissue and the nervous system. Patterning occurs due to specific environmental conditions - different concentrations of signaling molecules
Dorsoventral axis [2]
The ventral half of the neural plate is controlled by the notochord, which acts as the 'organiser'. The dorsal half is controlled by the ectoderm plate which flanks the neural plate on either side.
Ectoderm follows a default pathway to become neural tissue. Evidence for this comes from single, cultured cells of ectoderm which go on to form neural tissue. This is postulated to be because of a lack of BMPs, which are blocked by the organiser. The organiser may produce molecules such as follistatin, noggin and chordin which inhibit BMPs.
The ventral neural tube is patterned by Sonic Hedgehog (Shh) from the notochord, which acts as the inducing tissue. Notochord-derived Shh signals to the floor plate, and induces Shh expression in the floor plate. Floor plate-derived Shh subsequently signals to other cells in the neural tube, and is essential for proper specification of ventral neuron progenitor domains. Loss of Shh from the notochord and/or floor plate prevents proper specification of these progenitor domains. Shh binds Patched1, relieving Patched-mediated inhibition of Smoothened, leading to activation of Gli family of transcription factors (Gli1, Gli2, and Gli3) transcription factors.
In this context Shh acts as a morphogen - it induces cell differentiation dependent on its concentration. At low concentrations it forms ventral interneurones, at higher concentrations it induces motor neuron development, and at highest concentrations it induces floor plate differentiation. Failure of Shh-modulated differentiation causes holoprosencephaly.
The dorsal neural tube is patterned by BMPs from the epidermal ectoderm flanking the neural plate. These induce sensory interneurones by activating Sr/Thr kinases and altering SMAD transcription factor levels.
Rostrocaudal (Anteroposterior) axis
Signals that control anteroposterior neural development include FGF and retinoic acid which act in the hindbrain and spinal cord.[3] The hindbrain, for example, is patterned by Hox genes, which are expressed in overlapping domains along the anteroposterior axis under the control of retinoic acid. The 3' genes in the Hox cluster are induced by retinoic acid in the hindbrain, whereas the 5' Hox genes are not induced by retinoic acid and are expressed more posteriorly in the spinal cord. Hoxb-1 is expressed in rhombomere 4 and gives rise to the facial nerve. Without this Hoxb-1 expression, a nerve which is similar to the trigeminal nerve arises.
Neuronal migration
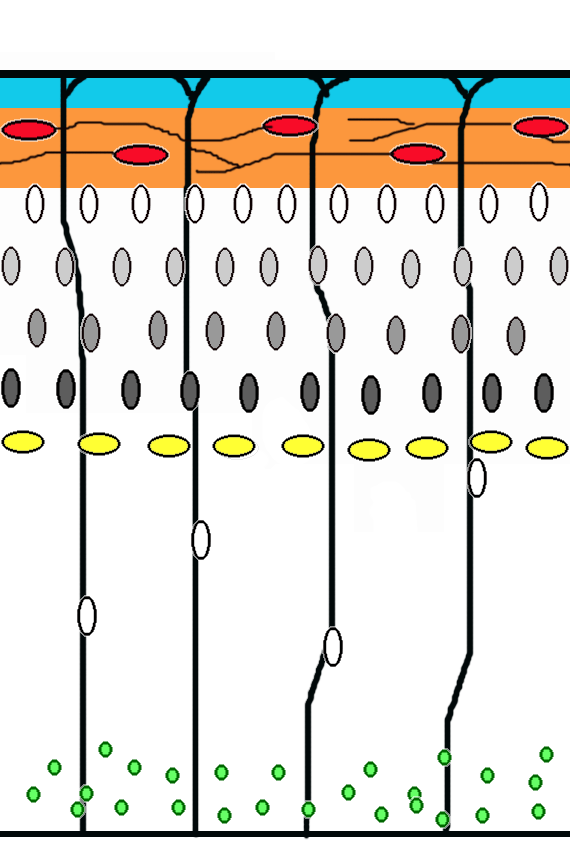
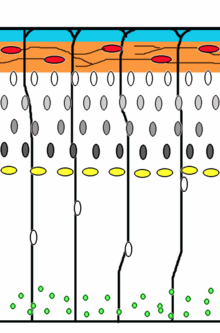 Corticogenesis: younger neurons migrate past older ones using radial glia as a scaffolding. Cajal-Retzius cells (red) release reelin (orange).
Corticogenesis: younger neurons migrate past older ones using radial glia as a scaffolding. Cajal-Retzius cells (red) release reelin (orange).Neuronal migration is the method by which neurons travel from their origin or birth place to their final position in the brain. There are several ways they can do this, e.g. by radial migration or tangential migration. (see time lapse sequences of radial migration (also known as glial guidance) and somal translocation.)[4]

Radial migration Neuronal precursor cells proliferate in the ventricular zone of the developing neocortex. The first postmitotic cells to migrate form the preplate which are destined to become Cajal-Retzius cells and subplate neurons. These cells do so by somal translocation. Neurons migrating with this mode of locomotion are bipolar and attach the leading edge of the process to the pia. The soma is then transported to the pial surface by nucleokinesis, a process by which a microtubule "cage" around the nucleus elongates and contracts in association with the centrosome to guide the nucleus to its final destination.[5] Radial glia, whose fibers serve as a scaffolding for migrating cells, can itself divide[6] or translocate to the cortical plate and differentiate either into astrocytes or neurons.[7] Somal translocation can occur at any time during development.[4]
Subsequent waves of neurons split the preplate by migrating along radial glial fibres to form the cortical plate. Each wave of migrating cells travel past their predecessors forming layers in an inside-out manner, meaning that the youngest neurons are the closest to the surface.[8][9] It is estimated that glial guided migration represents 90% of migrating neurons in human and about 75% in rodents.[10]
Tangential migration Most interneurons migrate tangentially through multiple modes of migration to reach their appropriate location in the cortex. An example of tangential migration is the movement of interneurons from the ganglionic eminence to the cerebral cortex. One example of ongoing tangential migration in a mature organism, observed in some animals, is the rostral migratory stream connecting subventricular zone and olfactory bulb.
Others modes of migration There is also a method of neuronal migration called multipolar migration.[11][12] This is seen in multipolar cells, which are abundantly present in the cortical intermediate zone. They do not resemble the cells migrating by locomotion or somal translocation. Instead these multipolar cells express neuronal markers and extend multiple thin processes in various directions independently of the radial glial fibers.[11]
Neurotrophic factors
The survival of neurons is regulated by survival factors, called trophic factors. The neurotrophic hypothesis was formulated by Victor Hamburger and Rita Levi Montalcini based on studies of the developing nervous system. Victor Hamburger discovered that implanting an extra limb in the developing chick led to an increase in the number of spinal motor neurons. Initially he thought that the extra limb was inducing proliferation of motor neurons, but he and his colleagues later showed that there was a great deal of motor neuron death during normal development, and the extra limb prevented this cell death. According to the neurotrophic hypothesis, growing axons compete for limiting amounts of target-derived trophic factors and axons that neurons that fail to receive insufficient trophic support die by apoptosis. It is now clear that factors produced by a number of sources contribute to neuronal survival.
Nerve Growth Factor (NGF): Rita Levi Montalcini and Stanley Cohen purified the first trophic factor, Nerve Growth Factor (NGF), for which they received the Nobel Prize. There are three NGF-related trophic factors: BDNF, NT3, and NT4, which regulate survival of various neuronal populations. The Trk proteins act as receptors for NGF and related factors. Trk is a receptor tyrosine kinase. Trk dimerization and phosphorylation leads to activation of various intracellular signaling pathways including the MAP kinase, Akt, and PKC pathways.
CNTF: Ciliary neurotrophic factor is another protein that acts as a survival factor for motor neurons. CNTF acts via a receptor complex that includes CNTFRα, GP130, and LIFRβ. Activation of the receptor leads to phosphorylation and recruitment of the JAK kinase, which in turn phosphorylates LIFRβ. LIFRβ acts as a docking site for the STAT transcription factors. JAK kinase phosphorylates STAT proteins, which dissociate from the receptor and translocate to the nucleus to regulate gene expression.
GDNF: Glial derived neurotrophic factor is a member of the TGFb family of proteins, and is a potent trophic factor for striatal neurons. The functional receptor is a heterodimer, composed of type 1 and type 2 receptors. Activation of the type 1 receptor leads to phosphorylation of Smad proteins, which translocate to the nucleus to activate gene expression.
Synapse formation
Neuromuscular junction Much of our understanding of synapse formation comes from studies at the neuromuscular junction. The transmitter at this synapse is acetylcholine. The acetylcholine receptor (AchR) is present at the surface of muscle cells before synapse formation. The arrival of the nerve induces clustering of the receptors at the synapse. McMahan and Sanes showed that the synaptogenic signal is concentrated at the basal lamina. They also showed that the synaptogenic signal is produced by the nerve, and they identified the factor as Agrin. Agrin induces clustering of AchRs on the muscle surface and synapse formation is disrupted in agrin knockout mice. Agrin transuces the signal via MuSK receptor to rapsyn. Fischbach and colleagues showed that receptor subunits are selectively transcribed from nuclei next to the synaptic site. This is mediated by neuregulins.
In the mature synapse each muscle fiber is innervated by one motor neuron. However, during development many of the fibers are innervated by multiple axons. Lichtman and colleagues have studied the process of synapses elimination. This is an activity-dependent event. Partial blockage of the receptor leads to retraction of corresponding presynaptic terminals.
CNS synapses Agrin appears not to be a central mediator of CNS synapse formation and there is active interest in identifying signals that mediate CNS synaptogenesis. Neurons in culture develop synapses that are similar to those that form in vivo, suggesting that synaptogenic signals can function properly in vitro. CNS synaptogenesis studies have focused mainly on glutamatergic synapses. Imaging experiments show that dendrites are highly dynamic during development and often initiate contact with axons. This is followed by recruitment of postsynaptic proteins to the site of contact. Stephen Smith and colleagues have shown that contact initiated by dendritic filopodia can develop into synapses.
Induction of synapse formation by glial factors: Barres and colleagues made the observation that factors in glial conditioned media induce synapse formation in retinal ganglion cell cultures. Synapse formation in the CNS is correlated with astrocyte differentiation suggesting that astrocytes might provide a synaptogenic factor. The identity of the astrocytic factors is not yet known.
Neuroligins and SynCAM as synaptogenic signals: Sudhof, Serafini, Scheiffele and colleagues have shown that neuroligins and SynCAM can act as factors that will induce presynaptic differentiation. Neuroligins are concentrated at the postsynaptic site and act via neurexins concentrated in the presynaptic axons. SynCAM is a cell adhesion molecule that is present in both pre- and post-synaptic membranes.
Synapse elimination
Several motorneurones compete for each neuromuscular junction, but only one survives till adulthood. Competition in vitro has been shown to involve a limited neurotrophic substance that is released, or that neural activity infers advantage to strong post-synaptic connections by giving resistance to a toxin also released upon nerve stimulation. In vivo it is suggested that muscle fibres select the strongest neuron through a retrograde signal.
See also
- Neural development in humans
- Axon guidance
- Pioneer neuron
- Neural Darwinism
- Neurodevelopmental disorder
- Pre- and perinatal psychology
- Brain development timelines
- Malleable intelligence
- Human brain development timeline
- Mouse brain development timeline
- Macaque brain development timeline
References
- ^ Estomih Mtui; Gregory Gruener (2006). Clinical Neuroanatomy and Neuroscience. Philadelphia: Saunders. pp. 1. ISBN 1-4160-3445-5.
- ^ Jessell, Thomas M.; Kandel, Eric R.; Schwartz, James H. (2000). "Chapter 55". Principles of neural science (4th ed.). New York: McGraw-Hill. ISBN 0-8385-7701-6.
- ^ Duester, G (September 2008). "Retinoic acid synthesis and signaling during early organogenesis". Cell 134 (6): 921–31. doi:10.1016/j.cell.2008.09.002. PMC 2632951. PMID 18805086. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2632951.
- ^ a b Nadarajah B, Brunstrom J, Grutzendler J, Wong R, Pearlman A (2001). "Two modes of radial migration in early development of the cerebral cortex". Nat Neurosci 4 (2): 143–50. doi:10.1038/83967. PMID 11175874. http://www.nature.com/neuro/journal/v4/n2/full/nn0201_143.html.
- ^ Samuels B, Tsai L (2004). "Nucleokinesis illuminated". Nat Neurosci 7 (11): 1169–70. doi:10.1038/nn1104-1169. PMID 15508010. http://www.nature.com/neuro/journal/v7/n11/full/nn1104-1169.html.
- ^ Tamamaki N, Nakamura K, Okamoto K, Kaneko T (September 2001). "Radial glia is a progenitor of neocortical neurons in the developing cerebral cortex". Neurosci. Res. 41 (1): 51–60. doi:10.1016/S0168-0102(01)00259-0. PMID 11535293. http://linkinghub.elsevier.com/retrieve/pii/S0168010201002590.
- ^ Miyata T, Kawaguchi A, Okano H, Ogawa M (September 2001). "Asymmetric inheritance of radial glial fibers by cortical neurons". Neuron 31 (5): 727–41. doi:10.1016/S0896-6273(01)00420-2. PMID 11567613. http://linkinghub.elsevier.com/retrieve/pii/S0896-6273(01)00420-2.
- ^ Nadarajah B, Parnavelas J (2002). "Modes of neuronal migration in the developing cerebral cortex". Nat Rev Neurosci 3 (6): 423–32. doi:10.1038/nrn845. PMID 12042877.
- ^ Rakic P (1972). "Mode of cell migration to the superficial layers of fetal monkey neocortex". J Comp Neurol 145 (1): 61–83. doi:10.1002/cne.901450105. PMID 4624784.
- ^ Letinic K, Zoncu R, Rakic P (June 2002). "Origin of GABAergic neurons in the human neocortex". Nature 417 (6889): 645–9. doi:10.1038/nature00779. PMID 12050665.
- ^ a b Tabata H, Nakajima K (5 November 2003). "Multipolar migration: the third mode of radial neuronal migration in the developing cerebral cortex". J Neurosci 23 (31): 9996–10001. PMID 14602813. http://www.jneurosci.org/cgi/content/full/23/31/9996.
- ^ Nadarajah B, Alifragis P, Wong R, Parnavelas J (2003). "Neuronal migration in the developing cerebral cortex: observations based on real-time imaging". Cereb Cortex 13 (6): 607–11. doi:10.1093/cercor/13.6.607. PMID 12764035. http://cercor.oxfordjournals.org/cgi/content/full/13/6/607.
External links
- Neural Development (peer-reviewed open access journal).
- Translating Neurodevelopmental Time Across Mammalian Species
Prenatal development/Mammalian development of nervous system (GA 9.733 and GA 10.1002, TE E5.13-16) Neurogenesis Rostral neuropore
Cephalic flexure · Pontine flexure
Alar plate (sensory) · Basal plate (motor)
Germinal matrixEye development Auditory development M: EYE
anat(g/a/p)/phys/devp/prot
noco/cong/tumr, epon
proc, drug(S1A/1E/1F/1L)
M: EAR
anat(e/p)/phys/devp
noco/cong, epon
proc, drug(S2)
Categories:- Developmental biology
- Embryology of nervous system
- Developmental neuroscience

Wikimedia Foundation. 2010.