- Feather
-
For other uses, see Feather (disambiguation).
 Feather variations
Feather variations
Feathers are one of the epidermal growths that form the distinctive outer covering, or plumage, on birds and some theropod dinosaurs. They are considered the most complex integumentary structures found in vertebrates,[1][2] and indeed a premier example of a complex evolutionary novelty.[3] They are among the characteristics that distinguish the extant Aves from other living groups. Feathers have also been noticed in those Theropoda which have been termed feathered dinosaurs. Although feathers cover most parts of the body of birds, they arise only from certain well-defined tracts on the skin. They aid in flight, thermal insulation, waterproofing and coloration that helps in communication and protection.[4]
Contents
Structure and characteristics
 Parts of a feather:
Parts of a feather:
1. Vane
2. Rachis
3. Barb
4. Afterfeather
5. Hollow shaft, calamus Featherstructure of a Blue-and-yellow Macaw
Featherstructure of a Blue-and-yellow Macaw Budgerigar feather, magnified, showing interlocking barbules
Budgerigar feather, magnified, showing interlocking barbulesFeathers are among the most complex integumentary appendages found in vertebrates and are formed in tiny follicles in the epidermis, or outer skin layer, that produce keratin proteins. The β-keratins in feathers, beaks and claws — and the claws, scales and shells of reptiles — are composed of protein strands hydrogen-bonded into β-pleated sheets, which are then further twisted and crosslinked by disulfide bridges into structures even tougher than the α-keratins of mammalian hair, horns and hoof.[5][6] The exact signals that induce the growth of feathers on the skin are not known but it has been found that the transcription factor cDermo-1 induces the growth of feathers on skin and scales on the leg.[7]
 Feather microstructure showing interlocking barbules.
Feather microstructure showing interlocking barbules.Classification
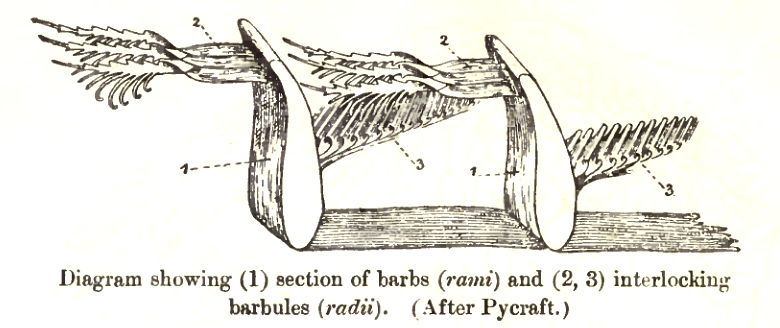
See also: Flight feather and Down featherThere are two basic types of feather: vaned feathers which cover the exterior of the body, and down feathers which are underneath the vaned feathers. The pennaceous feathers are vaned feathers. Also called contour feathers, pennaceous feathers arise from tracts and cover the whole body. A third rarer type of feathers, filoplumes, is hairlike and (if present in a bird) grows along the fluffy down feathers. In some passerines, filoplumes arise exposed beyond the contour feathers on the neck.[1] The remiges, or flight feathers of the wing, and rectrices, the flight feathers of the tail are the most important feathers for flight. A typical vaned feather features a main shaft, called the rachis. Fused to the rachis are a series of branches, or barbs; the barbs themselves are also branched and form the barbules. These barbules have minute hooks called barbicels for cross-attachment. Down feathers are fluffy because they lack barbicels, so the barbules float free of each other, allowing the down to trap much air and provide excellent thermal insulation. At the base of the feather, the rachis expands to form the hollow tubular calamus (or quill) which inserts into a follicle in the skin. The basal part of the calamus is without vanes. This part is embedded within the skin follicle and has an opening at the base (proximal umbilicus) and a small opening on the side (distal umbilicus).[8]
Hatchling birds of some species have a special kind of natal down (neossoptiles) and these are pushed out when the normal feathers (teleoptiles) emerge.[1]
Flight feathers are stiffened so as to work against the air in the downstroke but yield in other directions. It is noted that the pattern of orientation of β-keratin fibers in the feathers of flying birds differs from that in flightless birds. The fibers are better aligned in the middle of the feather and less aligned towards the tips.[9][10]
Functions
Feathers insulate birds from water and cold temperatures. They may also be plucked to line the nest and provide insulation to the eggs and young. The individual feathers in the wings and tail play important roles in controlling flight. Some species have a crest of feathers on their heads. Although feathers are light, a bird's plumage weighs two or three times more than its skeleton, since many bones are hollow and contain air sacs. Color patterns serve as camouflage against predators for birds in their habitats, and by predators looking for a meal. As with fish, the top and bottom colors may be different to provide camouflage during flight. Striking differences in feather patterns and colors are part of the sexual dimorphism of many bird species and are particularly important in selection of mating pairs. In some cases there are differences in the UV reflectivity of feathers across sexes even though no differences in color are noted in the visible range.[11] The wing feathers of male Club-winged Manakins Machaeropterus deliciosus have special structures that are used to produce sounds by stridulation.[12]
 A contour feather from a Guinea fowl
A contour feather from a Guinea fowlSome birds have a supply of powder down feathers which grow continuously, with small particles regularly breaking off from the ends of the barbules. These particles produce a powder that sifts through the feathers on the bird's body and acts as a waterproofing agent and a feather conditioner. Powder down has evolved independently in several taxa and can be found in down as well as pennaceous feathers. They may be scattered in plumage in the pigeons and parrots or in localized patches on the breast, belly or flanks as in herons and frogmouths. Herons use their bill to break the feathers and to spread them while cockatoos may use their head as a powder puff to apply the powder.[13] Waterproofing can be lost by exposure to emulsifying agents due to human pollution. Feathers can become waterlogged and birds may sink. It is also very difficult to clean and rescue birds whose feathers have been fouled by oil spills. The feathers of cormorants soak up water and help in reducing buoyancy and thereby allowing the birds to swim submerged.[14]
 Rictal bristles of a White-cheeked Barbet
Rictal bristles of a White-cheeked BarbetBristles are stiff, tapering feathers with a large rachis but few barbs. Rictal bristles are bristles found around the eyes and bill. They may serve a similar purpose to eyelashes and vibrissae in mammals. It has been suggested that they may aid insectivorous birds in prey capture or that it may have sensory functions, however there is no clear evidence.[15] In one study, Willow Flycatchers (Empidonax traillii) were found to catch insects equally well before and after removal of the rictal bristles.[16]
Grebes are peculiar in their habit of ingesting their own feathers and also feeding them to their young. Observations on the diet and feather eating frequency suggest that ingesting feathers particularly down from their flanks aids in forming easily ejectable pellets along with their diet of fish.[17]
Distribution
 Feather tracts or pterylae and their naming
Feather tracts or pterylae and their namingContour feathers are not uniformly distributed on the skin of the bird except in some groups such as the Penguins, ratites and screamers.[18] In most birds the feathers grow from specific tracts of skin called pterylae while there are regions which are free of feathers called apterylae. Filoplumes and down may arise from the apteriae, regions between the pterylae. The arrangement of these feather tracts, pterylosis or pterylography, varies across bird families and has been used in the past as a means for determining the evolutionary relationships of bird families.[19][20]
Coloration
 Colors resulting from different feather pigments
Colors resulting from different feather pigments
Left: turacin (red) and turacoverdin (green, with some structural blue iridescence at lower end) on the wing of Tauraco bannermani
Right: carotenoids (red) and melanins (dark) on belly/wings of Ramphocelus bresiliusThe colors of feathers are produced by the presence of pigments, or by microscopic refractive structures, or by a combination of both.
Most feather pigments are melanins (brown and beige pheomelanins, black and grey eumelanins) and carotenoids (red, yellow, orange); other pigments occur only in certain taxa – the yellow to red psittacofulvins[21] (found in some parrots) and the red turacin and green turacoverdin (porphyrin pigments found only in turacos). Structural coloration[4][22][23] is involved in the production of blue colors, iridescence, most ultraviolet reflectance and in the enhancement of pigmentary colors; structural iridescence has been reported[24] in fossil feathers dating back 40 million years. White feathers lack pigment and scatter light diffusely; albinism in birds is caused by defective pigment production, though structural coloration will not be affected (as can be seen e.g. in blue-and-white budgerigars).
 A feather with no pigment
A feather with no pigmentFor example, the blues and bright greens of many parrots are produced by constructive interference of light reflecting from different layers of the structures in feathers, in the case of green plumage in addition to the yellow pigments; the specific feather structure involved is sometimes called the Dyck texture.[25][26] Melanin is often involved in the absorption of some of the light; in combination with yellow pigment it produces dull olive-greens.
In some birds, the feather colors may be created or altered by uropygial gland secretions. The yellow bill colors of many hornbills are produced by preen gland secretions. Other differences that may only be visible in the ultraviolet region have been suggested[13] but studies have failed to find evidence.[27] Uropygial oil secretion may also have an inhibitory effect on feather bacteria.[28]
A bird's feathers undergo wear and tear and are replaced periodically during its life through molting. New feathers, known as blood, or pin feathers (depending on the stage of growth) when developing, are formed through the same follicle from which the old ones were fledged. The presence of melanin in feathers increases their resistance to abrasion.[29] One study notes that melanin based feathers were observed to degrade more quickly under bacterial action, even compared to unpigmented feathers from the same species, than those unpigmented or with carotenoid pigments.[30] However, another study the same year compared the action of bacteria on pigmentations of two song sparrow species and observed that the darker pigmented feathers were more resistant and they cited other research also published in 2004 that stated increased melanin provided greater resistance. They observed that the greater resistance of the darker birds confirmed Gloger's rule.[31] The evolution of coloration is based on sexual selection and it has been suggested that carotenoid-based pigments may have evolved since they are likely to be more honest signals of fitness because they are derived from special diets,[32][33] or because carotenoids are also required for immune function.[34]
Parasites
The feather surface is the home for some ectoparasites, notably feather lice (Phthiraptera) and feather mites. Feather lice typically live on a single host and can move only from parents to chicks or mating birds and occasionally by phoresy. This life history has resulted in most of the species being specific to the host and coevolving with the host, making them of interest in phylogenetic studies.[35]
Feather holes are chewing traces of lice (most probably Brueelia spp. lice) on the wing and tail feathers. They were described on barn swallows, and because of easy countability, many evolutionary, ecological, and behavioral publications use them to quantify the intensity of infestation.
Interestingly, parasitic cuckoos which grow up in the nests of other species also have host specific feather lice and these seem to be transmitted only after they leave the host nest.[36]
Birds maintain their feather condition by bathing in water, dust bathing and preening. A peculiar behavior of birds, anting, where ants are introduced into the plumage was suggested to help in reducing parasites but no supporting evidence has been found.[37]
Human usage
 Shaft of Indian Peacock tail feather
Shaft of Indian Peacock tail featherFeathers have a number of utilitarian, cultural and religious uses.
Utilitarian functions
Feathers are both soft and excellent at trapping heat; thus, they are sometimes used in high-class bedding, especially pillows, blankets, and mattresses. They are also used as filling for winter clothing, such as quilted coats and sleeping bags; goose and eider down have great loft, the ability to expand from a compressed, stored state to trap large amounts of compartmentalized, insulating air.[38]
Bird feathers have long been used for fletching arrows. Colorful feathers such as those belonging to pheasants have been used to decorate fishing lures.
Feathers of large birds (most often geese) have been and are used to make quill pens. The word pen itself is derived from the Latin penna for feather.[39] In French, plume can mean either feather or pen.
Feathers are also valuable in aiding the identification of species in forensic studies, particularly in bird strikes to aircraft. The ratios of hydrogen isotopes in feathers help in determining the geographic origins of birds.[40] Feathers may also be useful in the non-destructive sampling of pollutants.[41]
The poultry industry produces a large amount of feathers as waste, and like other forms of keratin, these are slow in their decomposition. Feather waste has been used in a number of industrial applications as a medium for culturing microbes,[42] biodegradeable polymers,[43] and production of enzymes.[44] Feather proteins have been tried as an adhesive for wood board.[45]
In religion and culture
Eagle feathers have great cultural and spiritual value to American Indians in the USA and First Nations peoples in Canada as religious objects. In the United States the religious use of eagle and hawk feathers are governed by the eagle feather law, a federal law limiting the possession of eagle feathers to certified and enrolled members of federally recognized Native American tribes.
Various birds and their plumages serve as cultural icons throughout the world, from the hawk in ancient Egypt to the bald eagle and the turkey (bird) in the United States. In Greek mythology, Daedelus the inventor and Icarus tried to escape his prison by attaching feathered wings to his shoulders with wax, which was melted by the Sun.
In South America, brews made from the feathers of Condors are used in traditional medications.[46] In India, feathers of the Indian Peacock have been used in traditional medicine for snakebite, infertility and coughs.[47][48]
During the 18th, 19th, and even 20th Centuries a booming international trade in plumes, to satisfy market demand in North America and Europe for extravagant head-dresses as adornment for fashionable women, caused so much destruction (for example, to egret breeding colonies) that a major campaign against it by conservationists led to the Lacey Act and caused the fashion to change and the market to finally collapse. Frank Chapman noted in 1886 that as many as 40 species of birds were used in about three-fourths of the 700 ladies' hats that he observed in New York City.[49][50][51]
More recently, rooster plumage has become a popular trend as a hairstyle accessory, with feathers formerly used solely as fishing lures being now used to provide color and style to hair.[52]
Evolution
Main article: Origin of avian flight Fossil feather of Archaeopteryx
Fossil feather of ArchaeopteryxThe functional view on the evolution of feathers has traditionally focused on insulation, flight and display. Discoveries of non-flying Late Cretaceous feathered dinosaurs in China however suggest that flight could not have been the original primary function.[53] There have been suggestions that feathers may have had their original function in thermoregulation, waterproofing or even as sinks for metabolic wastes such as sulphur.[54] While feathers have been suggested as having evolved from reptilian scales, there are numerous objections, and more recent explanations have arisen from the paradigm of evolutionary developmental biology.[2] Theories of the scale-based origins of feathers suggest that the planar scale structure was modified for their development into feathers by splitting to form the webbing; however, the developmental process involves a tubular structure arising from a follicle and the tube splitting longitudinally to form the webbing.[1][2] The number of feathers per unit area of skin is higher in smaller birds than in larger birds, and this trend indicates their important role in thermal insulation, since smaller birds lose more heat due to the relatively larger surface area in proportion to their body weight.[4] The miniaturization of birds also played a role in the evolution of powered flight.[55] The coloration of feathers is believed to be primarily evolved in response to sexual selection. In many cases the physiological condition of the birds (especially males) is indicated by the quality of their feathers and this is used (by the females) in mate choice.[56][57]
Feathered dinosaurs
Main article: Feathered dinosaurs Archaeopteryx lithographica (Berlin specimen)
Archaeopteryx lithographica (Berlin specimen)Several non-avian dinosaurs had feathers on their limbs that would not have functioned for flight.[2] One theory is that feathers originally evolved on dinosaurs as a result of insulation properties; those small dinosaurs that then grew longer feathers may have found them helpful in gliding, leading to the evolution of proto-birds like Archaeopteryx and Microraptor zhaoianus. Dinosaurs that had feathers or protofeathers include Pedopenna daohugouensis, [58] and Dilong paradoxus, a tyrannosauroid which is 60 to 70 million years older than Tyrannosaurus rex.[59]
The majority of dinosaurs known to have had feathers or protofeathers are saurischians, however featherlike "filamentous integumentary structures" are also known from the ornithischians Tianyulong and Psittacosaurus.[60] The exact nature of these structures is still under study. However, it is believed that the stage 1 feathers such as those seen in these two ornithischians likely functioned in display. [61]
Since the 1990s, dozens of feathered dinosaurs have been discovered in the clade Maniraptora, which includes the clade Avialae and the recent common ancestors of birds, Oviraptorosauria and Deinonychosauria. In 1998, the discovery of a feathered oviraptorosaurian, Caudipteryx zoui, challenged the notion that feathers were an exclusive structure of Avialae.[62] Buried in the Yixian Formation in Liaoning, China, C. zoui lived during the Early Cretaceous Period. Present on the forelimbs and tails, their integumentary structure has been accepted as pennaceous vaned feathers based on the rachis and herringbone pattern of the barbs. In the clade Deinonychosauria, the continued divergence of feathers is also apparent in the families Troodontidae and Dromaeosauridae. Branched feathers with ranchis, barbs, and barbules were discovered in many members including Sinornithosaurus millenii, a dromaeosaurid found in the Yixian formation (124.6 MYA).[63]
Previously, a temporal paradox existed in the evolution of feathers - theropods with highly derived bird-like characteristics occurred at a later time than Archaeopteryx, suggesting that the descendants of birds arose before the ancestor. However, this paradox was resolved in 2009 with the discovery of Anchiornis huxleyi, found in the Late Jurassic Tiaojishan Formation (160 MYA) in western Liaoning.[64][65] By predating Archaeopteryx, Anchornis proves the existence of a modernly feathered theropod ancestor, providing insight into the dinosaur-bird transition. The specimen shows distribution of large pennaceous feathers on the forelimbs and tail, implying that pennaceous feathers spread to the rest of the body at an earlier stage in theropod evolution.[66] The discovery, in 2011, of feathers preserved in amber, within samples dating to 80 mya, suggests the coexistence of theropods and birds, with both theropod and avian feather types commingled in the samples.[67] x
Evolutionary stages
 Diagram illustrating stages of evolution
Diagram illustrating stages of evolutionSeveral studies of feather development in the embryos of modern birds, coupled with the distribution of feather types among various prehistoric bird precursors, have allowed scientists to attempt a reconstruction of the sequence in which feathers first evolved and developed into the types found on modern birds.
Feather evolution was been broken down into the following stages by Xu and Guo in 2009:[61]
- Single filament
- Multiple filaments joined at their base
- Multiple filaments joined at their base to a central filament
- Multiple filaments along the length of a central filament
- Multiple filaments arising from the edge of a membranous structure
- Pennaceous feather with vane of barbs and barbules and central rachis
- Pennaceous feather with an asymmetrical rachis
- Undifferentiated vane with central rachis
However, Foth (2011) showed that some of these purported stages (stages 2 and 5 in particular) are likely simply artifacts of preservation caused by the way fossil feathers are crushed and the feather remains or imprints are preserved. Foth re-interpreted stage 2 feathers as crushed or misidentified feathers of at least stage 3, and stage 5 feathers as crushed stage 6 feathers.[68]
The following simplified diagram of dinosaur relationships follows these results, and shows the likely distribution of plumaceous (downy) and pennaceous (vaned) feathers among dinosaurs and prehistoric birds. The diagram follows one presented by Xu and Guo (2009) modified with the findings of Foth (2011).[61] The numbers accompanying each name refer to the presence of specific feather stages. Note that 's' indicates the known presence of scales on the body.
Dinosauria Ornithischia Thyreophora (s)
Ornithopoda (s)
Psittacosauridae (s, 1)
Ceratopsidae (s)
Saurischia Sauropodomorpha (s)
Aucasaurus (s)
Carnotaurus (s)
Ceratosaurus (s)
Coelurosauria Dilong (3?)
Other tyrannosauroids (s)
Juravenator (s, 3?)
Sinosauropteryx (3+)
Maniraptora Therizinosauria (1, 3+)
Alvarezsauridae (3?)
Oviraptorosauria (4, 6)
Paraves Troodontidae (3+, 6)
Other dromaeosaurids
Sinornithosaurus (3+, 6)
Microraptor (3+, 6, 7)
Scansoriopterygidae (3+, 6, 8)
Archaeopterygidae (3+, 6, 7)
Jeholornis (6, 7)
Confuciusornis (4, 6, 7, 8)
Enantiornithes (4, 6, 7, 8)
Neornithes (4, 6, 7, 8)
See also
References
- ^ a b c d Prum, Richard O. & AH Brush (2002). "The evolutionary origin and diversification of feathers". The Quarterly Review of Biology 77 (3): 261–295. doi:10.1086/341993. PMID 12365352. http://www.yale.edu/eeb/prum/pdf/Prum_n_Brush_2002.pdf. Retrieved 7 July 2010.
- ^ a b c d Prum, R.O., & Brush, A.H (March 2003). "Which Came First, the Feather or the Bird?". Scientific American 288 (3): 84–93. doi:10.1038/scientificamerican0303-84. PMID 12616863. http://www.yale.edu/eeb/prum/pdf/Prum_n_Brush_2003.pdf. Retrieved 7 July 2010
- ^ Prum, Richard O (1999). "Development and Evolutionary Origin of Feathers". Journal of Experimental Zoology (Molecular and Developmental Evolution) 285 (4): 291–306. doi:10.1002/(SICI)1097-010X(19991215)285:4<291::AID-JEZ1>3.0.CO;2-9. PMID 10578107. http://www.ncsce.org/PDF_files/feathers/Prum%20feather.pdf. Retrieved 7 July 2010
- ^ a b c Pettingill, OS Jr. (1970). Ornithology in Laboratory and Field. Fourth edition. Burgess Publishing Company. pp. 29–58. ISBN 808716093.
- ^ R. Schor and S. Krimm (1961). "Studies on the Structure of Feather Keratin II. A β-Helix Model for the Structure of Feather Keratin". Biophys J. 1 (6): 489–515. Bibcode 1961BpJ.....1..489S. doi:10.1016/S0006-3495(61)86904-X. PMC 1366335. PMID 19431311. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1366335.
- ^ Linus Pauling and Robert B. Corey (1951). "The Structure of Feather Rachis keratin". Proceedings of the National Academy of Sciences of the United States of America 37 (5): 256–261. Bibcode 1951PNAS...37..256P. doi:10.1073/pnas.37.5.256. PMC 1063351. PMID 14834148. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1063351.
- ^ Hornik, C., Krishan, K., Yusuf, F., Scaal, M., & Brand-Saberi, B. (2005). "cDermo-1 misexpression induces dense dermis, feathers, and scales". Developmental Biology 277 (1): 42–50. doi:10.1016/j.ydbio.2004.08.050. PMID 15572138.
- ^ McLelland, J. (1991). A color atlas of avian anatomy. W.B. Saunders Co.. ISBN 0721635369.
- ^ Cameron, G., Wess, T., & Bonser, R. (2003). "Young's modulus varies with differential orientation of keratin in feathers". Journal of Structural Biology 143 (2): 118–23. doi:10.1016/S1047-8477(03)00142-4. PMID 12972348.
- ^ Bonser, R., Saker, L., & Jeronimidis, G. (2004). "Toughness anisotropy in feather keratin". Journal of Materials Science 39 (8): 2895–2896. Bibcode 2004JMatS..39.2895B. doi:10.1023/B:JMSC.0000021474.75864.ff.
- ^ Muir D. Eaton and Scott M. Lanyon (2003). "The Ubiquity of Avian Ultraviolet Plumage Reflectance". Proceedings: Biological Sciences 270 (1525): 1721–1726. doi:10.1098/rspb.2003.2431. PMC 1691429. PMID 12965000. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1691429.
- ^ Bostwick, Kimberly S. and Richard O. Prum (2005). "Courting Bird Sings with Stridulating Wing Feathers". Science 309 (5735): 736. doi:10.1126/science.1111701. PMID 16051789. http://cumv.bio.cornell.edu/pdf/Bostwick_Prum_2005_manuscript.pdf. Retrieved 19 July 2010.
- ^ a b Delhey K, A. Peters, and B. Kempenaers (2007). "Cosmetic coloration in birds: occurrence, function and evolution" (PDF). Am. Nat. 169: S145–158. doi:10.1086/510095. PMID 19426089. http://www.orn.mpg.de/documents/peters/Delhey_AmNat2007_copy.pdf.
- ^ Ribak, G., Weihs, D. and Arad, Z. (2005). "Water retention in the plumage of diving great cormorants Phalacrocorax carbo sinensis". J. Avian Biol. 36: 89–95. doi:10.1111/j.0908-8857.2005.03499.x.
- ^ Roger J. Lederer (1972). "The role of avian rictal bristles". Wilson. Bull. 84, 193-97 [pdf]. University of New Mexico. http://elibrary.unm.edu/sora/Wilson/v084n02/p0193-p0197.pdf. Retrieved 16 September 2010.
- ^ Conover, M. R., and D. E. Miller (1980) Rictal bristle function in willow flycatcher. Condor 82:469-471.
- ^ Piersma, T & M R van Eerden (1989). "Feather eating in Great Crested Grebes Podiceps cristatus: a unique solution to the problems of debris and gastric parasites in fish-eating birds". Ibis 131 (4): 477–486. doi:10.1111/j.1474-919X.1989.tb04784.x.
- ^ Demay, Ida S. (1940). "A Study of the Pterylosis and Pneumaticity of the Screamer". The Condor 42 (2): 112–118. doi:10.2307/1364475. JSTOR 1364475.
- ^ K. Susanna S. Hall (2005). "Do nine-primaried passerines have nine or ten primary feathers? The evolution of a concept". Journal of Ornithology 146 (2): 121–126. doi:10.1007/s10336-004-0070-5.
- ^ Pycraft, W. P. (1895). "On the pterylography of the hoatzin (Opisthocomus cristatus)". Ibis 37: 345–373. doi:10.1111/j.1474-919X.1895.tb06744.x.
- ^ McGraw KH & MC Nogare (2005). "Distribution of unique red feather pigments in parrots". Biology Letters 1 (1): 38–43. doi:10.1098/rsbl.2004.0269. PMC 1629064. PMID 17148123. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1629064.
- ^ Hausmann, F., Arnold, K.E., Marshall, N.J. & Owens, I.P.F. (2003). "Ultraviolet signals in birds are special". Proc. R. Soc. B 270 (1510): 61–67. doi:10.1098/rspb.2002.2200. PMC 1691211. PMID 12590772. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1691211.
- ^ Matthew D Shawkey and Geoffrey E Hill (2005). "Carotenoids need structural colours to shine" (PDF). Biol Lett. 1 (2): 121–124. doi:10.1098/rsbl.2004.0289. PMC 1626226. PMID 17148144. http://nature.berkeley.edu/%7Emshawkey/9.pdf.
- ^ Vinther, Jakob; Derek E. G. Briggs; Julia Clarke; Gerald Mayr; Richard O. Prum (2009). "Structural coloration in a fossil feather". Biology Letters 6 (1): 128–31. doi:10.1098/rsbl.2009.0524. PMC 2817243. PMID 19710052. http://www.eeb.yale.edu/prum/pdf/Vinther%20et%20al%202010.pdf. Retrieved 19 July 2010.
- ^ Dyck J. (1971). "Structure and spectral reflectance of green and blue feathers of the Lovebird (Agapornis roseicollis)". Biol. Skr. 18: 1–67.
- ^ Shawkey MD & G E Hill (2005). "Feathers at a fine scale" (PDF). The Auk 121 (3): 652–655. doi:10.1642/0004-8038(2004)121[0652:FAAFS]2.0.CO;2. http://nature.berkeley.edu/%7Emshawkey/6.pdf.
- ^ Delhey, K., A. Peters, PHW Biedermann & B Kempenaers (2008). "Optical properties of the uropygial gland secretion: no evidence for UV cosmetics in birds". Naturwissenschaften 95 (10): 939–46. Bibcode 2008NW.....95..939D. doi:10.1007/s00114-008-0406-8. PMID 18560743.
- ^ Shawkey, M.D., S.R. Pillai, and G.E. Hill (2003). "Chemical warfare? Effects of uropygial oil on feather-degrading bacteria" (PDF). Journal of Avian Biology 34: 345–349. doi:10.1111/j.0908-8857.2003.03193.x. http://nature.berkeley.edu/~mshawkey/2.pdf.
- ^ Bonser, R. H. C. (1995). "Melanin and the abrasion resistance of feathers". Condor 97 (2): 590–591. doi:10.2307/1369048. JSTOR 1369048.
- ^ Grande JM, Negro JJ & MJ Torres (2004). "The evolution of bird plumage colouration: A role for feather-degrading bacteria?" (PDF). Ardeola 51 (2): 375–383. http://www.ardeola.org/files/Ardeola_51(2)_375-383.pdf.
- ^ Burtt, Edward H. Jr. & Ichida, Jann M. (2004). "Gloger's Rule, feather-degrading bacteria, and color variation among Song Sparrows" (PDF). Condor 106 (3): 681–686. doi:10.1650/7383. http://www.public.asu.edu/~kjmcgraw/pubs/Condor04b.pdf.
- ^ Endler, J. A. (1980). "Natural selection on color patterns in Poeci-lia reticulata". Evolution 34 (1): 76–91. doi:10.2307/2408316. JSTOR 2408316.
- ^ Badyaev AV & Hill GE (2000). "Evolution of sexual dichromatism: contribution of carotenoid versus melanin-based colouration". Biological Journal of the Linnean Society 69: 153–172. doi:10.1111/j.1095-8312.2000.tb01196.x.
- ^ Lozano, G. A. 1994. Carotenoids, parasites, and sexual selection. Oikos 70: 309-311
- ^ Toon, A., & Hughes, J. (2008). "Are lice good proxies for host history? A comparative analysis of the Australian magpie, Gymnorhina tibicen, and two species of feather louse". Heredity 101 (2): 127–135. doi:10.1038/hdy.2008.37. PMID 18461081.
- ^ Brooke, M. de L. and Hiroshi Nakamura (1998). "The acquisition of host-specific feather lice by common cuckoos (Cuculus canorus)". Journal of Zoology 244: 167–173. doi:10.1017/S0952836998002027.
- ^ Revis, Hannah C., and Deborah A. Waller (2004). "Bactericidal and fungicidal activity of ant chemicals on feather parasites: an evaluation of anting behavior as a method of self-medication in songbirds". Auk 121 (4): 1262–1268. doi:10.1642/0004-8038(2004)121[1262:BAFAOA]2.0.CO;2.
- ^ Bonser, R.H.C. & Dawson, C. (1999). "The structural mechanical properties of down feathers and biomimicking natural insulation materials". Journal of Materials Science Letter 18 (21): 1769–1770. doi:10.1023/A:1006631328233.
- ^ "3Pen". The Merriam-Webster Online Dictionary. Merriam-Webster, Inc. http://www.merriam-webster.com/dictionary/pen%5B3%5D. Retrieved 16 October 2010.
- ^ Bowen, Gabriel J; Wassenaar, Leonard I; Hobson, Keith A (2005). "Global application of stable hydrogen and oxygen isotopes to wildlife forensics". Oecologia 143 (3): 337–348. doi:10.1007/s00442-004-1813-y. PMID 15726429.
- ^ Jaspers, V., Voorspoels, S., Covaci, A., Lepoint, G., & Eens, M. (2007). "Evaluation of the usefulness of bird feathers as a non-destructive biomonitoring tool for organic pollutants: A comparative and meta-analytical approach". Environment International 33 (3): 328–337. doi:10.1016/j.envint.2006.11.011. PMID 17198730.
- ^ Subbiah Poopathi, S. Abidha (2007). "Use of feather-based culture media for the production of mosquitocidal bacteria". Biological Control 43 (1): 49–55. doi:10.1016/j.biocontrol.2007.04.019.
- ^ Schmidt, W.F., Barone, J.R. (2004). New uses for chicken feathers keratin fiber. Poultry Waste Management Symposium Proceedings. pp. 99–101.
- ^ Casarin, Franciani; Brandelli, Florencia Cladera-Olivera Adriano (2008). "Use of Poultry Byproduct for Production of Keratinolytic Enzymes". Food and Bioprocess Technology 1 (3): 301–305. doi:10.1007/s11947-008-0091-9.
- ^ Jiang, Z., Qin, D., Hse, C., Kuo, M., Luo, Z., Wang, G., et al. (2008). "Preliminary Study on Chicken Feather Protein-Based Wood Adhesives". Journal of Wood Chemistry & Technology 28 (3): 240–246. doi:10.1080/02773810802347073.
- ^ Steve Froemming (2006). "Traditional use of the Andean flicker (Colaptes rupicola) as a galactagogue in the Peruvian Andes". Journal of Ethnobiology and Ethnomedicine 2: 23. doi:10.1186/1746-4269-2-23. PMC 1484469. PMID 16677398. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1484469.
- ^ Murari, S.K., Frey, F.J., Frey, B.M., Gowda, T.V., Vishwanath, B.S. (2005). "Use of Pavo cristatus feather extract for the better management of snakebites: Neutralization of inflammatory reactions". Journal of Ethnopharmacology 99 (2): 229–237. doi:10.1016/j.jep.2005.02.027. PMID 15894132.
- ^ Mahawar, MM & DP Jaroli (2007). "Traditional knowledge on zootherapeutic uses by the Saharia tribe of Rajasthan, India". J Ethnobiol Ethnomedicine 3: 25. doi:10.1186/1746-4269-3-25. PMC 1892771. PMID 17547781. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1892771.
- ^ Doughty, Robin W. 197. Feather Fashions and Bird Preservation, A Study in Nature Protection. University of California Press.
- ^ Ehrlich, Paul R.; Dobkin. David S.; Wheye. Darryl (1988) Plume Trade Stanford University
- ^ Feather trade Smithsonian Institution
- ^ http://seattletimes.nwsource.com/html/outdoors/2015246435_flyfash.html
- ^ Sumida, SS & CA Brochu (2000). "Phylogenetic context for the origin of feathers". American Zoologist 40 (4): 486–503. doi:10.1093/icb/40.4.486. http://icb.oxfordjournals.org/cgi/content/abstract/40/4/486.
- ^ Bock, WJ (2000). "Explanatory History of the Origin of Feathers". Amer. Zool. 40 (4): 478–485. doi:10.1093/icb/40.4.478.
- ^ De Ricqles, A. J., K. Padian, J. R. Horner, E. T. Lamm, and N. Myhrvold (2003). "Osteohistology of confuciusornis sanctus (theropoda: Aves)". Journal of Vertebrate Paleontology 23: 753-761.
- ^ Saino, Nicola, and Riccardo Stradi (1999). "Carotenoid Plasma Concentration, Immune Profile, and Plumage Ornamentation of Male Barn Swallows". American Naturalist 154 (4): 441–448. doi:10.1086/303246. PMID 10523490.
- ^ Endler, John A., David A. Westcott, Joah R. Madden, Tim Robson, and Patrick Phillips (2005). "Animal visual systems and the evolution of color patterns: Sensory processing illumiates signal evolution". Evolution 59 (8): 1795–1818. PMID 16329248.
- ^ Xu, Xing & Fucheng Zhang (2005). "A new maniraptoran dinosaur from China with long feathers on the metatarsus". Naturwissenschaften 92 (4): 173–177. Bibcode 2005NW.....92..173X. doi:10.1007/s00114-004-0604-y. PMID 15685441.
- ^ Xu, Xing (2006). "Feathered dinosaurs from China and the evolution of major avian characters". Integrative Zoology 1 (1): 4–11. doi:10.1111/j.1749-4877.2006.00004.x.
- ^ Zheng,X. T.,H. L. You,X. Xu and Z. M. Dong (2009). "An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures". Nature 458: 333-336. doi:10.1038/nature07856.
- ^ a b c Xu, X.; Guo, Y. (2009). "The origin and early evolution of feathers: insights from recent paleontological and neontological data". Vertebrata PalAsiatica 47 (4): 311–329.
- ^ Ji, Q., P. J. Currie, M. A. Norell, and S. A. Ji (1998). "Two feathered dinosaurs from northeastern China". Nature 393 (6687): 753–761. Bibcode 1998Natur.393..753Q. doi:10.1038/31635.
- ^ Xu, X., H. H. Zhou, and R. O. Prum (2001). "Branched integumental structures in Sinornithosaurus and the origin of feathers". Nature 410 (6825): 200–204. doi:10.1038/35065589. PMID 11242078.
- ^ Hu, D. Y., L. H. Hou, L. J. Zhang, and X. Xu (2009). "A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus". Nature 461 (7264): 640–643. Bibcode 2009Natur.461..640H. doi:10.1038/nature08322. PMID 19794491.
- ^ Xu, X., Q. Zhao, M. Norell, C. Sullivan, D. Hone, G. Erickson, X. L. Wang et al. (2009). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin". Chinese Science Bulletin 54: 430–435. doi:10.1007/s11434-009-0009-6.
- ^ Witmer, L. M. (2009). "Feathered dinosaurs in a tangle". Nature 461 (7264): 601–602. Bibcode 2009Natur.461..601W. doi:10.1038/461601a. PMID 19794481.
- ^ [1]
- ^ Foth, C. (2011). "On the identification of feather structures in stem-line representatives of birds: evidence from fossils and actuopalaeontology." Paläontologische Zeitschrift, (advance publication) doi:10.1007/s12542-011-0111-3
External links
- McGraw, K. J. 2005. Polly want a pigment? Cracking the chemical code to red coloration in parrots. Australian Birdkeeper Magazine 18:608-611.
- DeMeo, Antonia M. Access to Eagles and Eagle Parts: Environmental Protection v. Native American Free Exercise of Religion (1995)
- Electronic Code of Federal Regulations (e-CFR), Title 50: Wildlife and Fisheries PART 22—EAGLE PERMITS
- U.S. v. Thirty Eight Golden Eagles (1986)
- Mechanical structure of feathers
- Documentary on the evolution of feathers
- Lecture notes on the avian integument
- U.S. National Fish and Wildlife Forensics Laboratory's Feather Atlas
- Federn.org
Categories:- Feathers
- Bird products
- Bird anatomy













Wikimedia Foundation. 2010.