- Confuciusornis
-
Confuciusornis
Temporal range: Early Cretaceous, 125–120 Ma
Specimen of C. sanctus with paired tail feathers Scientific classification 
Kingdom: Animalia Phylum: Chordata Class: Aves Order: †Confuciusornithiformes Family: †Confuciusornithidae Genus: †Confuciusornis
Hou et al., 1995Type species Confuciusornis sanctus
Hou et al., 1995Species - †C. sanctus Hou et al., 1995
- †C. dui Hou et al., 1999
- †C. feducciai Zhang et al., 2009
- †C. jianchangensis Li, Wang & Hou, 2010
Synonyms - Jinzhouornis? Hou et al., 2002
Confuciusornis is a genus of primitive crow-sized birds from the Early Cretaceous Yixian and Jiufotang Formations of China, dating from 125 to 120 million years ago. Like modern birds, Confuciusornis had a toothless beak, but close relatives of modern birds such as Hesperornis and Ichthyornis were toothed, indicating that the loss of teeth occurred convergently in Confuciusornis and living birds. It is the oldest known bird to have a beak.[1] It was named after the Chinese moral philosopher Confucius (551–479 BCE). Confuciusornis is one of the most abundant vertebrates found in the Yixian Formation, and several hundred complete, articulated specimens have been found.[2]
Contents
Discovery and naming
In November 1993 Chinese paleontologists Zhou Zhonge, Hu Yoaming and Hou Lianhai, of the Institute of Vertebrate Paleontology and Paleoanthropology at Beijing, were presented a bird fossil bought at a flea market in Sihetun by amateur paleontologist Zhang He. It showed a partial skeleton of a bird new to science of which even some feather remains had been preserved. In December 1993 two further specimens were acquired from a farmer, Yang Yushan. Soon afterwards, it was discovered that local farmers were in the process of collecting hundreds of specimens that were provisionally prepared by them to be illegally sold to commercial fossil dealers.[3] Many hundreds have since also been added to the official collections of Chinese institutions. In 2010 the Shandong Tianyu Museum of Nature alone possessed 536 specimens.[4] The fossils are compressed but otherwise typically very complete often showing the entire feathering of the animal.
Based on the abundant material, six species have been formally named and described: C. sanctus (the type species), C. dui, C. feducciai, C. jianchangensis, C. chuonzhous and C. suniae. The latter two are usually considered synonymous with C. sanctus.[5] Most species lived in the early Aptian 125 million year old Jianshangou Beds of the Yixian Formation, though C. jianchangnsis is found in the later (120Ma) middle Aptian Jiufotang Formation. C. sanctus is known from the former, and is one of the most common vertebrate species found in the Yixian, often present in dense concentrations. At one time forty were discovered on a surface of about 100 m². This has been explained as the result of entire flocks of birds being simultaneously killed by ash, heat or poisonous gas following the volcanic eruptions that caused the tuff stone in which the fossils were found to be deposited as lake sediments.[6]
In 1995 Zhou, Hou, Zhang and Gu Youcai named Confuciusornis sanctus. The generic name combines the philosopher Confucius with a Greek ὄρνις, (ornis), "bird". The specific name means "holy one" in Latin and is a translation of Chinese 圣贤, shèngxián, "sage", again in reference to Confucius. The holotype specimen is IVPP V10918.[7] In 1997 Hou named two species. The first was Confuciusornis chuonzhous, based on specimen IVPP V10919, originally a paratype of C. sanctus. The specific name refers to Chuanzhou, an ancient name for Beipiao. The second species was Confuciusornis suniae, based on specimen IVPP V11308. The specific name honours madam Sun, the wife of Shikuan Liang who donated the fossil to the IVPP.[8] In 1999 Hou, Zhou, Zhang, Larry Martin and Alan Feduccia named Confuciusornis dui, based on specimen IVPP V11553. The specific name again honours a donating collector: Du Wengya.[9] In 2009 Zhang Fucheng et al. named Confuciusornis feducciai, based on specimen D2454, the specific name honouring Feduccia.[10] In 2010 Li Li, Wang Jingqi and Hou Shilin named Confuciusornis jianchangensis, based on specimen PMOL-AB00114 found at Toudaoyingzi.[11]
In 2002 Hou named the genus Jinzhouornis. Luis Chiappe later concluded that this is probably a junior synonym of Confuciusornis.
Description
 Size of Confuciusornis, compared to a human.
Size of Confuciusornis, compared to a human.
 C. sanctus fossil preserving long wing and tail feathers
C. sanctus fossil preserving long wing and tail feathersConfuciusornis was about the size of a modern pigeon, with a wingspan of up to 0.7 meters (2.3 ft), and its body weight has been estimated to have been as much as 1.5 kilograms,[12] or less than 0.2 kilograms.[13] C. feducciai was about a third longer than average specimens of C. sanctus.[10]
Confuciusornis shows a mix of basal and derived traits. It was more "advanced" or derived than Archaeopteryx in possessing a short tail with a pygostyle (a bone formed from a series of short, fused tail vertebrae) and a bony sternum, but more basal or "primitive" than modern birds in retaining large claws on the forelimbs, having a primitive skull with a closed eye-socket, and a relatively small breastbone. At first the number of basal characteristics was exaggerated: Hou assumed in 1995 that a long tail was present and mistook grooves in the jaw bones for small degenerated teeth.[14]
The skull of Confuciusornis was equipped with a pointed toothless beak. It was relatively heavy-built and immobile, incapable of the kinesis of modern birds that can raise the snout relative to the back of the skull. This immobility was caused by the presence of a triradiate postorbital separating the eye-socket from the lower temporal opening, like with more basal theropods, and the premaxillae of the snout reaching all the way to the frontals, forcing the nasals to the sides of the snout.[15]
Fossils of Confuciusornis show that it had an exceptionally large humerus (upper arm bone). Near its shoulder-end this was equipped with a prominent deltopectoral crest. Characteristically this crista deltopectoralis was with Confuciusornis pierced by an oval hole which may have reduced the bone's weight or enlarged the attachment area of the flight miscles. The furcula or wishbone, like that of Archaeopteryx, was a simple curved bar lacking a pointed process at the back, a hypocleidum. The sternum (breastbone) was relatively broad and had a low keel which was raised at the back end. This bony keel may or may not have anchored a larger, cartilaginous, keel for enlarged pectoral muscles.[15] The scapulae (shoulder blades) were fused to the strut-like coracoid bones and may have formed a solid base for the attachment of wing muscles. The orientation of the shoulder joint was sideways, instead of angled upward as in modern birds; this means that Confuciusornis was unable to lift its wings high above its back. According to a study by Phil Senter in 2006, the joint was even pointed largely downwards meaning that the humerus could not be lifted above the horizontal. This would make Confuciusornis incapable of the upstroke required for flapping flight; the same would have been true for Archaeopteryx.[16]
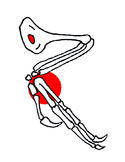 Illustration of Confuciusornis wing bones.
Illustration of Confuciusornis wing bones.The wrist of Confuciusornis shows fusion, forming a carpometacarpus. The second and third metacarpals were also partially fused but the first was unfused, however, and also the fingers could freely move relative to each other. The second metacarpal, supporting the flight feathers, was very heavily built; its finger carries a small claw. The claw of the first finger to the contrary was very large and curved, that of the third intermediate in size.[15] The formula of the finger phalanges was 2-3-4-0-0.
The pelvis was connected to a sacrum formed by seven sacral vertebrae. The pubis was strongly pointing backwards. The left and right ischia were not fused. The femur was straight; the tibia only slightly longer. The metatarsals of the foot were relatively short and fused to each other and to the lower ankle bones, forming a tarsometatarsus. A rudimentary fifth metatarsal is present. The first metatarsal was attached to the lower shaft of the second and supported a first toe or hallux, pointing to the back.[15] The formula of the toe phalanges was 2-3-4-5-0. The proportions of the toes suggest that they were used for both walking and perching, while the large claws of the thumb and third finger were probably used for climbing.
Feathers
 Life restoration of C. sanctus
Life restoration of C. sanctusThe wing feathers of Confuciusornis were long and modern in appearance. The primary wing feathers of a 0.5 kilogram individual reached 20.7 centimeters in length. The five longest primary feathers (remiges primarii) were more than 3.5 times the length of the hand and relatively longer than those of any living bird, while the secondary feathers of the lower arm were rather short by comparison. Thus, the wing shape was very unlike that of living birds, being long and narrow.[12] The primary feathers were asymmetrical to varying degree, and especially so in the outermost primaries.[15] It is unclear whether the upper arm carried tertiaries. Covert feathers are preserved covering the upper part of the wing feathers in some specimens, and some specimens have preserved the contour feathers of the body.[15] Unlike some more advanced birds, Confuciusornis lacked an alula, or "bastard wing". In modern birds this is formed by feathers anchored to the first digit of the hand, but this digit appears to have been free of feathers and independent of the body of the wing in Confuciusornis.[15] According to Dieter Stefan Peters to compensate for the lack of an alula, the third finger might have formed a separate winglet below the main wing, functioning like the flap of an aircraft.[17] Despite the relatively advanced and long wing feathers, the forearm bones lacked any indication of quill knobs (papillae ulnares), or bony attachment points for the feather ligaments.[15]
Many specimens preserve a pair of long, narrow tail feathers, which grew longer than the entire length of the rest of the body. Unlike the feathers of most modern birds, these feathers were not differentiated into a central quill and barbs for most of their length. Rather, most of the feather formed a ribbon-like sheet, about six millimetres wide. Only at the last one quarter of the feather, towards the rounded tip, does the feather become differentiated into a central shaft with interlocking barbs. Many individuals of Confuciusornis lacked even these two tail feathers, possibly due to sexual dimorphism. The rest of the tail around the pygostyle was covered in short, non-aerodynamic feather tufts similar to the contour feathers of the body, rather than the familiar feather fan of modern bird tails.[15]
Colouration
In early 2010, a group of scientists led by Zhang Fucheng examined fossils with preserved melanosomes (organelles which contain colors). By studying such fossils with an electron microscope, they found melanosomes preserved in a fossil Confuciusornis specimen, IVPP V13171.[18] The melanosomes where of two types, eumelanosomes and pheomelanosomes. This indicates that Confuciusornis had hues of grey, red/brown and black, possibly something like the modern zebra finch. It was also the first time an early bird fossil has been shown to contain preserved pheomelanosomes.[19][20]
Paleobiology
Flight
 Short tailed C. sanctus specimens in CosmoCaixa Barcelona
Short tailed C. sanctus specimens in CosmoCaixa BarcelonaConfuciusornis has traditionally been assumed to have been a competent flier based on its extremely long wings with strongly asymmetrical feathers. Other adaptations for improved flight capabilities include: a fused wrist, a short tail, an ossified sternum, a strut-like coracoid, a large deltopectoral crest, a strong ulna and an enlarged second metacarpal.[21]
However, over the years several claims have been made that the flight of Confuciusornis suffered from more or less severe limitations.
The first of these regarded problems to attain a steep flight path due to a limited wing amplitude. In the interpretation of Senter (2006) of the position of the shoulder joint, a normal upstroke would be impossible precluding flapping flight entirely. Less radical is the assessment that due to the lack of a keeled sternum and a high acrocoracoid, the Musculus pectoralis minor could not serve as a M. supracoracoideus lifting the humerus via a tendon running through a foramen triosseum. This, coupled with a limited upstroke caused by a lateral position of the shoulder joint, would have made it difficult to gain altitude. Some authors therefore proposed that Confuciusornis used its large thumb claws to climb tree trunks. Martin assumed that it could raise its torso almost vertically like a squirrel.[14] Daniel Hembree however, while acknowledging that tree climbing was likely, pointed out that the rump was apparently not lifted more than 25° relative to the femur in vertical position, as shown by the location of the antitrochanter in the hip joint.[22] Dieter S. Peters considered it very unlikely that Confuciusornis climbed trunks as turning the thumb claw inwards would stretch the very long wing forwards, right in the path of obstructing branches. Peters sees Confuciusornis as capable of flapping flight but specialised in soaring flight.[17]
A second problem is the strength of the feathers. In 2010 Robert Nudds and Gareth Dyke published a study arguing that in both Confuciusornis and Archaeopteryx, the raches (central shafts) of the primary feathers were too thin and weak to have remained rigid during the power stroke required for true flight. They argued that Confuciusornis would at most have employed gliding flight, which is also consistent with the unusual adaptations seen in its upper arm bones, and more likely used its wings for mere parachuting, limiting fall speed if it dropped from a tree.[12] Gregory S. Paul, however, disagreed with their study. He argued that Nudds and Dyke had overestimated the weights of these early birds, and that more accurate weight estimates allowed powered flight even with relatively narrow raches. Nudds and Dyke assumed a weight of 1.5 kilograms for Confuciusornis, as heavy as the modern teal. Paul argued that a more reasonable body weight estimate is about 180 grams, less than that of a pigeon. Paul also noted that Confuciusornis is commonly found as large assemblages in lake bottom sediments with little to no evidence of extensive postmortem transport, and that it would be highly unusual for gliding animals to be found in such large numbers in deep water. Rather, this evidence suggests that Confuciusornis traveled in large flocks over the lake surfaces, a habitat consistent with a flying animal.[13] A number of Chinese researchers questioned the correctness of the rachis measurements, stating that the specimens they had studied showed a shaft thickness of 2.1 to 2.3 millimetres as compared to the 1.2 millimetre reported by Nudds and Dyke.[4] The latter replied that, apart from the weight aspect, such greater shaft thickness alone would make flapping flight possible; however, they allowed for the possibility of two species being present in the Chinese fossil material with a differing rachis diameter.[23]
Sexual variation
Many specimens of Confuciusornis preserve a single pair of long, streamer-like tail feathers. However, most do not, even specimens which otherwise have exquisitely preserved feathers on the rest of the body; their predominance has been estimated to be as high as 90-95%.[14] This difference has been suggested to indicate sexual dimorphism, with one gender (likely the males) using the streamer-like feathers in courtship displays. However, while sexual variation is the most obvious explanation for the presence or absence of long tail feathers, other factors cannot be ruled out. For example, it is possible that some individuals lack tail feathers because they were fossilized during molting. As in modern birds, molting individuals may have been present alongside non-molting individuals, and males and females may have molted at different times during the year.[15]
Due to the large number of specimens, statistical analysis can be used to investigate this problem. In 2008 Chiappe et alii published a morphometrical study of 106 fossils. The population showed a clear bimodal distribution of the size of the animals with two weight classes. However, there was no correlation between size and the possession of the long tail feathers. From this it was concluded that either the sexes did not differ in size or both sexes had the long feathers. The first case was deemed most likely which left the size distribution to be explained. It was hypothesised that the smaller animals consisted of very young individuals, that the large animals were adults and that the rarity of individuals with an intermediate size was caused by Confuciusornis experiencing a growth spurt just prior to reaching adulthood, the shortness of which would have prevented many becoming fossilised during this phase.[24] This interpretation was criticised by Dieter S. Peters who pointed out that, as the smaller animals were not neonates, the young of Confuciusornis had to grow very fast immediately after hatching regardless. This would then imply two growth spurts, which assumption was not very parsimonious. He thought the data would be more simply explained by assuming that both sexes had the long feathers and one of them was the largest. He favoured the possibility that these were the females, having observed such a pattern with the modern Pheasant-tailed Jacana Hydrophasianus chirurgus, a water-bird in which both sexes have long tails and the female sex is the heaviest. He accounted for the fact that most specimens show no tails at all by the possibility that Confuciusornis shed the feathers as a defence mechanism, a method used by several extant species, which would have been triggered by the stress induced by the very volcanic explosion that buried the animals.[25]
The growth speed relevant to this question can be independently measured by researching the bone structure. In 1998 Zhang determined that growth was fast, leading to fibrolamellar bone.[26] Comparison with the bone structure of an alligator indicated that Confuciusornis had a high metabolism comparable to that of modern birds.[27] The growth speed of Confuciusornis would have been similar to that of Beipiaosaurus.[28] A study by Armand de Ricqlès in 2003 indicated that growth was essentially modern, Confuciusornis reaching its maximum size in at most twenty weeks, perhaps as few as thirteen. Basal birds would have limited their size, not by slowing growth but by limiting the period of growth.[29]
Diet
There are immature specimens known, and from the analysis of bone growth patterns of young adults it has been estimated that Confuciusornis reached maturity somewhat slower than extant small birds, but faster than advanced dinosaurs (de Ricqlès et al., 2003), which might indicate an omnivorous diet similar to modern crows.
It has been hypothesized that Confuciusornis fed on plant materials due to its toothless beak, but no gastroliths or stomach contents had been reported (Zhou & Zhang, 2003). Dalsätt and colleagues (2006) described a specimen, IVPP V13313 found in the Jiufotang Beds, that preserves seven to nine vertebrae and several ribs of a small fish, probably Jinanichthys. These fish bones are formed into a tight cluster about six millimeters across, and the cluster is in contact with the seventh and eighth cervical vertebrae of the bird. In this position it was likely in the crop of the bird, which may have been preparing to regurgitate a pellet when it died. No other fish remains are present in the slab. (Dalsätt et al., 2006). Andrzej Elzanowski already in 2002 had predicted that Confuciusornis was a predator.[30] Dieter S. Peters has hypothesized that, although no remains of toe webs have been conserved, it caught its prey swimming using its rather soft bill to search for prey below the waterline.[17] Andrei Zinoviev assumed it caught fish on the wing.[31][32]
Several extant bird species have been presented as modern analogues of Confuciusornis providing insight into its possible lifestyle. Dieter S. Peters thought that it could be best compared with the White-tailed Tropicbird Phaeton lepturus, a fisher that too has a long tail and narrow wings — and even often nests in the neighbourhood of volcanoes.[17] The paradise kingfishers (genus Tanysiptera) of modern Australia and New Guinea have elongated "racket plumes" as their central tail feathers, giving them a superficial resemblance to Confuciusornis. These kingfishers also take fish as prey, but they are not specialized fishers. They take many insects and other small prey from their forest habitat.[33]
Daily activity patterns
Comparisons between the scleral rings supporting the eyes of Confuciusornis and modern birds and other reptiles indicate that it may have been diurnal, similar to most modern birds.[34]
Classification
Hou assigned Confuciusornis to the Confuciusornithidae in 1995. At first he assumed it was a member of the Enantiornithes and the sister taxon of Gobipteryx. Later he understood that Confuciusornis was not a enantiornith but concluded it was the sister taxon of the Enantiornithes, within a larger Sauriurae.[35] This was heavily criticised by Chiappe who regarded Sauriurae to be paraphyletic as there were insufficient shared traits that indicated that the Confuciusornithidae and the Enantiornithes were closely related.[36] In 2001 Ji Qiang suggested an alternative position as the sister taxon of the Ornithothoraces.[37]
In 2002 Ji's hypothesis was confirmed by a cladistic analysis by Chiappe, who defined a new group: the Pygostylia of which Confuciusornis is by definition the most basal member.[38] Several traits of Confuciusornis illustrate its position in the tree of life; it has a more "primitive" skull than Archaeopteryx, but it is the first known bird to have lost the long tail of Archaeopteryx and develop fused tail vertebrae, a pygostyle.[39] One controversial study concluded that Confuciusornis may be more closely related to Microraptor and other dromaeosaurids than to Archaeopteryx, but this study was criticized on methodological grounds (Mayr et al., 2005).
The present standard interpretation of the phylogenetic position of Confuciusornis can be shown in this cladogram:
Aves Pygostylia Confuciusornithidae Confuciusornis
Ornithothoraces Ornithuromorpha (including the Neornithes)
A close relative, the confuciusornithid Changchengornis hengdaoziensis, also lived in the Yixian Formation. Changchengornis also possessed the paired, long tail feathers, as did several more advanced enantiornith birds. True, mobile tail fans only appeared in ornithuromorph birds, and possibly in the enantiornithine Shanweiniao.[40][41]
Footnotes
- ^ Ivanov, M., Hrdlickova, S. & Gregorova, R. (2001) The Complete Encyclopedia of Fossils. Rebo Publishers, Netherlands. pp. 312
- ^ Xu, X. and Norell, M.A. (2006). "Non-Avian dinosaur fossils from the Lower Cretaceous Jehol Group of western Liaoning, China."Geological Journal, 41: 419–437.
- ^ Zhou Z., 1995, "The discovery of Early Cretaceous birds in China", Acta Palaeornithologica 181: 9-22
- ^ a b Xiaoting Zheng, Xing Xu, Zhonghe Zhou, Desui Miao, and Fucheng Zhang, 2010, "Comment on “Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability”", Science 15 October 2010: 320
- ^ Li, D., Sulliven, C., Zhou, Z. and Zhang, Z. (2010). "Basal birds from China: a brief review." Chinese Birds, 1(2): 83–96 doi:10.5122/cbirds.2010.0002
- ^ Wang X., Zhang F,, Xu X., Wang Y., Gu G., 2000, "Taphonomy and mass mortality of Confuciusornis and feathered dinosaurs at the Sihetun and Zhangjiagou sites in western Liaoning, China", Vertebrata PalAsiatica, 38(supp): 32
- ^ Hou L., Zhou Z., Gu Y., Zhang H., 1995, "Confuciusornis sanctus, a new Late Jurassic sauriurine bird from China", Chinese Science Bulletin, 40: 1545–1551
- ^ Hou L., 1997, [Chinees] [Mesozoic Birds of China. Phoenix Valley Provincial Aviary of Taiwan]. Taiwan
- ^ Hou L.; Martin, L.D., Zhou Z., Feduccia, A., Zhang F., 1999, "A diapsid skull in a new species of the primitive bird Confuciusornis, Nature 399: 679–682
- ^ a b Zhang Z., Gao C., Meng Q., Liu J., Hou L., Zheng G., 2009, "Diversification in an Early Cretaceous avian genus: evidence from a new species of Confuciusornis from China", Journal of Ornithology, 150: 783–790
- ^ L. Li, J.-q. Wang, and S.-l. Hou. 2010. [Chinese] [A new species of Confuciusornis from Lower Cretaceous of Jianchung, Liaoning, China]. Global Geology 29(2): 183-187
- ^ a b c Nudds, R.L. and Dyke, G.J (2010). "Narrow primary feather rachises in Confuciusornis and Archaeopteryx suggest poor flight ability." Science, 328(5980): 887. doi:10.1126/science.1188895 PMID 20466930
- ^ a b Paul, G.S. (2010). "Comment on 'Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability.'" Science, 330(6002): 320. (15 October 2010). doi:10.1126/science.1192963
- ^ a b c Martin, L.D., Zhou, Z., Hou, L. & Feduccia, A., 1998, "Confuciusornis sanctus compared to Archaeopteryx lithographica", Naturwissenschaften 85: 286–289
- ^ a b c d e f g h i j Chiappe, Luis M., Shu-An, Ji, Qiang, Ji, Norell, Mark A. (1999) "Anatomy and systematics of the Confuciusornithidae (Theropoda:Aves) from the Late Mesozoic of northeastern China" "Bulletin of the American museum of Natural History no.242 89pp.
- ^ Senter, P. (2006). "Scapular orientation in theropods and basal birds, and the origin of flapping flight." Acta Palaeontologica Polonica, 51(2): 305–313.PDF fulltext
- ^ a b c d D.S. Peters & Q. Ji, 1999, "Muβte Confuciusornis klettern?", J. Ornithol. 140(1): 41–50
- ^ Zhang, F., X. Xu, M.J. Benton, Stuart L. Kearns et al. (2010). "Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds", Nature, 463: 1075–1078. See the article online.
- ^ e!Science News: The color of dinosaur feathers discovered.
- ^ ScienceNOW: The Lost World, Now in Colors by Sverker Lundin. January, 27 2010.
- ^ Zhou Z. and Farlow, J.O., 2001, "Flight capability and habits of Confuciusornis". In: Gauthier and Gall (eds). New perspectives on the origin and early evolution of birds: proceedings of the international symposium in honor of John H. Ostrom. Peabody Museum of Natural History. Yale University, New Haven. pp. 237-254
- ^ Hembree, D., 1999, "Re-evaluation of the posture and claws of Confuciusornis", Journal of Vertebrate Paleontology 19: 50A
- ^ Nudds, R.L. & Dyke G., 2010, Response to comments on “Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability”", Science 15 October 2010: 320
- ^ Chiappe, L.M.; Marugan-Lobon, J.; Ji S. and Zhou Z., 2008, "Life history of a basal bird: morphometrics of the Early Cretaceous Confuciusornis", Biology Letters 4(6): 719-723
- ^ Winfried S. Peters and Dieter Stefan Peters, 2009, "Life history, sexual dimorphism and ‘ornamental’ feathers in the mesozoic bird Confuciusornis sanctus", Biology Letters, 5: 817-820
- ^ Zhang, F., L. Hou, and L. Ouyang, 1998, "Osteological microstructure of Confuciusornis: preliminary report", Vertebrata PalAsiatica 36: 126–135
- ^ Zhang, F.-C., Xu X. & Lü J., 1999, "Some microstructure difference among Confuciusornis, Alligator and a small theropod dinosaur, and its implications", Paleoworld, December 1999: 296–308
- ^ Zhang, F.-C., et al., 2000, "Microstructures of Confuciusornis and Beipiaosaurus and their physiological implications", Vertebrata Pal-Asiatica 38 (suppl. Abstracts 5th Society of Avian Paleontology and Evolution Meeting and Jehol Biota Symposium, Beijing 2000): 36
- ^ De Ricqlès, A.J., Padian, K., Horner, J.R., Alamm, E.T. & Myhrvold, N., 2003, "Osteohistology of Confuciusornis sanctus (Theropoda: Aves)", Journal of Vertebrate Paleontology, 23: 373–386
- ^ Elzanowski, A., 2002, "Biology of basal birds and the origin of avian flight". In: Zhou Z., Zhang F. (eds) Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution, Beijing, 1–4 June 2000. Science, Beijing, pp 211–226
- ^ Zinoviev, A., 2009, "ОПЫТ РЕКОНСТРУКЦИИ ЭКОЛОГИЧЕСКОГО ОБЛИКА КОНФУЦИУСОРНИТИД (AVES, CONFUCIUSORNITHIFORMES, CONFUCIUSORNITHIDAE)", Палеонтологический журнал, 4: 83-91
- ^ Zinoviev, A.V., 2009, "An attempt to reconstruct the lifestyle of confuciusornithids (Aves, Confuciusornithiformes)", Paleontological Journal, (43)3: 444-452
- ^ Bleiwess, Robert "Development and evolution of avian racket plumes: Fine structure and serial homology of the wire" Journal of Morphology 194:1 pp.23–39 DOI 5370610.1002/jmor.1051940103
- ^ Schmitz, L.; Motani, R. (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science in press. Bibcode 2011Sci...332..705S. doi:10.1126/science.1200043. PMID 21493820.
- ^ Zhou Z, and Hou L., 1998, "Confuciusornis and the early evolution of birds", Vertebrata PalAsiatica 36(2): 136-146
- ^ Chiappe, L., 1997, "The Chinese early bird Confuciusornis and the paraphyletic status of <<Sauriurae>>", Journal of Vertebrate Paleontology, 17(3): 37A
- ^ Ji Q., 2001, [Chinees] "New advances in the study of the primitive bird Confuciusornis, Geological Science and Technology Information 20: 30-34
- ^ L. M. Chiappe, 2002, "Basal bird phylogeny: problems and solutions". In: L. M. Chiappe and L. M. Witmer (eds.), Mesozoic Birds: Above the Heads of Dinosaurs. University of California Press, Berkeley pp. 448-472
- ^ Clarke,,Julia. A. , Norell, Mark. A. (2002) "The Morphology and Phylogenetic Position of Apsaravis ukhaana from the Late Cretaceous of Mongolia" American Museum Novitates, No. 3387, American Museum of Natural History, New York, NY 10024.
- ^ Clarke, Julia A., Zhou, Zhonghe, Zhang, Fucheng. (2006) "Insight into the evolution of avian flight from a new clade of Early Cretaceous ornithurines from China and the morphology of Yixianornis grabaui" "Journal of Anatomy" 208:287–308.
- ^ Jingmai K. O’connor, Xuri Wang, Luis M. Chiappe, Chunling Gao, Qingjin Meng,Xiaodong Cheng, And Jinyuan Liu (2009). "Phylogenetic support for a specialized clade of Cretaceous enantiornithine birds with information from a new species" Journal of Vertebrate Paleontology 29(1):188–204, March 2009# 2009 by the Society of Vertebrate Paleontology
References
- Dalsätt, J; Zhou, Z; Zhang, F. & Ericson, Per G. P. (2006). Food remains in Confuciusornis sanctus suggest a fish diet. Naturwissenschaften 93(9): 444–446. doi:10.1007/s00114-006-0125-y (HTML abstract)
- Hou, L; Zhou, Z; Gu, Y. & Zhang, H. (1995). [Description of Confuciusornis sanctus]. Chinese Science Bulletin 10: 61–63.
- Hou, L.-H; Zhou, Z; Martin, L.D. & Feduccia, A. (1995): A beaked bird from the Jurassic of China. Nature 377: 616–618. doi:10.1038/377616a0 (HTML abstract)
- de Ricqlès, A.J; Padian, K; Horner, J.R; Lamm, E.-T. & Myhrvold, N. (2003): Osteohistology of Confuciusornis sanctus (Theropoda: Aves). Journal of Vertebrate Paleontology 23(2): 373–386. DOI:10.1671/0272–4634(2003)023[0373:OOCSTA]2.0.CO;2 HTML abstract
- Mayr, G; Pohl, B. & Peters, D. S. (2005). A well-preserved Archaeopteryx specimen with theropod features. Science 310(5753): 1483–1486. doi:10.1126/science.1120331 PMID 16322455 (HTML abstract) Supporting Online Material
- Senter, Phil (2006): Scapular orientation in theropods and basal birds, and the origin of flapping flight. Acta Palaeontologica Polonica 51(2): 305–313. PDF fulltext
- Zhou, Z. & Zhang, F. (2003): Jeholornis compared to Archaeopteryx, with a new understanding of the earliest avian evolution. Naturwissenschaften 90: 220–225. PDF fulltext
External links
Categories:- Confuciusornithiformes
- Dinosaurs of Asia
- Cretaceous birds
- Genera of birds





{kind=link}
Wikimedia Foundation. 2010.