- Massospondylus
-
Massospondylus
Temporal range: Early Jurassic, 200–183 Ma
This artist's impression of Massospondylus depicts the animal as bipedal. Scientific classification 
Kingdom: Animalia Phylum: Chordata Class: Reptilia Superorder: Dinosauria Order: Saurischia Suborder: †Sauropodomorpha Node: †Plateosauria Branch: †Massopoda Family: †Massospondylidae Genus: †Massospondylus
Owen, 1854Species Synonyms - ?Aetonyx Broom, 1911
- ?Aristosaurus Hoepen, 1920
- ?Dromicosaurus Hoepen, 1920
- ?Gyposaurus Broom, 1911
- ?Hortalotarsus Seeley, 1894
- ?Ignavusaurus Knoll, 2010
- ?Leptospondylus Owen, 1895
- ?Pachyspondylus Owen, 1854
Massospondylus (pronounced /ˌmæsɵˈspɒndɨləs/ mas-oh-spon-di-ləs, from Greek, μάσσων (massōn, "longer") and σπόνδυλος (spondylos, "vertebra") is a genus of prosauropod dinosaur from the early Jurassic Period (Hettangian to Pliensbachian ages, ca. 200–183 million years ago). It was described by Sir Richard Owen in 1854 from remains found in South Africa, and is thus one of the first dinosaurs to have been named. Fossils have since been found at other locations in South Africa, Lesotho, and Zimbabwe. Material from Arizona's Kayenta Formation, India, and Argentina has been assigned to this genus at various times, but the Arizonan and Argentinian material are now assigned to other genera.
The type species is M. carinatus; seven other species have been named during the past 150 years, but only M. kallae among these is still considered valid. Prosauropod systematics have undergone numerous revisions during the last several years, and many scientists disagree where exactly Massospondylus lies on the dinosaur evolutionary tree. The family name Massospondylidae was once coined for the genus, but because knowledge of prosauropod relationships is in a state of flux, it is unclear which other dinosaurs—if any—belong in a natural grouping of massospondylids; several 2007 papers support the family's validity.
Although Massospondylus was long depicted as quadrupedal, a 2007 study found it to be bipedal. It was probably a plant eater (herbivore), although it is speculated that the prosauropods may have been omnivorous. This animal, 4–6 meters (13–20 feet) long, had a long neck and tail, with a small head and slender body. On each of its forefeet, it bore a sharp thumb claw that was used in defense or feeding. Recent studies indicate Massospondylus grew steadily throughout its lifespan, possessed air sacs similar to those of birds, and may have cared for its young.
Contents
Description
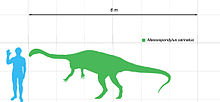 Size compared to a human
Size compared to a human
Massospondylus was a mid-sized prosauropod that was around 4 meters (13 ft) in length[1][2][3] and weighed approximately 135 kilograms (300 lb),[3] although a few sources have estimated its length at up to 6 meters (20 ft).[4][5] Although long assumed to have been quadrupedal, a 2007 anatomical study of the forelimbs has questioned this, arguing that their range of motion precluded effective habitual quadrupedal gait. The study also ruled out the possibility of "knuckle-walking" and other forms of locomotion that would avoid the issue of the limited ability of Massospondylus to pronate its hands. Although its mass suggests a quadrupedal nature, it would have been restricted to its hind legs for locomotion.[6]
Massospondylus was a typical prosauropod in most other respects. It possessed a slender body and long neck, with around nine long cervical (neck) vertebrae, 13 dorsal (back) vertebrae, three sacral (hip) vertebrae, and at least 40 caudal (tail) vertebrae. The pubis faced forward, as with most saurischians. It had a slighter build than that of Plateosaurus, an otherwise similar prosauropod dinosaur.[7] A recent discovery shows that Massospondylus possessed well-developed clavicles, joined in a furcula-like arrangement, suggesting both that it had immobile shoulder blades and that clavicles were not rudimentary and nonfunctional in those dinosaurs that did not have true furculae. This discovery also indicates that the furcula of birds is derived from clavicles.[8]
Like Plateosaurus, it had five digits on each foot, with a large thumb claw used for feeding or defense against predators. The fourth and fifth digits of the forepaws were tiny, giving the forepaws a lopsided look. The 2007 study indicated that Massospondylus held its manus (hands) in a semi-supinated ("prayer-like") orientation, with the palmar surfaces facing one another; the wrist was never found rotated in articulated (still-connected) fossil specimens.[6]
Cranial anatomy
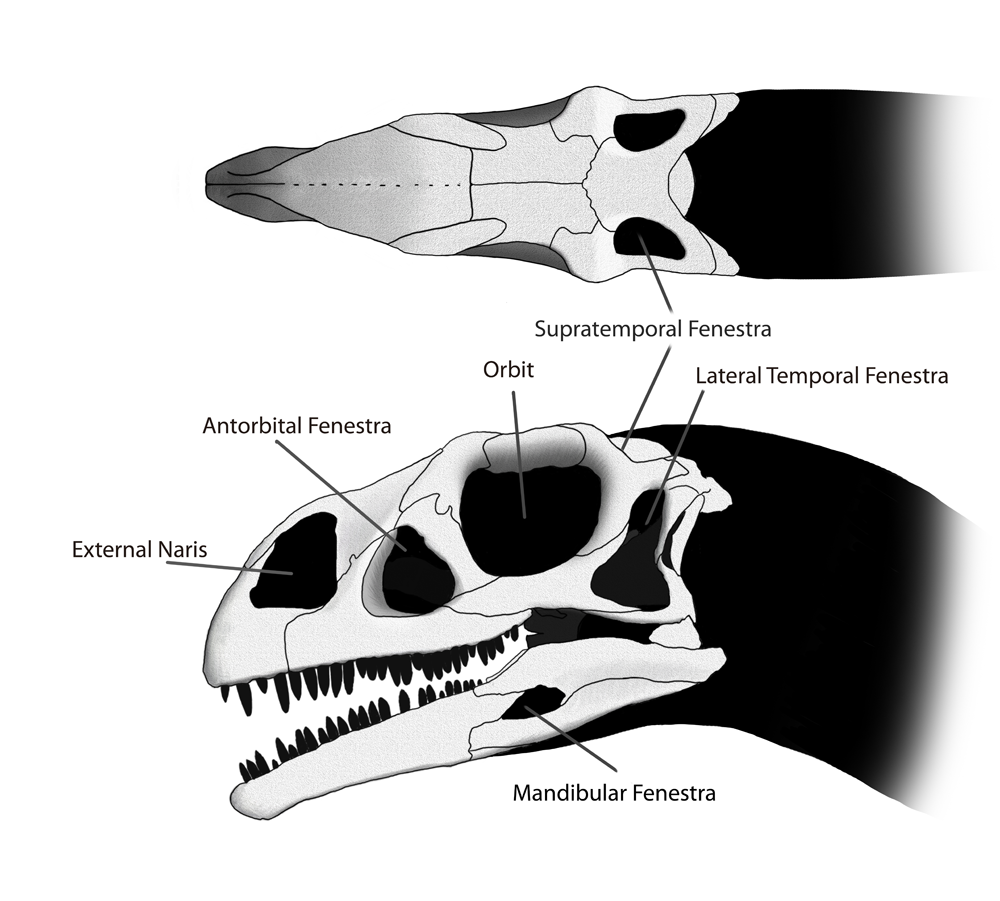
 Above is a diagram of the skull of Massospondylus, showing the various skull openings.
Above is a diagram of the skull of Massospondylus, showing the various skull openings.The small head of Massospondylus was approximately half the length of the femur. Numerous openings, or fenestrae, in the skull reduced its weight and provided space for muscle attachment and sensory organs. These fenestrae were present in pairs, one on each side of the skull. At the front of the skull were two large, elliptical nares. The orbits were proportionally larger in Massospondylus than in related genera such as Plateosaurus. The antorbital fenestrae, smaller than those seen in Plateosaurus, were situated between the eyes and the nose. At the rear of the skull were two more pairs of temporal fenestrae: the lateral temporal fenestrae immediately behind the eye sockets and the supratemporal fenestrae on top of the skull. Small fenestrae also penetrated each mandible.[7] The shape of the skull is traditionally restored as wider and shorter than that of Plateosaurus, but this appearance may be due just to differential crushing experienced by the various specimens.[7] Some features of the skull are variable between individuals; for example, the thickness of the upper border of the orbit and the height of the posterior maxilla. These differences may be due to sexual dimorphism[9] or individual variation.[10]
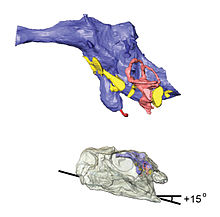 Endocast of brain cavity (above) and transparent skulls with endocasts in place (below) showing head posture, based on µCT scans of specimen BP 1/4779
Endocast of brain cavity (above) and transparent skulls with endocasts in place (below) showing head posture, based on µCT scans of specimen BP 1/4779As with other prosauropods, it has been proposed that Massospondylus had cheeks. This theory was proposed because there are a few large holes for blood vessels on the surfaces of the jaw bones, unlike the numerous small holes present on the jaws of cheekless reptiles. The cheeks would have prevented food from spilling out when Massospondylus ate.[7] Crompton and Attridge (1986) described skulls of Massospondylus as possessing pronounced overbites and suggested the presence of a horny beak on the tip of the lower jaw to make up the difference in length and account for tooth wear on the teeth at the tip of the snout.[11] However, this was later shown to be a misinterpretation based on crushing in a top–bottom plane. Skulls not crushed in this orientation do not show an overbite.[9] There also seems to be some variation of tooth morphology based on the position of teeth in the jaw.[12] The heterodonty present in Massospondylus is greater than that present in Plateosaurus, although unsurprisingly not as pronounced as the specialization of teeth in Heterodontosaurus.[12] Teeth closer to the front of the snout had round cross-sections and tapered to points, unlike the back teeth, which were spatulate and had oval cross-sections.[7][12]
Discovery
The first fossils of Massospondylus were described by paleontologist Sir Richard Owen in 1854,[13] the name having been derived from the Greek terms masson/μάσσων 'longer'[14] and spondylos/σπόνδυλος 'vertebra'.[15] These fossils were found in 1853 by J. M. Orpen in the Upper Elliot Formation at Harrismith, South Africa. Included among the remains were vertebrae from the neck, back, and tail; a shoulder blade; a humerus; a partial pelvis; a femur; a tibia; and bones of the hands and feet. The original holotype material was part of the Royal College of Surgeons collection in London and was destroyed in World War II; only casts remain.[16] Possible Massospondylus remains have been found in the Upper Elliot Formation, the Clarens Formation, and the Bushveld Sandstone of South Africa and Lesotho; the Forest Sandstone and the Upper Karroo Sandstone of Zimbabwe; and the Kayenta Formation of Arizona. These remains consist of at least 80 partial skeletons and four skulls, representing both juveniles and adults.[7]
The report of Massospondylus from Arizona's Kayenta Formation is based on a skull described in 1985. The skull of the Kayenta specimen from Arizona is 25% larger than the largest skull from any African specimen.[12] The Kayenta specimen possesses four teeth in the premaxilla and sixteen in the maxilla. Uniquely among dinosaurs, it also had tiny, one-millimeter-(0.04 in-) long palatal teeth.[17] Recent restudy of African Massospondylus skulls, however, indicates that the Kayenta specimen does not pertain to Massospondylus.[18] This Kayenta skull and associated postcranial elements, identified collectively as MCZ 8893, has been recently referred to the newly described genus Sarahsaurus.[19]
Massospondylus had also been reported from Argentina,[7] but this has been reassessed as a closely related but distinct genus. The fossils included several partial skeletons and at least one skull, found in the Lower Jurassic Canon del Colorado Formation of San Juan, Argentina. This material was named Adeopapposaurus in 2009.[20]
Species
Many species have been named, although most are no longer considered valid. M. carinatus, named by Richard Owen, is the type species.[21] Other named species include: M. browni (Seeley, 1895),[22] M. harriesi (Broom 1911),[23] M. hislopi (Lydekker, 1890),[24] M. huenei (Cooper, 1981),[25] M. kaalae (Barrett 2009),[26] M. rawesi (Lydekker, 1890),[24] and M. schwarzi (Haughton, 1924).[27]
M. browni, M. harriesi, and M. schwarzi were all found in the Upper Elliot Formation of Cape Province, South Africa. All three are based on fragmentary material, and were regarded as indeterminate in the most recent review.[7] M. browni is based on two cervical, two back, and three caudal vertebrae and miscellaneous hind limb elements.[22] M. harriesi is known from a forelimb.[23] M. schwarzi is known from an incomplete hind limb and sacrum.[27] M. hislopi and M. rawesi were named from fossils found in India.[24] M. hislopi is based on vertebrae from the Upper Triassic Maleri Formation of Andhra Pradesh, whereas M. rawesi is based on a tooth from the Upper Cretaceous Takli Formation of Maharashtra.[28] M. hislopi was tentatively retained as an indeterminate sauropodomorph in the latest review,[7] but M. rawesi may be a theropod[29] or nondinosaur.[16] M. huenei is a combination derived by Cooper for Lufengosaurus huenei, as he considered Lufengosaurus and Massospondylus to be synonyms.[25] This synonymy is no longer accepted.[7]
M. kaalae was described in 2009 on the basis of a partial skull found from the upper Elliot Formation in the Herschel District of South Africa. This species is known from the same time and region as some specimens of M. carinatus. It differs from the type species in the morphology of the braincase, as well as in several other characters of the skull such as the proportions of the premaxilla.[26]
Dubious names
Several dinosaurs are often considered synonymous with Massospondylus. These include Aristosaurus, Dromicosaurus, Gryponyx taylori, Hortalotarsus, Leptospondylus, and Pachyspondylus, which are dubious names of little scientific value.[25] Hortalotarsus skirtopodus was named by Harry Seeley in 1894. According to Broom (1911), "Originally most of the skeleton was in the rock, and it was regarded by the farmers as the skeleton of a Bushman, but it is said to have been destroyed through fear that a Bushman skeleton in the rock might tend to weaken the religious belief of the rising generation." Some partial leg bones were salvaged. The following year, Richard Owen named a few fossil vertebrae Leptospondylus capensis and Pachyspondylus orpenii. These fragmentary fossils were later destroyed in World War II. Aristosaurus erectus was named by E.C.N. van Hoepen in 1920 based on a nearly complete skeleton. Hoepen also named Dromicosaurus gracilis, which consisted of a partial skeleton. Gryponyx taylori was named by Sidney H. Haughton in 1924. It consists of hip bones. All of the above fossils come from the Hettangian or Sinemurian faunal stages of South Africa, where Massospondylus has been found.[30][31] Under the rules of zoological nomenclature, these names are junior synonyms. They were named after Massospondylus was described in a scientific paper; the name Massospondylus thus takes priority.
Ignavusaurus, known from a young specimen, may also be synonymous with Massospondylus.[32]
Classification
Plateosauria Plateosauridae Massopoda Riojasauridae (unnamed) Massospondylidae Massospondylus
(unnamed) (unnamed) Basal sauropodomorph phylogeny simplified after Yates, 2007.[33] In this review, Massospondylus is shown as closely related to Lufengosaurus and Coloradisaurus. This is only one of many proposed cladograms for basal sauropodomorphs. Massospondylus is a prosauropod, a grouping of early saurischian dinosaurs that lived during the Triassic and Jurassic, but which had died out by the end of the Jurassic. Other members of the group include Plateosaurus,[7] Yunnanosaurus,[7] and Riojasaurus.[34] Basal sauropodomorph systematics continue to undergo revision, and many genera once considered classic prosauropods have recently been removed from the group in phylogenetic nomenclature, on the grounds that their inclusion would not constitute a clade (a natural grouping containing all descendants of a single common ancestor). Exactly which animals constitute a monophyletic (natural grouping) of prosauropods is unclear. Yates and Kitching (2003) published a clade consisting of Riojasaurus, Plateosaurus, Coloradisaurus, Massospondylus, and Lufengosaurus.[35] Galton and Upchurch (2004) included Ammosaurus, Anchisaurus, Azendohsaurus, Camelotia, Coloradisaurus, Euskelosaurus, Jingshanosaurus, Lessemsaurus, Lufengosaurus, Massospondylus, Melanorosaurus, Mussaurus, Plateosaurus, Riojasaurus, Ruehleia, Saturnalia, Sellosaurus, Thecodontosaurus, Yimenosaurus and Yunnanosaurus in a monophyletic Prosauropoda.[7] Wilson (2005) considered Massospondylus, Jingshanosaurus, Plateosaurus, and Lufengosaurus a natural group, with Blikanasaurus and Antetonitrus possible sauropods.[36] Bonnan and Yates (2007) considered Camelotia, Blikanasaurus and Melanorosaurus possible sauropods.[37] Yates (2007) placed Antetonitrus, Melanorosaurus, and Blikanasaurus as basal sauropods and declined to use the term Prosauropoda, as he considered it synonymous with Plateosauridae. However, he did not rule out the possibility that a small group of prosauropods consisting of Plateosaurus, Riojasaurus, Massospondylus and their closest kin were monophyletic.[33]
Massospondylus is the type genus of the proposed family Massospondylidae, to which it gives its name. The Massospondylidae family may also include Yunnanosaurus,[38] although Lu et al. (2007) placed Yunnanosaurus in its own family.[39] Yates (2007) considered Massospondylus, Coloradisaurus, and Lufengosaurus massospondylids, with Yunnanosaurus in Anchisauria.[33] Smith and Pol (2007) also found a Massospondylidae in their phylogenetic analysis, including Massospondylus, Coloradisaurus, and Lufengosaurus, as well as their new genus, Glacialisaurus.[40] Adeopapposaurus, based on the fossils once thought to belong to a South American form of Massospondylus, was also classified as a massospondylid.[20] Pradhania was originally regarded as a basal sauropodomorph but new cladistic analysis performed by Novas et al., 2011 suggests that Pradhania is a massospondylid. Pradhania presents two synapomorphies of Masospondylidae recovered in their phylogenetic analysis and its fossils were discovered from the same region and basin in India as M. hislopi.[41]
Paleoecology
The faunas and floras of the Early Jurassic were similar worldwide, with conifers adapted for hot weather becoming the common plants,[42] and prosauropods and basal theropods the main constituents of a worldwide dinosaur fauna.[43] The environment of early Jurassic southern Africa has been described as a desert.[44] African Massospondylus was a contemporary of early crocodylomorphs; tritylodontid and trithelodontid therapsids; morganucodontid mammals;[7] and dinosaurs including the small theropod Megapnosaurus rhodesiensis; a species of Melanorosaurus (M. thabanensis); and several genera of early ornithischians, such as Lesothosaurus and the heterodontosaurids Abrictosaurus, Heterodontosaurus, and Lycorhinus.[31]
It is not clear which carnivores may have preyed on Massospondylus. Most of the theropods which have been discovered in rocks of Early Jurassic age in southern Africa, such as Megapnosaurus, were smaller than mid-sized prosauropods like Massospondylus. These smaller predators have been postulated as using fast slashing attacks to wear down prosauropods, which could have defended themselves with their large hand and foot claws.[44] The 6-meter-(20 ft-) long [45] carnivorous theropod Dracovenator lived during the same period (Hettangian to Sinemurian stages) as Massospondylus and has also been found in the Elliot Formation of South Africa.[46]
Paleobiology
 A mounted Massospondylus skeleton at the Natural History Museum, London, showing an outdated pre-2007 pose
A mounted Massospondylus skeleton at the Natural History Museum, London, showing an outdated pre-2007 poseAs with all dinosaurs, much of the biology of Massospondylus, including its behavior, coloration, and physiology, remains unknown. However, recent studies have allowed for informed speculation on subjects such as growth patterns,[47] diet,[48] posture,[6] reproduction,[49] and respiration.[50]
A 2007 study suggested that Massospondylus may have used its short arms in defense from predators ("defensive swats"), in intraspecies combat, or in feeding, although its arms were too short to reach its mouth. Scientists speculate that Massospondylus could have used its large pollex (thumb) claw in combat, to strip plant material from trees,[6] digging, or for grooming.[12]
Growth
 Skulls casts of adult (the neotype) and juvenile M. carinatus
Skulls casts of adult (the neotype) and juvenile M. carinatusA 2005 study indicated that Massospondylus' sister taxon, Plateosaurus, exhibited growth patterns affected by environmental factors. The study indicated that when food was plentiful, or when the climate was favorable, Plateosaurus exhibited accelerated growth. This pattern of growth is called "developmental plasticity". It is unseen in other dinosaurs, including Massospondylus, despite the close relationship between Plateosaurus and Massospondylus. The study indicated that Massospondylus grew along a specific growth trajectory with little variation in the growth rate and ultimate size of an individual.[47] Another study of age determination indicated that Massospondylus grew at a maximum rate of 34.6 kg (76.3 lb) per year and was still growing at around 15 years of age.[51]
Diet
Prosauropods such as Massospondylus may have been herbivorous or omnivorous. As recently as the 1980s, paleontologists debated the possibility of carnivory in prosauropods.[17][25] However, the hypothesis of carnivorous prosauropods has been discredited, and all recent studies favor a herbivorous or omnivorous lifestyle for these animals. Galton and Upchurch (2004) found that cranial characteristics (such as jaw articulation) of most prosauropods are closer to those of herbivorous reptiles than those of carnivorous ones, and the shape of the tooth crown is similar to those of modern herbivorous or omnivorous iguanas. The maximum width of the crown was greater than that of the root, resulting in a cutting edge similar to those of extant herbivorous or omnivorous reptiles.[7] Barrett (2000) proposed that prosauropods supplemented their herbivorous diets with small prey or carrion.[52] Gastroliths (gizzard stones) have been found in association with Massospondylus fossils in South Africa,[12] and with a Massospondylus-like animal from the Late Triassic of Virginia.[48] Massospondylus swallowed these stones to aid in digestion;[12] muscular contractions in the creature's gizzard would have pulverized swallowed plant material, compensating for its inability to chew.
Reproduction
 An artist's depiction of a juvenile Massospondylus, shown here as a quadruped
An artist's depiction of a juvenile Massospondylus, shown here as a quadrupedIn 1977, seven 190-million-year-old Massospondylus eggs were found in Golden Gate Highlands National Park in South Africa by James Kitching, who identified them as most likely belonging to Massospondylus. It was nearly 30 years before extraction was started on the fossils of the 15-centimeter- (6 in-) long embryos. They remain the oldest dinosaur embryos ever found. Notably, the near-hatchlings had no teeth, suggesting they had no way of feeding themselves. Based on the lack of teeth and the animal's body proportions, scientists speculate that postnatal care might have been necessary. The four legs of the near-hatchlings were of equal length, indicating that newly hatched Massospondylus were quadrupedal.[49] The skull and eyes were proportionately oversized when compared to adults, as is common in other very young vertebrates, although the skulls of juveniles were taller and more narrow as well.[12] The quadrupedality of the hatchlings suggests that the quadrupedal posture of later sauropods may have evolved from retention of juvenile characteristics in adult animals, an evolutionary phenomenon known as pedomorphosis.[49]
Respiratory system
Many saurischian dinosaurs possessed vertebrae and ribs that contained hollowed-out cavities (pneumatic foramina), which reduced the weight of the bones and may have served as a basic 'flow-through ventilation' system similar to that of modern birds. In such a system, the neck vertebrae and ribs are hollowed out by the cervical air sac; the upper back vertebrae, by the lung; and the lower back and sacral (hip) vertebrae, by the abdominal air sac. These organs constitute a complex and very efficient method of respiration.[53] Prosauropods are the only major group of saurischians without an extensive system of pneumatic foramina. Although possible pneumatic indentations have been found in Plateosaurus and Thecodontosaurus, the indentations were very small. One study in 2007 concluded that prosauropods like Massospondylus likely had abdominal and cervical air sacs, based on the evidence for them in sister taxa (theropods and sauropods). The study concluded that it was impossible to determine whether prosauropods had a bird-like flow-through lung, but that the air sacs were almost certainly present.[50]
References
- ^ Lambert, David; and the Diagram Group (1983). A Field Guide to Dinosaurs. New York: Avon Books. p. 103. ISBN 0-380-83519-3.
- ^ Lambert, David; and the Diagram Group (1990). The Dinosaur Data Book. New York: Avon Books. p. 75. ISBN 0-380-75896-3.
- ^ a b Seebacher, Frank (2001). "A new method to calculate allometric length-mass relationships of dinosaurs". Journal of Vertebrate Paleontology 21 (1): 51–60. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2.
- ^ Lessem, Don; Donald F. Glut (1993). "Massospondylus". The Dinosaur Society Dinosaur Encyclopedia. Random House. p. 295. ISBN 0-679-41770-2.
- ^ Glut, Donald F. (2000). "Massospondylus". Dinosaurs: The Encyclopedia: Supplement One. Jefferson, North Carolina: McFarland & Co. p. 258. ISBN 0-7864-0591-0.
- ^ a b c d Bonnan, Matthew F.; and Senter, Phil (2007). "Were the basal sauropodomorph dinosaurs Plateosaurus and Massospondylus habitual quadrupeds?". In Paul M. Barrett & D. J. Batten (eds.). Evolution and Palaeobiology of Early Sauropodomorph Dinosaurs. Special Papers in Palaeontology 77. London: The Palaeontological Association. pp. 139–155. ISBN 978-1-4051-6933-2.
- ^ a b c d e f g h i j k l m n o Galton, P.M. and Upchurch, P. (2004). "Prosauropoda". Weishampel & als: The Dinosauria (2nd edition), pp. 232–258.
- ^ Yates, Adam M.; and Vasconcelos, Cecilio C. (2005). "Furcula-like clavicles in the prosauropod dinosaur Massospondylus". Journal of Vertebrate Paleontology 25 (2): 466–468. doi:10.1671/0272-4634(2005)025[0466:FCITPD]2.0.CO;2.
- ^ a b Gow, Christoper E.; J.W. Kitching and Michael K. Raath (1990). "Skulls of the prosauropod dinosaur Massospondylus carinatus Owen in the collections of the Bernard Price Institute for Palaeontological Research". Palaeontologia Africana 27: 45–58.
- ^ Galton, Peter M. (1997). "Comments on sexual dimorphism in the prosauropod dinosaur Plateosaurus engelhardti (Upper Triassic, Trossingen)". Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 11: 674–682.
- ^ Crompton, A.W.; John Attridge (1986). "Masticatory apparatus of the larger herbivores during Late Triassic and Early Jurassic times". In Kevin Padian (ed.). The Beginning of the Age of Dinosaurs: Faunal Change Across the Triassic–Jurassic Boundary. New York: Cambridge University Press. pp. 223–236. ISBN 0-521-36779-4.
- ^ a b c d e f g h "Massospondylus." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 39. ISBN 0-7853-0443-6.
- ^ Owen, Richard (1854). "Descriptive catalogue of the Fossil organic remains of Reptilia and Pisces contained in the Museum of the Royal College of Surgeons of England." London p. 1–184 OCLC 14825172
- ^ Creisler, Ben (2003). "Dinosauria Translation and Pronunciation Guide". Dinosauria.com. http://www.dinosauria.com/dml/names/dinom.htm. Retrieved 2007-11-23.
- ^ Liddell, Henry George; Robert Scott (1980). A Greek-English Lexicon (Abridged Edition). Oxford: Oxford University Press. ISBN 0-19-910207-4. OCLC 17396377.
- ^ a b Glut, Donald F. (1997). "Massospondylus". Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co. pp. 581–586. ISBN 0-89950-917-7.
- ^ a b Attridge, J.; A.W. Crompton and Farish A. Jenkins, Jr. (1985). "The southern Liassic prosauropod Massospondylus discovered in North America". Journal of Vertebrate Paleontology 5 (2): 128–132. doi:10.1080/02724634.1985.10011850.
- ^ Sues, H.-D.; R.R. Reisz, S. Hinic and M.A. Raath (2004). "On the skull of Massospondylus carinatus Owen, 1854 (Dinosauria: Sauropodomorpha) from the Elliot and Clarens formations (Lower Jurassic) of South Africa". Annals of Carnegie Museum 73 (4): 239–257.
- ^ Timothy B. Rowe, Hans-Dieter Sues and Robert R. Reisz (2010). "Dispersal and diversity in the earliest North American sauropodomorph dinosaurs, with a description of a new taxon". Proc. R. Soc. B In press. doi:10.1098/rspb.2010.1867. PMC 3049036. PMID 20926438. http://rspb.royalsocietypublishing.org/content/early/2010/09/29/rspb.2010.1867.abstract.
- ^ a b Martínez, Ricardo N. (2009). "Adeopapposaurus mognai, gen. et sp. nov (Dinosauria: Sauropodomorpha), with comments on adaptations of basal sauropodomorpha". Journal of Vertebrate Paleontology 29 (1): 142–164. doi:10.1671/039.029.0102.
- ^ "Massospondylus". The Paleobiology Database. 2007. http://paleodb.org/cgi-bin/bridge.pl?action=checkTaxonInfo&taxon_no=38642&is_real_user=1. Retrieved 2007-11-03.
- ^ a b Seeley, H.G. (1895). "On the type of the genus Massospondylus and on some Vertebrae and limb-bone of M. (?) browni". Annals and Magazine of Natural History 15: 102–125.
- ^ a b Broom, Robert (1911). "On the dinosaurs of the Stormberg, South Africa". Annals of the South African Museum 7 (4): 291–308.
- ^ a b c Lydekker, Richard (1890). "Note on certain vertebrate remains from the Nagpur district". Records of the Geological Survey of India 23 (1): 21–24.
- ^ a b c d Cooper, M.R. (1980). "The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance". Occasional Papers of the National Museums and Monuments of Rhodesia, Series B, Natural Sciences 6 (10): 689–840.
- ^ a b Barrett, P. M. (2009). "A new basal sauropodomorph dinosaur from the Upper Elliot Formation (Lower Jurassic) of South Africa". Journal of Vertebrate Paleontology 29 (4): 1032–1045. doi:10.1671/039.029.0401.
- ^ a b Haughton, Sydney H. (1924). "The fauna and stratigraphy of the Stormberg Series". Annals of the South African Museum 12: 323–497.
- ^ Weishampel, David B. (1990). "Dinosaur distribution". In David B. Weishampel, Peter Dodson and Halszka Osmólska (eds.). The Dinosauria (1st ed.). Berkeley: University of California Press. pp. 63–139. ISBN 0-520-06727-4.
- ^ Molnar, R.E. (1990). Problematic Theropoda: "Carnosaurs". In Weishampel & als: The Dinosauria (1st ed.), pp. 306–317.
- ^ Tweet, Justin (2007). "Sauropodomorpha". Thescelosaurus!. http://www.thescelosaurus.com/sauropodomorpha.htm. Retrieved 2007-12-16.
- ^ a b Weishampel, David B.; Paul M. Barrett, Rodolfo Coria, A., Jean Le Loeuff, Zhao Xijin Xu Xing, Ashok Sahni, Elizabeth M.P. Gomani and Christopher R. Noto (2004). "Dinosaur Distribution". In David B. Weishampel, Peter Dodson and Halszka Osmólska (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 517–606. ISBN 0-520-24209-2.
- ^ Yates, Adam M.; Matthew F. Bonnan and Johann Neveling (2011). "A new basal sauropodomorph dinosaur from the Early Jurassic of South Africa". Journal of Vertebrate Paleontology 31 (3): 610–625. doi:10.1080/02724634.2011.560626. http://www.informaworld.com/smpp/content~db=all~content=a937433139~frm=titlelink.
- ^ a b c Yates, Adam M., "The first complete skull of the Triassic dinosaur Melanorosaurus Haughton (Sauropodomorpha: Anchisauria)". In Barrett & Batten (eds.), Evolution and Palaeobiology (2007), pp. 9–55.
- ^ Galton, P.M; J. Van Heerden and A.M. Yates (2005). "Postcranial anatomy of referred specimens of the sauropodomorph dinosaur Melanorosaurus from the Upper Triassic of South Africa". In Virginia Tidwell and Kenneth Carpenter (eds.). Thunder-Lizards: The Sauropodomorph Dinosaurs. Bloomington: Indiana University Press. pp. 1–37. ISBN 0-253-34542-1.
- ^ Yates, Adam M.; James W. Kitching (2003). "The earliest known sauropod dinosaur and the first steps towards sauropod locomotion". Proceedings of the Royal Society B 270 (1525): 1753–1758. doi:10.1098/rspb.2003.2417. PMC 1691423. PMID 12965005. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1691423.
- ^ Wilson, Jeffrey A.; Curry Rogers, Kristina (2005). The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press. pp. 15–49. ISBN 978-0-520-24623-2.
- ^ Bonnan, Matthew F. and Adam M. Yates, "A new description of the forelimbs of the basal sauropodomorph Melanorosaurus: implications for the evolution of pronation, manus shape and quadrupedalism in sauropod dinosaurs". In Barrett & Batten (eds.), Evolution and Palaeobiology (2007), pp. 157–168.
- ^ Sereno, Paul C. (1999). "The evolution of dinosaurs". Science 284 (5423): 2137–2147. doi:10.1126/science.284.5423.2137. PMID 10381873.
- ^ Lu, J.; T. Li, S. Zhong, Y. Azuma, M. Fujita, Z Dong and Q. Ji (2007). "New yunnanosaurid dinosaur (Dinosauria, Prosauropoda) From the Middle Jurassic Zhanghe Formation of Yuanmou, Yunnan province of China". Memoir of the Fukui Prefectural Dinosaur Museum 6: 1–15.
- ^ Smith, Nathan D.; Diego Pol (2007). "Anatomy of a basal sauropodomorph dinosaur from the Early Jurassic Hanson Formation of Antarctica" (pdf). Acta Palaeontologica Polonica 52 (4): 657–674. http://www.app.pan.pl/acta52/app52-657.pdf.
- ^ Fernando E. Novas, Martin D. Ezcurra, Sankar Chatterjee and T. S. Kutty (2011). "New dinosaur species from the Upper Triassic Upper Maleri and Lower Dharmaram formations of central India". Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101 (3-4): 333–349. doi:10.1017/S1755691011020093.
- ^ Russell, Dale A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. p. 45. ISBN 1-55971-038-1.
- ^ Holtz, Thomas R., Jr., Ralph E. Chapman and Matthew C. Lamanna, "Mesozoic biogeography of Dinosauria". In Weishampel & als., The Dinosauria (2nd edition), pp. 627–642.
- ^ a b Paul, Gregory S. (1988). "Coelophysis rhodesiensis". Predatory Dinosaurs of the World. New York: Simon and Schuster. pp. 262–265. ISBN 978-0-671-61946-6.
- ^ Smith, N.D.; P.J. Makovicky, D. Pol, W.R. Hammer and P.J. Currie (2007). "The Dinosaurs of the Early Jurassic Hanson Formation of the Central Transantarctic Mountains: Phylogenetic Review and Synthesis". In Alan Cooper, Carol Raymond et als. (eds.) (PDF). Antarctica: A Keystone in a Changing World—Online Proceedings for the Tenth International Symposium on Antarctic Earth Sciences. Reston, Virginia: U.S. Geological Survey. doi:doi:10.3133/of2007-1047.srp003. ISBN 1-4113-1788-2. OCLC 85782509. http://pubs.usgs.gov/of/2007/1047/srp/srp003/of2007-1047srp003.pdf.
- ^ Yates, A.M. (2006). "A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods". Palaeontologia africana 41: 105–122.
- ^ a b Sander, P. Martin; Nicole Klein (2005). "Developmental plasticity in the life history of a prosauropod dinosaur". Science 310 (5755): 1800–1802. Bibcode 2005Sci...310.1800S. doi:10.1126/science.1120125. PMID 16357257.
- ^ a b Weems, Robert E.; Michelle J. Culp and Oliver Wings (2007). "Evidence for Prosauropod Dinosaur Gastroliths in the Bull Run Formation (Upper Triassic, Norian) of Virginia". Ichnos 13 (3–4): 271–295. doi:10.1080/10420940601050030.
- ^ a b c Reisz, Robert R.; Diane Scott, Hans-Dieter Sues, David C. Evans, Michael A. Raath (2005). "Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance". Science 309 (5735): 761–764. Bibcode 2005Sci...309..761R. doi:10.1126/science.1114942. PMID 16051793.
- ^ a b Wedel, Mathew (2007). "What pneumaticity tells us about 'prosauropods', and vice versa" (PDF). Special Papers in Palaeontology 77: 207–222. http://sauroposeidon.net/Wedel_2007_pneumaticity-and-prosauropods.pdf. Retrieved 2007-10-31.
- ^ Erickson, Gregory M.; Kristina Curry Rogers and Scott A. Yerby (2001). "Dinosaurian growth patterns and rapid avian growth rates". Nature 412 (6845): 429–433. doi:10.1038/35086558. PMID 11473315.
- ^ Barrett, P.M. (2000). "Prosauropod dinosaurs and iguanas: Speculations on the diets of extinct reptiles". In Hans-Dieter Sues. Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record. Cambridge: Cambridge University Press. pp. 42–78. ISBN 978-0-521-59449-3.
- ^ O'Connor, Patrick M.; Leon P.A.M. Claessens (2006). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature 436 (7048): 253–256. Bibcode 2005Natur.436..253O. doi:10.1038/nature03716. PMID 16015329.
Further reading
- Chinsamy, A. (1992). "Ontogenetic growth of the dinosaurs Massospondylus carinatus and Syntarsus rhodesiensis". In: Abstracts of papers. Society of Vertebrate Paleontology, fifty-second annual meeting. Royal Ontario Museum Toronto, Ontario". Journal of Vertebrate Paleontology 12 (3): 23A.
- Gow, C. E. (1990). "Morphology and growth of the Massospondylus braincase (Dinosauria, Prosauropoda)". Palaeontologia Africana 27: 59–75.
- Hinic, S. (2002). "The cranial anatomy of Massospondylus carinatus Owen, 1854 and its implications for prosauropod phylogeny". Journal of Vertebrate Paleontology. Abstracts of papers. Society of Vertebrate Paleontology, 22, Supplement to number 3, 65A.
- Martínez, R. (1999). "The first South American record of Massospondylus (Dinosauria: Sauropodomorpha)". Journal of Vertebrate Paleontology, Abstracts of papers. Society of Vertebrate Paleontology, 20–23 October, 19, Suppl. 3, 61A.
- Martínez, R.N. (1999). "Massospondylus (Dinosauria: Sauropodomorpha) in northwestern Argentina". Abstracts VII International Symposium on Mesozoic Terrestrial Ecosystems, Buenos Aires, 40.
External links
- Locomotion and Ontogeny of Massospondylus carinatus from the University of Toronto at Missisauga.
- Massospondylidae from Palaeos.com (technical).
- Massospondylus at DinoData.org.
- Massospondylus from PrimeOrigins.co.za (for children).
- "Oldest known dinosaur embryos identified" University of the Witwatersrand.
- "Rare fossil embryos reveal growth" at the Wayback Machine (archived May 3, 2007) National Geographic.
- Sauropodomorpha on Thescelosaurus!
Categories:- Dinosaurs of Africa
- Jurassic dinosaurs
- Prosauropods





Wikimedia Foundation. 2010.