- Skeletal striated muscle
-
Skeletal striated muscle 
A top-down view of skeletal muscle Latin textus muscularis striatus skeletalis Code TH H2.00.05.2.00002 Skeletal muscle is a form of striated muscle tissue existing under control of the somatic nervous system- i.e. it is voluntarily controlled. It is one of three major muscle types, the others being cardiac and smooth muscle. As their name suggests, most skeletal muscles are attached to bones by bundles of collagen fibers known as tendons.
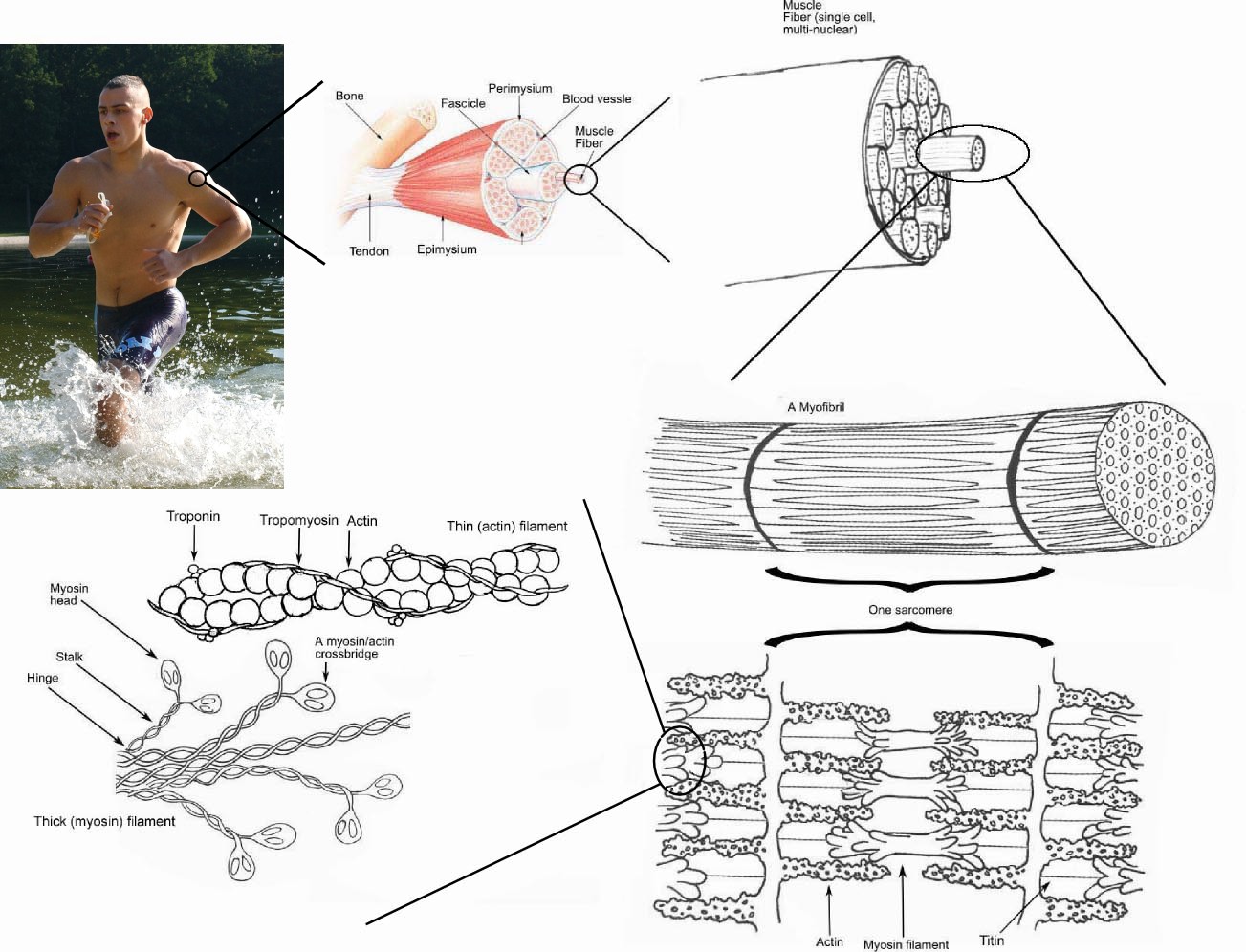
Skeletal muscle is made up of individual components known as muscle fibers. These fibers are formed from the fusion of developmental myoblasts (a type of embryonic progenitor cell that gives rise to a muscle cell). The muscle fibers are long, cylindrical, multinucleated cells composed of myofibrils. The myofibrils are composed of actin and myosin myofibrils repeated as a sarcomere, the basic functional unit of the muscle fiber and responsible for skeletal muscle's striated appearance and forming the basic machinery necessary for muscle contraction. The term muscle refers to multiple bundles of muscle fibers held together by connective tissue.
Contents
Muscle fibers
Individual muscle fibers are formed during development from the fusion of several undifferentiated immature cells known as myoblasts into long, cylindrical, multi-nucleated cells. Differentiation into this state is primarily completed before birth with the cells continuing to grow in size thereafter. Skeletal muscle exhibits a distinctive banding pattern when viewed under the microscope due to the arrangement of cytoskeletal elements in the cytoplasm of the muscle fibers. The principal cytoplasmic proteins are myosin and actin (also known as "thick" and "thin" filaments, respectively) which are arranged in a repeating unit called a sarcomere. The interaction of myosin and actin is responsible for muscle contraction.
There are two principal ways to categorize muscle fibers: the type of myosin (fast or slow) present, and the degree of oxidative phosphorylation that the fiber undergoes. Skeletal muscle can thus be broken down into two broad categories: Type I and Type II. Type I fibers appear red due to the presence of the oxygen binding protein myoglobin. These fibers are suited for endurance and are slow to fatigue because they use oxidative metabolism to generate ATP. Type II fibers are white due to the absence of myoglobin and a reliance on glycolytic enzymes. These fibers are efficient for short bursts of speed and power and use both oxidative metabolism and anaerobic metabolism depending on the particular sub-type. These fibers are quicker to fatigue.
Type I fibers Type II a fibers Type II x fibers Type II b fibers Contraction time Slow Moderately Fast Fast Very fast Size of motor neuron Small Medium Large Very large Resistance to fatigue High Fairly high Intermediate Low Activity Used for Aerobic Long-term anaerobic Short-term anaerobic Short-term anaerobic Maximum duration of use Hours <30 minutes <5 minutes <1 minute Power produced Low Medium High Very high Mitochondrial density Very High High Medium Low Capillary density High Intermediate Low Low Oxidative capacity High High Intermediate Low Glycolytic capacity Low High High High Major storage fuel Triglycerides Creatine phosphate, glycogen ATP, Creatine phosphate, glycogen (little) ATP, Creatine phosphate Note Consume lactic acid Produce lactic acid and Creatine phosphate Consume Creatine phosphate Consume Creatine phosphate Myosin heavy chain,
human genesMYH7 MYH2 MYH1 MYH4 Skeletal muscle fibers are not all the same. Traditionally, they were categorized depending on their varying color, which is a reflection of myoglobin content.
Red Fibers: Those containing high levels of myoglobin and oxygen storing proteins had a red appearance. Red muscle fibers tend to have more mitochondria and greater local capillary density.
White Fibers: Those with a low content had a white appearance.
As more was learned about the functional differences between skeletal muscle fibers, they were also classified, depending on their twitch capabilities, into fast and slow twitch, traits that largely, but not completely, overlap the previous classification based on color.
Fast Twitch: Some authors define a fast twitch fiber as one in which the myosin can split ATP very quickly.
However, fast twitch fibers also demonstrate a higher capability for electrochemical transmission of action potentials and a rapid level of calcium release and uptake by the sarcoplasmic reticulum. The fast twitch fibers rely on a well-developed, short term, glycolytic system for energy transfer and can contract and develop tension at 2-3 times the rate of slow twitch fibers.
Slow Twitch: The slow twitch fibers generate energy for ATP re-synthesis by means of a long term system of aerobic energy transfer. They tend to have a low activity level of ATPase, a slower speed of contraction with a less well developed glycolytic capacity. They contain large and numerous mitochondria and with the high levels of myoglobin that gives them a red pigmentation. They have been demonstrated to have high concentration of mitochondrial enzymes, thus they are fatigue resistant.
The 2 main categories of muscle fibers become several, when further differentiating type II into several subtypes, based on myosin isoforms and denoted with letters of the alphabet. In humans the two subtypes are IIa and IIx; IIx is often referred to as IIb because earlier classification had clumped together two different types. Later on, further research recognized these subtypes as distinct, but the use of the name IIb remained entrenched.[1] Non human fiber types include true IIb fibers, IIc, IId and so on.
Type I Red fibers. Slow oxidative (also called slow twitch or fatigue resistant fibers). Contain:
- Large amounts of myoglobin.
- Many mitochondria.
- Many blood capillaries.
- Generate ATP by the aerobic system, hence the term oxidative fibers.
- Split ATP at a slow rate.
- Slow contraction velocity.
- Resistant to fatigue.
- Found in large numbers in postural muscles.
- Needed for aerobic activities like long distance running.
Type IIa Red fibers. Fast oxidative (also called fast twitch A or fatigue resistant fibers). Contain:
- Large amounts of myoglobin.
- Many mitochondria.
- Many blood capillaries.
- Large amount of glycogen.
- High capacity for generating ATP by oxidation. Split ATP at a very rapid rate and, hence, high contraction velocity
- Resistant to fatigue but not as much as slow oxidative fibers.
- Needed for sports such as middle distance running and swimming.
Type IIx / IIb (dependent upon species) White. Fast glycolytic (also called fast twitch B or fatigable fibers). Contain:
- Low myoglobin content.
- Few mitochondria.
- Few blood capillaries.
- Large amount of Creatine phosphate.
- Split ATP very quickly.
- Fatigue easily.
- Needed for sports like sprinting.
Individual muscles are a mixture of 3 types of muscle fibers (type 1, type 2a and type 2b), but their proportions vary depending on the action of that muscle. It must be remembered that skeletal muscles, although a mixture, can only have one type of muscle fiber within a motor unit. This is demonstrated if we look at contractions. E.g. If a weak contraction is needed only the type 1 motor units will be activated. These fibers are used mainly for endurance activities. If a stronger contraction is required the type 2a fibers will be activated or used to assist the type 1 fibers. Maximal contractions facilitate the use of type 2b fibers which are always activated last. These fibers are used during ballistic activities but tire easily. With advanced EMG techniques it is possible to look at which muscle fibers are recruited when performing an exercise/test. The total number of skeletal muscle fibers has traditionally been thought not to change. It is believed there are no sex or age differences in fiber distribution, however, relative fiber types vary considerably from muscle to muscle and person to person. Sedentary men and women (as well as young children) have 45% type 2 and 55% type 1 fibers.[citation needed] People at the higher end of any sport tend to demonstrate patterns of fiber distribution e.g. endurance athletes show a higher level of type 1 fibers. Sprint athletes, on the other hand, require large numbers of type 2 b fibers. Middle distance event athletes show approximately equal distribution of the 2 types. This is also often the case for power athletes such as throwers and jumpers. It has been suggested that various types of exercise can induce changes in the fibers of a skeletal muscle.[citation needed] It is thought that if you perform endurance type events for a sustained period of time, some of the type 2b fibers transform into type 2a fibers. However, there is no consensus on the subject. It may well be that the type 2b fibers show enhancements of the oxidative capacity after high intensity endurance training which brings them to a level at which they are able to perform oxidative metabolism as effectively as slow twitch fibers of untrained subjects. This would be brought about by an increase in mitochondrial size and number and the associated related changes not a change in fiber type.
Structure of skeletal muscle fiber
Every organelle and macromolecule of a muscle fiber are arranged to ensure form meets function. The plasma membrane is called the sarcolemma with the cytoplasm known as the sarcoplasm. In the sarcoplasm are the myofibrils. The myofibrils are long protein bundles about 1 micrometer in diameter each containing myofilaments. Pressed against the inside of the sarcolemma are the unusual flattened nuclei. Between the myofibrils are the mitochondria. While the muscle fiber does not have a smooth endoplasmic reticulum it contains a sarcoplasmic reticulum. The sarcoplasmic reticulum surrounds the myofibrils and holds a reserve of the calcium ions needed to cause a muscle contraction. Periodically it has dilated end sacs known as terminal cisternae. These cross the muscle fiber from one side to the other. In between two terminal cisternae is a tubular infoldings called a transverse tubule (T tubule). The T tubule are the pathway for the action potential to signal the sarcoplasmic reticulum to release calcium causing a muscle contraction. Together two terminal cisternae and a transverse tubule form a triad.[2]
Organization of skeletal muscle fibers
While the muscle fibers of the fascicles lie parallel to one another, the fascicles themselves can vary in their relationship to one another and to their tendons.[3] The different patterns of arrangement of the fasciles produce four different types of skeletal muscles: parallel muscles, convergent muscles, pennate muscles, and sphincter muscles.[3]
- Parallel muscles
The fascicles of parallel muscles run parallel to the direction of the muscle, thus these muscles on a whole function similarly to a single muscle fiber.[3] Most skeletal muscles in the body are parallel muscles; although they can be seen in a variety of shapes such as flat bands, spindle shaped, and some can have large protrusions in their middle known as the belly of the muscle.[3]
- ex: Triceps, biceps
- Convergent muscles
The fibers in convergent muscles fan out from a common point of attachment.[3] Covering a broad surface these fibers allow for more versatile types of movement.[3] These muscles do not pull as hard on their corresponding tendons as their parallel muscle counterparts however due to the fibers not all pulling in the same direction, even pulling in different directions at opposite ends.[3]
- ex: Pectoralis muscles
- Pennate muscles
See also: Pennate muscleIn a pennate muscle one or more tendons run through the body of the muscle with the fascicles forming an oblique angle to the tendons.[3] Because the fascicles pull on the tendons at an angle they do not move the tendon as far as their parallel muscle counterparts. Despite this they generate greater tension due to their possessing a greater amount of muscle fibers than similarly sized parallel muscles.[3]
- ex: Rectus femoris
- Sphincter muscles
See also: SphincterThe fibers of the sphincter or circular muscles are arranged concentrically around an opening or recess.[3] As the muscle contracts, the opening it circumvents gets smaller, for this reason these muscles are often found at the entrances and exits of external and internal passage ways.[3]
- ex: Orbicularis oris
Cellular physiology and contraction
In addition to the actin and myosin components that constitute the sarcomere, skeletal muscle fibers also contain two other important regulatory proteins, troponin and tropomyosin, that are necessary for muscle contraction to occur. These proteins are associated with actin and cooperate to prevent its interaction with myosin. Skeletal muscle cells are excitable and are subject to depolarization by the neurotransmitter acetylcholine, released at the neuromuscular junction by motor neurons.[4]
Once a cell is sufficiently stimulated, the cell's sarcoplasmic reticulum releases ionic calcium (Ca2+), which then interacts with the regulatory protein troponin. Calcium-bound troponin undergoes a conformational change that leads to the movement of tropomyosin, subsequently exposing the myosin-binding sites on actin. This allows for myosin and actin ATP-dependent cross-bridge cycling and shortening of the muscle.
Physics
Muscle force is proportional to physiologic cross-sectional area (PCSA), and muscle velocity is proportional to muscle fiber length.[5] The strength of a joint, however, is determined by a number of biomechanical parameters, including the distance between muscle insertions and pivot points and muscle size. Muscles are normally arranged in opposition so that as one group of muscles contract, another group relaxes or lengthens. Antagonism in the transmission of nerve impulses to the muscles means that it is impossible to fully stimulate the contraction of two antagonistic muscles at any one time. During ballistic motions such as throwing, the antagonist muscles act to 'brake' the agonist muscles throughout the contraction, particularly at the end of the motion. In the example of throwing, the chest and front of the shoulder (anterior Deltoid) contract to pull the arm forward, while the muscles in the back and rear of the shoulder (posterior Deltoid) also contract and undergo eccentric contraction to slow the motion down to avoid injury. Part of the training process is learning to relax the antagonist muscles to increase the force input of the chest and anterior shoulder.
Contracting muscles produce vibration and sound.[6] Slow twitch fibers produce 10 to 30 contractions per second (10 to 30 Hz). Fast twitch fibers produce 30 to 70 contractions per second (30 to 70 Hz).[7] The vibration can be witnessed and felt by highly tensing one's muscles, as when making a firm fist. The sound can be heard by pressing a highly tensed muscle against the ear, again a firm fist is a good example. The sound is usually described as a rumbling sound. Some individuals can voluntarily produce this rumbling sound by contracting the tensor tympani muscle of the middle ear. The rumbling sound can also be heard when the neck or jaw muscles are highly tensed.
Signal transduction pathways
Skeletal muscle fiber-type phenotype in adult animals, and probably people, is regulated by several independent signaling pathways. These include pathways involved with the Ras/mitogen-activated protein kinase (MAPK), calcineurin, calcium/calmodulin-dependent protein kinase IV, and the peroxisome proliferator γ coactivator 1 (PGC-1). The Ras/MAPK signaling pathway links the motor neurons and signaling systems, coupling excitation and transcription regulation to promote the nerve-dependent induction of the slow program in regenerating muscle. Calcineurin, a Ca2+/calmodulin-activated phosphatase implicated in nerve activity-dependent fiber-type specification in skeletal muscle, directly controls the phosphorylation state of the transcription factor NFAT, allowing for its translocation to the nucleus and leading to the activation of slow-type muscle proteins in cooperation with myocyte enhancer factor 2 (MEF2) proteins and other regulatory proteins. Calcium-dependent Ca2+/calmodulin kinase activity is also upregulated by slow motor neuron activity, possibly because it amplifies the slow-type calcineurin-generated responses by promoting MEF2 transactivator functions and enhancing oxidative capacity through stimulation of mitochondrial biogenesis.
Contraction-induced changes in intracellular calcium or reactive oxygen species provide signals to diverse pathways that include the MAPKs, calcineurin and calcium/calmodulin-dependent protein kinase IV to activate transcription factors that regulate gene expression and enzyme activity in skeletal muscle.
 Exercise-Included Signaling Pathways in Skeletal Muscle That Determine Specialized Characteristics of ST and FT Muscle Fibers
Exercise-Included Signaling Pathways in Skeletal Muscle That Determine Specialized Characteristics of ST and FT Muscle Fibers
PGC1-α (PPARGC1A), a transcriptional coactivator of nuclear receptors important to the regulation of a number of mitochondrial genes involved in oxidative metabolism, directly interacts with MEF2 to synergistically activate selective ST muscle genes and also serves as a target for calcineurin signaling. A peroxisome proliferator-activated receptor δ (PPARδ)-mediated transcriptional pathway is involved in the regulation of the skeletal musclefiber phenotype. Mice that harbor an activated form of PPARd display an “endurance” phenotype, with a coordinated increase in oxidative enzymes and mitochondrial biogenesis and an increased proportion of ST fibers. Thus—through functional genomics—calcineurin, calmodulin-dependent kinase, PGC-1α, and activated PPARδ form the basis of a signaling network that controls skeletal muscle fiber-type transformation and metabolic profiles that protect against insulin resistance and obesity.
The transition from aerobic to anaerobic metabolism during intense work requires that several systems are rapidly activated to ensure a constant supply of ATP for the working muscles. These include a switch from fat-based to carbohydrate-based fuels, a redistribution of blood flow from nonworking to exercising muscles, and the removal of several of the by-products of anaerobic metabolism, such as carbon dioxide and lactic acid. Some of these responses are governed by transcriptional control of the FT glycolytic phenotype. For example, skeletal muscle reprogramming from an ST glycolytic phenotype to an FT glycolytic phenotype involves the Six1/Eya1 complex, composed of members of the Six protein family. Moreover, the Hypoxia Inducible Factor-1α (HIF-1α) has been identified as a master regulator for the expression of genes involved in essential hypoxic responses that maintain ATP levels in cells. Ablation of HIF-1α in skeletal muscle was associated with an increase in the activity of bob-limiting enzymes of the mitochondria, indicating that the citric acid cycle and increased fatty acid oxidation may be compensating for decreased flow through the glycolytic pathway in these animals. However, hypoxia-mediated HIF-1α responses are also linked to the regulation of mitochondrial dysfunction through the formation of excessive reactive oxygen species in mitochondria.
Other pathways also influence adult muscle character. For example, physical force inside a muscle fiber may release the transcription factor Serum Response Factor (SRF) from the structural protein titin, leading to altered muscle growth.
Research
Research on skeletal muscle properties uses many techniques. Electrical muscle stimulation is used to determine force and contraction speed at different stimulation frequencies, which are related to fiber-type composition and mix within an individual muscle group. In vitro muscle testing is used for more complete characterization of muscle properties.
See also
- Hill's muscle model
- Myopathy
- Muscle atrophy
- Musculoskeletal injury
References
- ^ Scott, Wayne; Stevens, Jennifer; Binder–Macleod, Stuart A. (2001). "Human Skeletal Muscle Fiber Type Classifications". Physical Therapy 81 (11): 1810–1816. PMID 11694174. http://ptjournal.apta.org/content/81/11/1810.full.
- ^ Saladin, Kenneth S. (2010). Anatomy and Physiology (5nd ed.). New York: Watnick. pp. 405–406. ISBN 978-0-07-727620-1.
- ^ a b c d e f g h i j k Martini, Frederic H.; Timmons, Michael J.; Tallitsch, Robert B. (2008). Human Anatomy (6 ed.). Benjamin Cummings. pp. 251–252. ISBN 9780321500427.
- ^ Costanzo, Linda S. (2002). Physiology (2nd ed.). Philadelphia: Saunders. pp. 23. ISBN 0-7216-9549-3.
- ^ Quoted from National Skeletal Muscle Research Center; UCSD, Muscle Physiology Home Page - Skeletal Muscle Architecture, Effect of Muscle Architecture on Muscle Function
- ^ Barry, D. T. (1992). "Vibrations and sounds from evoked muscle twitches". Electromyogr Clin Neurophysiol. 32 (1–2): 35–40. PMID 1541245.
- ^ [1], Peak Performance - Endurance training: understanding your slow twitch muscle fibres will boost performance
Muscular system (TA A04.0, GA 4.361) Topics Types of muscles Other Unipennate muscle · Bipennate muscle · Origin · Insertion · Fascia (Superficial fascia, Deep fascia, Visceral fascia) · Tendon/Aponeurosis · Fascial compartmentHistology: muscle tissue (TH H2.00.05, H3.3) Smooth
muscleStriated
muscleSkeletal
muscleCostamere/
DAPCMembrane/
extracellularIntracellularDystrophin · Dystrobrevin (A, B) · Syntrophin (A, B1, B2, G1, G2) · Syncoilin · Dysbindin · Synemin/desmuslin
related: NOS1 · Caveolin 3GeneralNeuromuscular junction · Motor unit · Muscle spindle · Excitation-contraction coupling · Sliding filament mechanismBothFiberCellsOtherOther/
ungroupedCategories:- Muscular system
- Somatic motor system

Wikimedia Foundation. 2010.