- Phylogenetic comparative methods
-
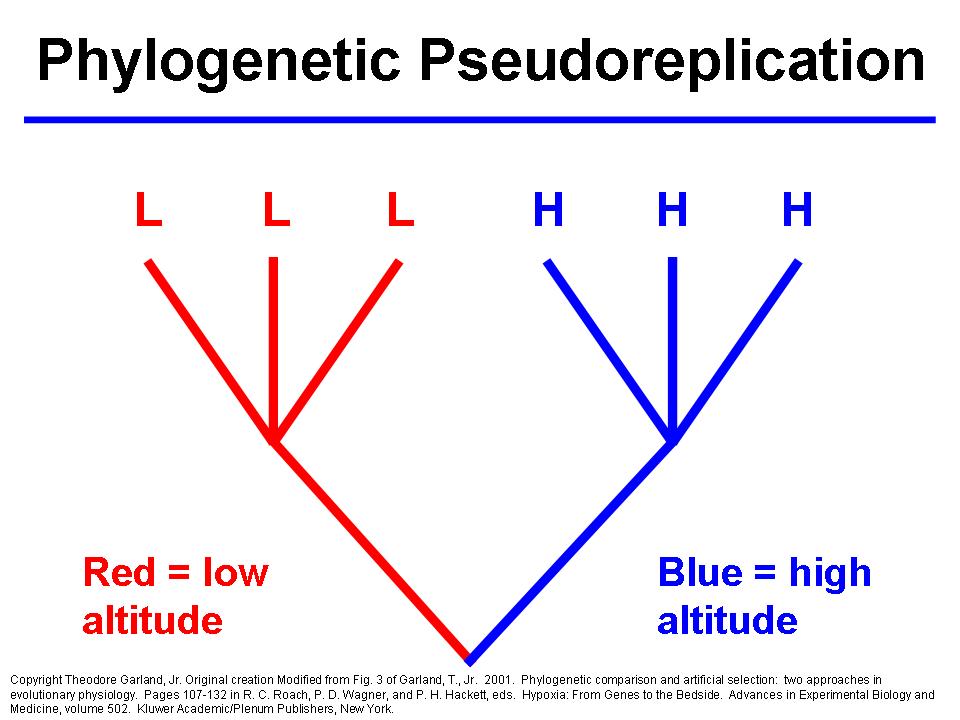
 When applied to comparative data, conventional statistical methods assume, in effect, that all species are completely unrelated. As if they descended from a "big bang" of special creation. Such a scenario can be depicted as a "star" phylogeny (left). Most comparative studies will involve species that have descended in a hierarchical fashion from common ancestors, as shown on the right. Comparative data sets may include species that have gone extinct, and rates of evolution may vary among branches. Therefore, trees used in phylogenetic comparative methods may not be "ultrametric" (have tips that are contemporaneous).
When applied to comparative data, conventional statistical methods assume, in effect, that all species are completely unrelated. As if they descended from a "big bang" of special creation. Such a scenario can be depicted as a "star" phylogeny (left). Most comparative studies will involve species that have descended in a hierarchical fashion from common ancestors, as shown on the right. Comparative data sets may include species that have gone extinct, and rates of evolution may vary among branches. Therefore, trees used in phylogenetic comparative methods may not be "ultrametric" (have tips that are contemporaneous).
Phylogenetic comparative methods (PCMs) use information on the evolutionary relationships of organisms (phylogenetic trees) to compare species (Harvey and Pagel, 1991). The most common applications are to test for correlated evolutionary changes in two or more traits, or to determine whether a trait contains a phylogenetic signal (the tendency for related species to resemble each other). However, several methods are available to relate particular phenotypic traits to variation in rates of speciation and/or extinction, including attempts to identify evolutionary key innovations. Although most studies that employ PCMs focus on extant organisms, the methods can also be applied to extinct taxa and can incorporate information from the fossil record.
Owing to their computational requirements, they are usually implemented by computer programs (see list below). PCMs can be viewed as part of evolutionary biology, systematics, phylogenetics, bioinformatics or even statistics, as most methods involve statistical procedures and principles for estimation of various parameters and drawing inferences about evolutionary processes.
What distinguishes PCMs from most traditional approaches in systematics and phylogenetics is that they typically do not attempt to infer the phylogenetic relationships of the species under study. Rather, they use an independent estimate of the phylogenetic tree (topology plus branch lengths) that is derived from a separate phylogenetic analysis, such as comparative DNA sequences that have been analyzed by maximum parsimony or maximum likelihood methods. PCMs are consumers of phylogenetic trees, not primary producers of them. Accordingly, the list of phylogenetics software shows little overlap with the programs for PCMs (see below).
Comparison of species to elucidate aspects of biology has a long history. Charles Darwin relied on such comparisons as a major source of evidence when writing The Origin of Species. Many other fields of biology use interspecific comparison as well, including behavioral ecology, ethology, ecophysiology, comparative physiology, evolutionary physiology, functional morphology, comparative biomechanics, and the study of sexual selection.
The comparative method is also used heavily in linguistics.
Contents
Applications
PCMs can be used to analyze the origin and maintenance of biodiversity. Biodiversity is most commonly discussed in terms of the number of species, but it can also be phrased in terms of the amount of phenotypic (e.g., physiological, morphological) space that a given set of species occupies (see also Cambrian explosion).
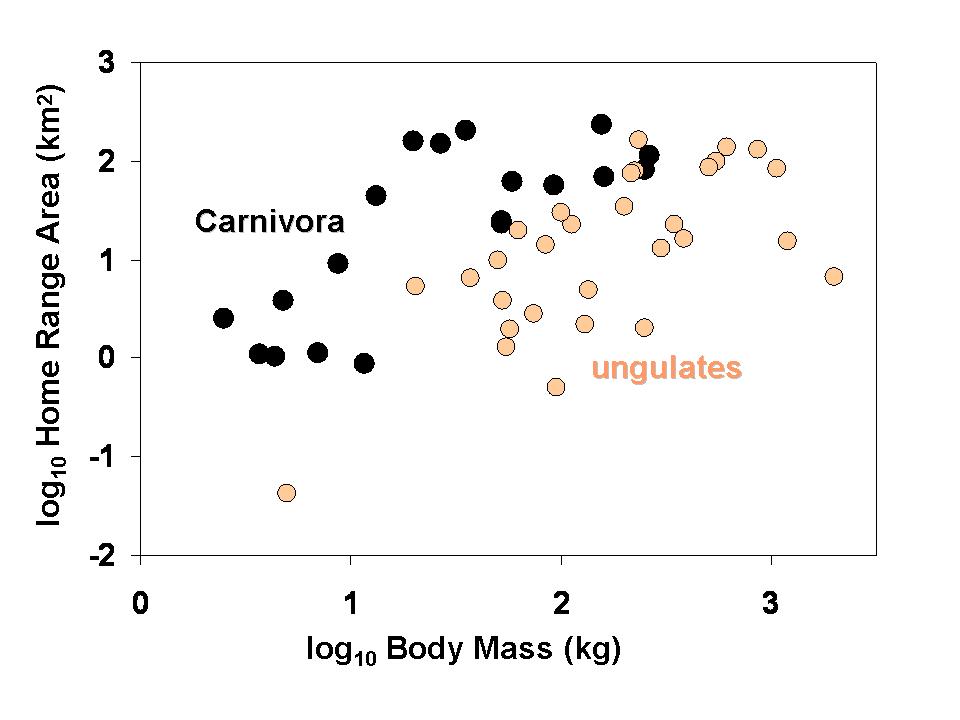
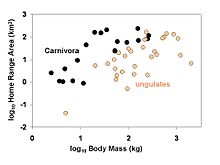 Home range areas of 49 species of mammals in relation to their body size. Larger-bodied species tend to have larger home ranges, but at any given body size members of the order Carnivora (carnivores and omnivores) tend to have larger horme ranges than ungulates (all of which are herbivores). Whether this difference is considered statistically significant depends on what type of analysis is applied (Garland et al., 1993).
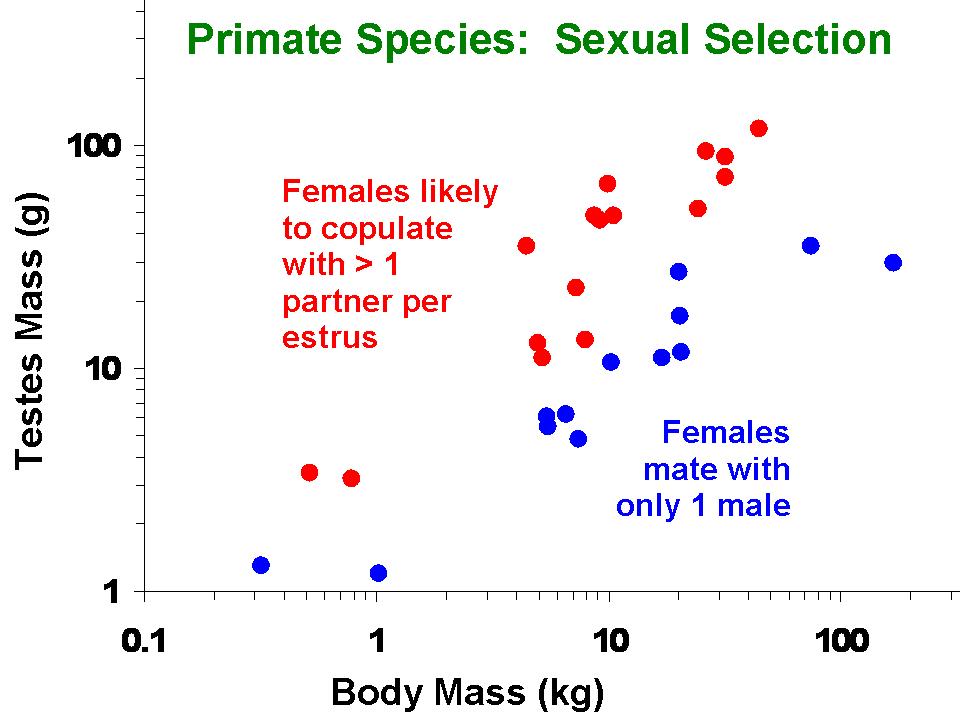
Home range areas of 49 species of mammals in relation to their body size. Larger-bodied species tend to have larger home ranges, but at any given body size members of the order Carnivora (carnivores and omnivores) tend to have larger horme ranges than ungulates (all of which are herbivores). Whether this difference is considered statistically significant depends on what type of analysis is applied (Garland et al., 1993). Testes mass of various species of Primates in relation to their body size and mating system. Larger-bodied species tend to have larger testes, but at any given body size species in which females tend to mate with multiple males have males with larger testes.
Testes mass of various species of Primates in relation to their body size and mating system. Larger-bodied species tend to have larger testes, but at any given body size species in which females tend to mate with multiple males have males with larger testes.Phylogenetic comparative methods are commonly applied to such questions as:
- What is the slope of an allometric scaling relationship?
→ Example: how does brain mass vary in relation to body mass?
- Do different clades of organisms differ with respect to some phenotypic trait?
→ Example: do canids have larger hearts than felids?
- Do groups of species that share a behavioral or ecological feature (e.g., social system, diet) differ in average phenotype?
→ Example: do carnivores have larger home ranges than herbivores?
- What was the ancestral state of a trait?
→ Example: where did endothermy evolve in the lineage that led to mammals?
→ Example: where, when, and why did placentas and viviparity evolve?
- Does a trait exhibit significant phylogenetic signal in a particular group of organisms? Do certain types of traits tend to "follow phylogeny" more than others?
→ Example: are behavioral traits more labile during evolution?
- Do species differences in life history traits trade-off, as in the so-called fast-slow continuum?
→ Example: why do small-bodied species have shorter life spans than their larger relatives?
Phylogenetically independent contrasts
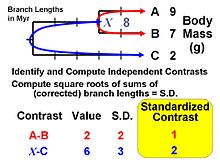 The standardized contrasts are used in conventional statistical procedures, with the constraint that all regressions, correlations, analysis of covariance, etc., must pass through the origin.
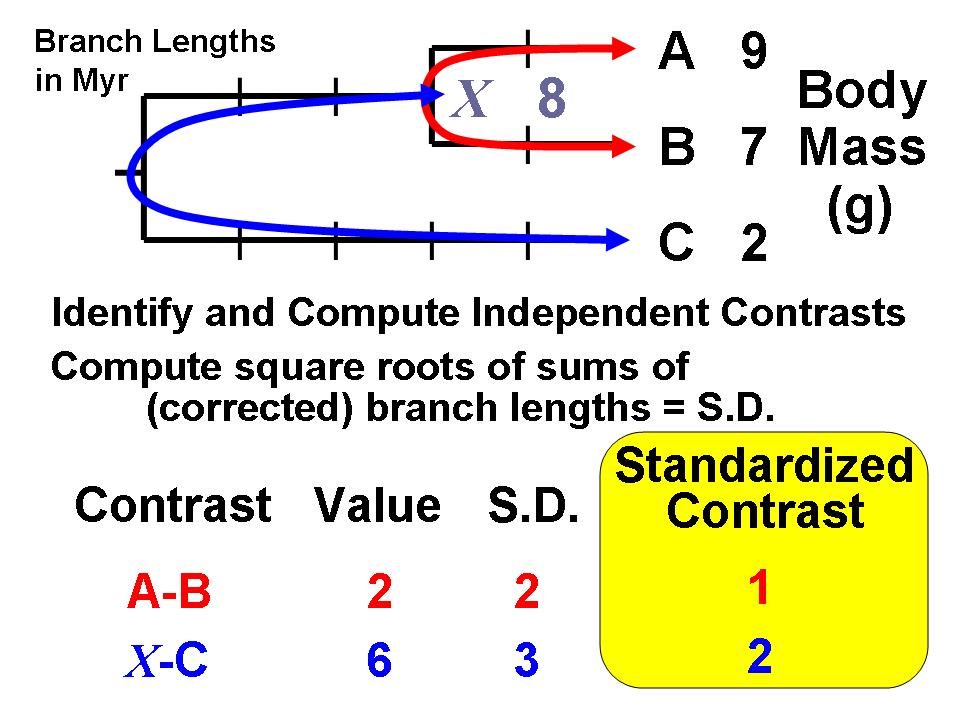
The standardized contrasts are used in conventional statistical procedures, with the constraint that all regressions, correlations, analysis of covariance, etc., must pass through the origin.Felsenstein (1985) proposed the first general statistical method for incorporating phylogenetic information, i.e., the first that could use any arbitrary topology (branching order) and a specified set of branch lengths. The method is now recognized as an algorithm that implements a special case of what are termed phylogenetic generalized least-squares models (Grafen, 1989). The logic of the method is to use phylogenetic information (and an assumed Brownian motion like model of trait evolution) to transform the original tip data (mean values for a set of species) into values that are statistically independent and identically distributed.
The algorithm involves computing values at internal nodes as an intermediate step, but they are generally not used for inferences by themselves. An exception occurs for the basal (root) node, which can be interpreted as an estimate of the ancestral value for the entire tree (assuming that no directional evolutionary trends [e.g., Cope's rule] have occurred) or as a phylogenetically weighted estimate of the mean for the entire set of tip species (terminal taxa). The value at the root is equivalent to that obtained from the "squared-change parsimony" algorithm and is also the maximum likelihood estimate under Brownian motion. The independent contrasts algebra can also be used to compute a standard error or confidence interval.
Phylogenetically informed Monte Carlo computer simulations
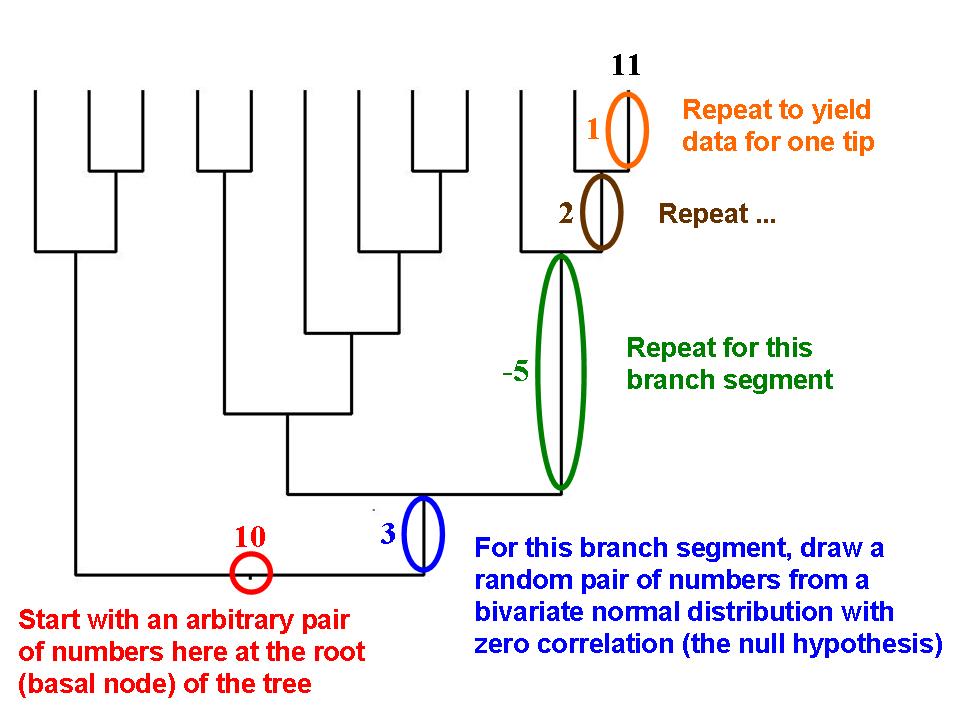
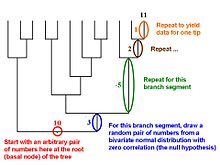 Data for a continuous-valued trait can be simulated in such a way that taxa at the tips of a hypothetical phylogenetic tree will exhibit phylogenetic signal, i.e., closely related species will tend to resemble each other.
Data for a continuous-valued trait can be simulated in such a way that taxa at the tips of a hypothetical phylogenetic tree will exhibit phylogenetic signal, i.e., closely related species will tend to resemble each other.Martins and Garland (1991) proposed that one way to account for phylogenetic relations when conducting statistical analyses was to use computer simulations to create many data sets that are consistent with the null hypothesis under test (e.g., no correlation between two traits, no difference between two ecologically defined groups of species) but that mimic evolution along the relevant phylogenetic tree. If such data sets (typically 1,000 or more) are analyzed with the same statistical procedure that is used to analyze a real data set, then results for the simulated data sets can be used to create phylogenetically correct (or "PC" [Garland et al., 1993]) null distributions of the test statistic (e.g., a correlation coefficient, t, F). Such simulation approaches can also be combined with methods like phylogenetically independent contrasts (see above).
Related quotations
In addition to having a long and extremely productive history in biology, comparative methods are often controversial. Rather than attempt to recount the various controversies, many of which are ongoing, quotes (in chronological order) are illustrative.
"Ought we, for instance, to begin by discussing each separate species - man, lion, ox, and the like - taking each kind in hand independently of the rest, or ought we rather to deal first with the attributes which they have in common in virtue of some common element of their nature, and proceed from this as a basis for the consideration of the separately?" (Aristotle, De partibus animalium; quoted in Harvey and Pagel, 1991, p. 35)
"In parallel with laboratory experimental methods, the comparative method increases in value with its sample size, i.e., the number of species being compared. When species are few and their phylogenetic relationships are clouded in the distant past of unfossilized ancestors, the comparative method reaps fewer conclusions of trust." (Hailman, 1976, p. 20)
"... there is no easy way, except by comparison, to test most questions about the long-term history of life, or to generate predictions from evolutionary considerations." (Alexander, 1979, p. 13)
"... we must learn to treat comparative data with the same respect as we would treat experimental results ..." (Maynard Smith and Holliday, 1979, p. vii)
"In the past, however, cooperation between systematists and other comparative biologists has been sporadic at best. Most experimental biologists have ignored taxonomy and systematics, some even to the extent of not bothering to provide their animals with proper identifications or scientific names." (Atz et al., 1980, p. 7)
"If we assume that the ... cladogram .. is correct, we can then hypothesize what the particular common ancestor must have been like." (Atz et al., 1980, p. 14)
"... biology will never secure fossils of all species in the past because fossilization is such a rare process, requiring just the right physical and chemical conditions. Therefore, in order to trace probable phylogenetic lineages one must reason from the evidence at hand: the characteristics of contemporary animals themselves which are the end-points of phylogeny (evolutionary history)." (Hailman, 1981, p. 93)
"... the comparative method of 1950 was indistinguishable from the comparative method of 350 BC ..." (Ridley, 1983, p. 6)
"Focusing only on highly adapted species may, of course, bring valuable information on extreme situations but might also obscure the basic mechanisms." (Bankir and Rouffignac, 1985, p. R663)
"Most of what we know is based upon comparison. When asked to describe a food not previously tasted or a new kind of music, one often responds that the taste is "like" some other food, or that the sensation "differs" in a particular way from something that is familiar. Indeed, comparison and the similarities and differences it discloses is ingrained in our approach to description of objects, events and processes. Hence the questions "what can we compare?" and its ancillary "how shall we compare?" prove to be the key to any study of natural phenomena." (Gans, 1985, p. 291)
"Some reviewers of this paper felt that the message was "rather nihilistic," and suggested that it would be much improved if I could present a simple and robust method that obviated the need to have an accurate knowledge of phylogeny. I entirely sympathize, but do not have a method that solves the problem. The best we can do is perhaps to use pairs of close relatives as suggested above, although this discards at least half of the data. Comparative biologists may understandably feel frustrated upon being told that they need to know the phylogenies of their groups in great detail, when this is not something they had much interest in knowing. Nevertheless, efforts to cope with the effects of the phylogeny will have to be made. Phylogenies are fundamental to comparative biology; there is no doing it without taking them into account." (Felsenstein, 1985, p. 14)
"Comparative biologists tend to suspect comparisons of distantly related species; they hope to base their comparisons on recent evolutionary events that have not been overlaid by much subsequent change." (Felsenstein, 1988, p. 465)
"Yet, one of the most embarrassing things that could be done to a group of respected biologists would be to ask them to spend a few minutes to write down what is meant by the comparative method, and what are the basic goals and principles of biological comparison." (Bock, 1989, p. 18 [cited in Starck, 1998, p. 110])
"As welcome as it is to see the lung successfully employed as a systematic index, it is, on the other hand, unfortunate. Thereby, lungs lose their innocence in the sense that phylogenetic trees and cladograms can no longer be used to help resolve the sequence in the development of lung structure without the danger of circular argumentation." (Perry, 1989, p. 200)
"To be maximally informative, such studies should be undertaken on closely related groups of organisms, so that factors extraneous to the comparison can be minimized ..." (Bennett and Huey, 1990, p. 272)
"The comparative approach is not new. Indeed, it was Darwin's favoured technique. ... In short, comparative studies have taught us most of what we know about adaptation." (Harvey and Pagel, 1991, p. v)
"In short, all useful comparative methods are based on explicit models of evolutionary change." (Harvey and Pagel, 1991, p. v)
"Adaptation is an inherently comparative idea ..." (Harvey and Pagel, 1991, p. 13)
"The usual symptom of non-independence is that closely related species tend to be more alike than more distantly related species." (Harvey and Pagel, 1991, p. 81)
"... to use species as independent data points in a comparative analysis requires that one ignores phylogenetic relationships." (Harvey and Pagel, 1991, p. 122)
"Because life-history traits are likely to be correlated with a species' phylogenetic history, unequivocal evidence for adaptation to local environmental conditions may be recognized only after the variation in a trait attributable to phylogeny is removed." (Miles and Dunham, 1992, p. 848)
"Broad-scale comparative evidence from across a large number of taxa may often help set limits to the applicability of hypotheses that have been generated from a particular phenomenon in a particular species." (Moller and Birkhead, 1992, p. 650)
"Any comparative study lacking a phylogenetic perspective now would be incomplete." (Miles and Dunham, 1993, p. 590)
"However, in general, the evolutionary process involves descent with modification, and in the absence of modification, one must conclude that similarities among closely related taxa reflect shared ancestry. Phylogeny, then, is an important explanatory principle for understanding shared characteristics and should be the null hypothesis in all tests of similarity or differentiation among taxa." (Di Fiore and Rendall, 1994, p. 9945)
"Moreover, comparative studies supply only correlational data. Correlation does not demonstrate causation ..." (Garland and Adolph, 1994, p. 823)
"In addition to his theory of natural selection, the comparative method is what made Darwin great. If you don't believe this claim, look at any of his major works. They are packed full with interspecific comparisons based on detailed studies and anecdotal observations." (Hauser, 1996, p. 10)
"Population comparisons can provide particularly powerful means of evaluating adaptive hypotheses for two reasons. The first is that there tend to be fewer differences between populations than between species. Consequently, there are fewer covarying traits to confound analyses ... Second, divergent populations are often relatively young and may be more likely to reside in the habitats in which their derived character states evolved than is the case for divergent species with potentially longer intervening histories ..." (Foster and Cameron, 1996, p. 140).
"Naive, prephylogenetic comparative tests should be kept at the other end of a barge pole." (Ridley and Grafen, 1996, p. 87)
"It is the study of the bizarre, the outliers, the freaks, that gives us some of our clearest insights into the hows and whys of evolution." (Torr, 1998, p. 52)
See also
- Allometry
- Behavioral ecology
- Biodiversity
- Bioinformatics
- Cladistics
- Comparative anatomy
- Comparative method in linguistics
- Comparative physiology
- Computational phylogenetics
- Ecophysiology
- Evolutionary neurobiology
- Evolutionary physiology
- Generalized linear model
- Joe Felsenstein
- Maximum likelihood
- Maximum parsimony
- Phylogenetics
- Sexual selection
- Statistics
- Systematics
- Theodore Garland, Jr.
References
- Felsenstein, J. 1985. Phylogenies and the comparative method. American Naturalist 125:1-15.
- Garland, T., Jr., A. W. Dickerman, C. M. Janis, and J. A. Jones. 1993. Phylogenetic analysis of covariance by computer simulation. Systematic Biology 42:265-292. PDF
- Grafen, A. 1989. The phylogenetic regression. Philosophical Transactions of the Royal Society of London B 326:119-157. PDF
- Harvey, P. H., and M. D. Pagel. 1991. The comparative method in evolutionary biology. Oxford University Press, Oxford. 239 pp.
- Martins, E. P., and T. Garland, Jr.. 1991. Phylogenetic analyses of the correlated evolution of continuous characters: a simulation study. Evolution 45:534-557. PDF
Further reading
- Ackerly, D. D. 1999. Comparative plant ecology and the role of phylogenetic information. Pages 391-413 in M. C. Press, J. D. Scholes, and M. G. Braker, eds. Physiological plant ecology. The 39th symposium of the British Ecological Society held at the University of York 7-9 September 1998. Blackwell Science, Oxford, U.K.
- Berenbrink, M., P. Koldkjær, O. Kepp, and A. R. Cossins. 2005. Evolution of oxygen secretion in fishes and the emergence of a complex physiological system. Science 307:1752-1757.
- Blomberg, S. P., T. Garland, Jr., and A. R. Ives. 2003. Testing for phylogenetic signal in comparative data: behavioral traits are more labile. Evolution 57:717-745. PDF
- Brooks, D. R., and D. A. McLennan. 1991. Phylogeny, ecology, and behavior: a research program in comparative biology. Univ. Chicago Press, Chicago. 434 pp.
- Cheverud, J. M., M. M. Dow, and W. Leutenegger. 1985. The quantitative assessment of phylogenetic constraints in comparative analyses: sexual dimorphism in body weight among primates. Evolution 39:1335-1351.
- Eggleton, P., and R. I. Vane-Wright, eds. 1994. Phylogenetics and ecology. Linnean Society Symposium Series Number 17. Academic Press, London.
- Felsenstein, J. 2004. Inferring phylogenies. Sinauer Associates, Sunderland, Mass. xx + 664 pp.
- Freckleton, R. P., P. H. Harvey, and M. Pagel. 2002. Phylogenetic analysis and comparative data: a test and review of evidence. American Naturalist 160:712-726.
- Garland, T., Jr., and A. R. Ives. 2000. Using the past to predict the present: Confidence intervals for regression equations in phylogenetic comparative methods. American Naturalist 155:346-364. PDF
- Garland, T., Jr., Jr., A. F. Bennett, and E. L. Rezende. 2005. Phylogenetic approaches in comparative physiology. Journal of Experimental Biology 208:3015-3035. PDF
- Garland, T., Jr., P. H. Harvey, and A. R. Ives. 1992. Procedures for the analysis of comparative data using phylogenetically independent contrasts. Systematic Biology 41:18-32. PDF
- Gittleman, J. L., and M. Kot. 1990. Adaptation: statistics and a null model for estimating phylogenetic effects. Systematic Zoology 39:227-241.
- Herrada, E. A., C. J. Tessone, K. Klemm, V. M. Eguiluz, E. Hernandez-Garcia, and C. M. Duarte. 2008. Universal Scaling in the Branching of the Tree of Life. PLoS One 3(7): e2757. doi:10.1371/journal.pone.0002757. PDF
- Housworth, E. A., E. P. Martins, and M. Lynch. 2004. The phylogenetic mixed model. American Naturalist 163:84-96. PDF
- Ives, A. R., P. E. Midford, and T. Garland, Jr. 2007. Within-species variation and measurement error in phylogenetic comparative methods. Systematic Biology 56:252-270.
- Maddison, D. R. 1994. Phylogenetic methods for inferring the evolutionary history and process of change in discretely valued characters. Annual Review of Entomology 39:267-292.
- Maddison, W. P. 1990. A method for testing the correlated evolution of two binary characters: Are gains or losses concentrated on certain branches of a phylogenetic tree? Evolution 44:539-557.
- Maddison, W. P., and D. R. Maddison. 1992. MacClade. Analysis of phylogeny and character evolution. Version 3. Sinauer Associates, Sunderland, Mass. 398 pp.
- Martins, E. P., ed. 1996. Phylogenies and the comparative method in animal behavior. Oxford University Press, Oxford. 415 pp.
- Martins, E. P., and T. F. Hansen. 1997. Phylogenies and the comparative method: a general approach to incorporating phylogenetic information into the analysis of interspecific data. American Naturalist 149:646-667. Erratum Am. Nat. 153:448.
- Nunn, C. L., and R. A. Barton. 2001. Comparative methods for studying primate adaptation and allometry. Evolutionary Anthropology 10:81-98.
- Oakley, T. H., Z. Gu, E. Abouheif, N. H. Patel, and W.-H. Li. 2005. Comparative methods for the analysis of gene-expression evolution: an example using yeast functional genomic data. Molecular Biology and Evolution 22:40-50. PDF
- O’Meara, B. C., C. M. Ané, M. J. Sanderson, and P. C. Wainwright. 2006. Testing for different rates of continuous trait evolution in different groups using likelihood. Evolution 60:922-933. PDF
- Organ, C. L., A. M. Shedlock, A. Meade, M.. Pagel, and S. V. Edwards. 2007. Origin of avian genome size and structure in non-avian dinosaurs. Nature 446:180-184.
- Page, R. D. M., ed. 2003. Tangled trees : phylogeny, cospeciation, and coevolution. University of Chicago Press, Chicago.
- Pagel, M. D. 1993. Seeking the evolutionary regression coefficient: an analysis of what comparative methods measure. Journal of Theoretical Biology 164:191-205.
- Pagel, M. 1999. Inferring the historical patterns of biological evolution. Nature 401:877-884.
- Paradis, E. 2005. Statistical analysis of diversification with species traits. Evolution 59:1-12.
- Paradis, E., and J. Claude. 2002. Analysis of comparative data using generalized estimating equations. Journal of Theoretical Biology 218:175-185.
- Purvis, A., and T. Garland, Jr. 1993. Polytomies in comparative analyses of continuous characters. Systematic Biology 42:569-575. PDF
- Rezende, E. L., and T. Garland, Jr. 2003. Comparaciones interespecíficas y métodos estadísticos filogenéticos. Pages 79-98 in F. Bozinovic, ed. Fisiología Ecológica & Evolutiva. Teoría y casos de estudios en animales. Ediciones Universidad Católica de Chile, Santiago. PDF
- Ridley, M. 1983. The explanation of organic diversity: The comparative method and adaptations for mating. Clarendon, Oxford, U.K.
- Rohlf, F. J. 2001. Comparative methods for the analysis of continuous variables: geometric interpretations. Evolution 55:2143-2160.
- Rohlf, F. J. 2006. A comment on phylogenetic correction. Evolution 60:1509-1515.
- Sanford, G. M., W. I. Lutterschmidt, and V. H. Hutchison. 2002. The comparative method revisited. BioScience 52:830-836.
- Schluter, D., T. Price, A. O. Mooers, and D. Ludwig. 1997. Likelihood of ancestor states in adaptive radiation. Evolution 51:1699-1711.
- Smith, R. J., and J. M. Cheverud. 2002. Scaling of sexual size dimorphism in body mass: a phylogenetic analysis of Rensch's rule in primates. International Journal of Primatology 23:1095-1135.
- Steppan, S. J., P. C. Phillips, and D. Houle. 2002. Comparative quantitative genetics: evolution of the G matrix. Trends in Ecology and Evolution 17:320-327. PDF
- Vanhooydonck, B., and R. Van Damme. 1999. Evolutionary relationships between body shape and habitat use in lacertid lizards. Evolutionary Ecology Research 1:785-805.
External links
Journals
- American Naturalist
- Behavioral Ecology
- Ecology
- Evolution
- Evolutionary Ecology Research
- Functional Ecology
- Journal of Evolutionary Biology
- Philosophical Transactions of the Royal Society of London B
- Physiological and Biochemical Zoology
- Systematic Biology
Software packages (incomplete list)
- Analyses of Phylogenetics and Evolution
- Comparative Analysis by Independent Contrasts
- COMPARE
- Felsenstein's List
- Mesquite PDAP:PDTree for Mesquite
- Ornstein-Uhlenbeck for Comparative Hypotheses
- Phenotypic Diversity Analysis Programs
- Phylocom
- Phylogenetic Regression
- PHYSIG
- BayesTraits
Laboratories
Branches of Biology Anatomy · Astrobiology · Biochemistry · Biogeography · Biomechanics · Biophysics · Bioinformatics · Biostatistics · Botany · Cell biology · Cellular microbiology · Chemical biology · Chronobiology · Conservation biology · Developmental biology · Ecology · Epidemiology · Epigenetics · Evolutionary biology · Genetics · Genomics · Histology · Human biology · Immunology · Marine biology · Mathematical biology · Microbiology · Molecular biology · Mycology · Neuroscience · Nutrition · Origin of life · Paleontology · Parasitology · Pathology · Pharmacology · Physiology · Quantum biology · Systematics · Systems biology · Taxonomy · Toxicology · ZoologyCategories:

Wikimedia Foundation. 2010.