- Programmed cell death
-
Programmed cell-death (or PCD) is death of a cell in any form, mediated by an intracellular program.[1][2] PCD is carried out in a regulated process which generally confers advantage during an organism's life-cycle. PCD serves fundamental functions during both plant and metazoa (multicellular animals) tissue development.
Contents
Types
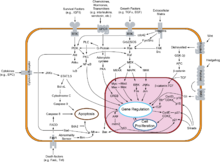 Overview of signal transduction pathways involved in apoptosis.
Overview of signal transduction pathways involved in apoptosis.
- Apoptosis or Type I cell-death.
- Autophagic or Type II cell-death. (Cytoplasmic: characterized by the formation of large vacuoles which eat away organelles in a specific sequence prior to the nucleus being destroyed.)[3]
- Necrotic cell death or Type-III[4]
Besides these three types of PCD, other pathways have been discovered.[5] Called "non-apoptotic programmed cell-death" (or "caspase-independent programmed cell-death" or "necrosis-like programmed cell-death"), these alternative routes to death are as efficient as apoptosis and can function as either backup mechanisms or the main type of PCD.
Other forms of programmed cell death include anoikis, almost identical to apoptosis except in its induction; cornification, a form of cell death exclusive to the eyes; excitotoxicity and Wallerian degeneration.
Plant cells undergo particular processes of PCD which are similar to autophagic cell death. However, some common features of PCD are highly conserved in both plants and metazoa.
Atrophic factors
An atrophic factor is a force that causes a cell to die. Only natural forces on the cell are considered to be atrophic factors.[by whom?] The following are generally considered[weasel words] to be common types of atrophic factors;[6]
- Decreased workload
- Loss of innervation
- Diminished blood supply
- Inadequate nutrition
- Loss of endocrine stimulation
- Senility
- Compression
History
The concept of "programmed cell-death" was used by Lockshin & Williams[7] in 1964 in relation to insect tissue development, around eight years before "apoptosis" was coined. Since then, PCD has become the more general of these two terms.
PCD has been the subject of increasing attention and research efforts. This trend has been highlighted with the award of the 2002 Nobel Prize in Physiology or Medicine to Sydney Brenner (United Kingdom), H. Robert Horvitz (US) and John E. Sulston (UK).[8]
Programmed cell-death in plant tissue
Programmed cell death in plants has a number of molecular similarities to animal apoptosis, but it also has differences, most obviously the presence of a cell wall and the lack of an immune system which removes the pieces of the dead cell. Instead of an immune response, the dying cell synthesizes substances to break itself down and places them in a vacuole which ruptures as the cell dies.[9]
In "APL regulates vascular tissue identity in Arabidopsis",[10] Bonke and colleagues state that one of the two long-distance transport systems in vascular plants, xylem, consists of several cell-types "the differentiation of which involves deposition of elaborate cell-wall thickenings and programmed cell-death." The authors emphasize that the products of plant PCD play an important structural role.
Basic morphological and biochemical features of PCD have been conserved in both plant and animal kingdoms.[11] It should be noted, however, that specific types of plant cells carry out unique cell-death programs. These have common features with animal apoptosis—for instance, nuclear DNA degradation—but they also have their own peculiarities, such as nuclear degradation being triggered by the collapse of the vacuole in tracheary elements of the xylem.[12]
Janneke Balk and Christopher J. Leaver, of the Department of Plant Sciences, University of Oxford, carried out research on mutations in the mitochondrial genome of sun-flower cells. Results of this research suggest that mitochondria play the same key role in vascular plant PCD as in other eukaryotic cells.[13]
PCD in pollen prevents inbreeding
During pollination, plants enforce self-incompatibility (SI) as an important means to prevent self-fertilization. Research on the corn poppy (Papaver rhoeas) has revealed that proteins in the pistil on which the pollen lands, interact with pollen and trigger PCD in incompatible (ie. self) pollen. The researchers, Steven G. Thomas and Veronica E. Franklin-Tong, also found that the response involves rapid inhibition of pollen-tube growth, followed by PCD.[14]
Programmed cell death in slime molds
The social slime mold Dictyostelium discoideum has the peculiarity of adopting either a predatory amoeba-like behavior in its unicellular form, or coalescing into a mobile slug-like form when dispersing the spores which will give birth to the next generation.[15]
The stalk is composed of dead cells which have undergone a type of PCD that shares many features of an autophagic cell-death: massive vacuoles forming inside cells, a degree of chromatin condensation, but no DNA fragmentation.[16] The structural role of the residues left by the dead cells is reminiscent of the products of PCD in plant tissue.
D. discoideum is a slime mold, part of a branch which may have emerged from eukaryotic ancestors about a billion years before the present. They apparently emerged after the ancestors of green plants and the ancestors of fungi and animals had differentiated. But in addition to their place in the evolutionary tree, the fact that PCD has been observed in the humble, simple, six-chromosome D. discoideum has additional significance: it permits the study of a developmental PCD path which does not depend on the caspases which are characteristic of apoptosis.[17]
Evolutionary origin of PCD
Biologists had long suspected that mitochondria originated from bacteria which had been incorporated as endosymbionts ("living together inside") of larger eukaryotic cells. It was Lynn Margulis who from 1967 on championed this theory, which has since become widely accepted.[18] The most convincing evidence for this theory is the fact that mitochondria possess their own DNA and are equipped with genes and replication apparatus.
This evolutionary step would have been more than risky for the primitive eukaryotic cells which began to engulf the energy-producing bacteria and conversely, a perilous step for the ancestors of mitochondria which began to invade their proto-eukaryotic hosts. This process is still evident today, between human white blood cells and bacteria. Most of the time, invading bacteria are destroyed by the white blood cells; however, it is not uncommon for the chemical warfare waged by prokaryotes to succeed, with the consequence known as infection by its resulting damage.
One of these rare evolutionary events, about two billion years before the present, made it possible for certain eukaryotes and energy-producing prokaryotes not only to coexist, but to mutually benefit from their symbiosis.[19]
Mitochondriate eukaryotic cells live poised between life and death, because mitochondria still retain their repertoire of molecules which can trigger cell suicide.[20] This process has now been evolved to happen only when programmed.[citation needed] Given certain signals to cells (such as feedback from neighbors, stress or DNA damage), mitochondria release caspase activators which trigger the cell-death inducing biochemical cascade. As such, the cell suicide mechanism is now crucial to all of our lives.
Programmed death of entire organisms
Main article: PhenoptosisReferences
- ^ Engelberg-Kulka H, Amitai S, Kolodkin-Gal I, Hazan R (2006). "Bacterial Programmed Cell Death and Multicellular Behavior in Bacteria". PLoS Genetics 2 (10): e135. doi:10.1371/journal.pgen.0020135. PMC 1626106. PMID 17069462. http://genetics.plosjournals.org/perlserv/?request=get-document&doi=10.1371/journal.pgen.0020135.
- ^ Green, Douglas (2011). Means To An End. New York: Cold Spring Harbor Laboratory Press. ISBN 978-0-87969-888-1. http://celldeathbook.wordpress.com/.
- ^ Schwartz LM, Smith SW, Jones ME, Osborne BA (1993). "Do all programmed cell deaths occur via apoptosis?". PNAS 90 (3): 980–4. doi:10.1073/pnas.90.3.980. PMC 45794. PMID 8430112. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=45794.;and, for a more recent view, see Bursch W, Ellinger A, Gerner C, Fröhwein U, Schulte-Hermann R (2000). "Programmed cell death (PCD). Apoptosis, autophagic PCD, or others?". Annals of the New York Academy of Sciences 926: 1–12. doi:10.1111/j.1749-6632.2000.tb05594.x. PMID 11193023.
- ^ Smith CC, Yellon DM (2011). "Necroptosis, necrostatins and tissue injury". J Cell Mol Med 15 (9): 1797–806. doi:10.1111/j.1582-4934.2011.01341.x. PMID 21564515.
- ^ Kroemer G, Martin SJ (2005). "Caspase-independent cell death". Nature Medicine 11 (7): 725–30. doi:10.1038/nm1263. PMID 16015365.
- ^ Chapter 10: All the Players on One Stage from PsychEducation.org
- ^ Lockshin RA, Williams CM (1964). "Programmed cell death—II. Endocrine potentiation of the breakdown of the intersegmental muscles of silkmoths". Journal of Insect Physiology 10 (4): 643–649. doi:10.1016/0022-1910(64)90034-4. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6T3F-49N9JTD-BN&_user=10&_coverDate=08%2F31%2F1964&_alid=509668351&_rdoc=1&_fmt=summary&_orig=search&_cdi=4945&_sort=d&_docanchor=&view=c&_acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=244d74b599c3c6e6979401175da87303.
- ^ "The Nobel Prize in Physiology or Medicine 2002". The Nobel Foundation. 2002. http://nobelprize.org/nobel_prizes/medicine/laureates/2002/index.html. Retrieved 2009-06-21.
- ^ Collazo C, Chacón O, Borrás O (2006). "Programmed cell death in plants resembles apoptosis of animals". Biotecnología Aplicada 23: 1–10. http://elfosscientiae.cigb.edu.cu/PDFs/BA/2006/23/1/BA002301RV001-010.pdf.
- ^ Bonke M, Thitamadee S, Mähönen AP, Hauser MT, Helariutta Y (2003). "APL regulates vascular tissue identity in Arabidopsis". Nature 426 (6963): 181–6. doi:10.1038/nature02100. PMID 14614507.
- ^ Solomon M, Belenghi B, Delledonne M, Menachem E, Levine A (1999). "The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants". The Plant Cell 11 (3): 431–44. doi:10.2307/3870871. JSTOR 3870871. PMC 144188. PMID 10072402. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=144188. See also related articles in The Plant Cell Online
- ^ Ito J, Fukuda H (2002). "ZEN1 Is a Key Enzyme in the Degradation of Nuclear DNA during Programmed Cell Death of Tracheary Elements". The Plant Cell 14 (12): 3201–11. doi:10.1105/tpc.006411. PMC 151212. PMID 12468737. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=151212.
- ^ Balk J, Leaver CJ (2001). "The PET1-CMS Mitochondrial Mutation in Sunflower Is Associated with Premature Programmed Cell Death and Cytochrome c Release". The Plant Cell 13 (8): 1803–18. doi:10.1105/tpc.13.8.1803. PMC 139137. PMID 11487694. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=139137.
- ^ Thomas SG, Franklin-Tong VE (2004). "Self-incompatibility triggers programmed cell death in Papaver pollen". Nature 429 (6989): 305–9. doi:10.1038/nature02540. PMID 15152254.
- ^ Crespi B, Springer S (2003). "Ecology. Social slime molds meet their match". Science 299 (5603): 56–7. doi:10.1126/science.1080776. PMID 12511635.
- ^ Levraud JP, Adam M, Luciani MF, de Chastellier C, Blanton RL, Golstein P (2003). "Dictyostelium cell death: early emergence and demise of highly polarized paddle cells". Journal of Cell Biology 160 (7): 1105–14. doi:10.1083/jcb.200212104. PMC 2172757. PMID 12654899. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2172757.
- ^ Roisin-Bouffay C, Luciani MF, Klein G, Levraud JP, Adam M, Golstein P (2004). "Developmental cell death in dictyostelium does not require paracaspase". Journal of Biological Chemistry 279 (12): 11489–94. doi:10.1074/jbc.M312741200. PMID 14681218.
- ^ de Duve C (1996). "The birth of complex cells". Scientific American 274 (4): 50–7. doi:10.1038/scientificamerican0496-50. PMID 8907651.
- ^ Dyall SD, Brown MT, Johnson PJ (2004). "Ancient invasions: from endosymbionts to organelles". Science 304 (5668): 253–7. doi:10.1126/science.1094884. PMID 15073369.
- ^ Chiarugi A, Moskowitz MA (2002). "Cell biology. PARP-1--a perpetrator of apoptotic cell death?". Science 297 (5579): 200–1. doi:10.1126/science.1074592. PMID 12114611.
See also
Developmental biology > Human embryogenesis (development of embryo) and development of fetus (TE E2.0) First three
weeksWeek 1Fertilization · Oocyte activation · Zygote · Cleavage · Morula · Blastula (Blastomere) · Blastocyst · Inner cell massWeek 2
(Bilaminar)Week 3
(Trilaminar)Archenteron/Primitive streak (Primitive pit, Primitive knot/Blastopore, Primitive groove) · Gastrula/Gastrulation · Regional specification · Embryonic discSplanchnopleuric mesenchymeChorda- · Paraxial (Somite/Somitomere) · Intermediate · Lateral plate (Intraembryonic coelom, Splanchnopleuric mesenchyme/Somatopleuric mesenchyme)External links
Categories:- Programmed cell death
- Mitochondria
Wikimedia Foundation. 2010.