- Albertosaurus
-
Albertosaurus sarcophagus
Temporal range: Late Cretaceous, 73–70 Ma
Mounted skeleton, Royal Tyrrell Museum Scientific classification 
Kingdom: Animalia Phylum: Chordata Class: Reptilia Superorder: Dinosauria Order: Saurischia Suborder: Theropoda Family: †Tyrannosauridae Subfamily: †Albertosaurinae Genus: †Albertosaurus
Osborn, 1905Species: †A. sarcophagus Binomial name Albertosaurus sarcophagus
Osborn, 1905Synonyms Deinodon sarcophagus (Osborn, 1905)
Albertosaurus arctunguis Parks, 1928
Deinodon arctunguis (Parks, 1928)Albertosaurus (
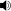 /ælˌbɜrtɵˈsɔrəs/; meaning "Alberta lizard") is a genus of tyrannosaurid theropod dinosaur that lived in western North America during the Late Cretaceous Period, more than 70 million years ago. The type species, A. sarcophagus, was apparently restricted in range to the modern-day Canadian province of Alberta, after which the genus is named. Scientists disagree on the content of the genus, with some recognizing Gorgosaurus libratus as a second species.
/ælˌbɜrtɵˈsɔrəs/; meaning "Alberta lizard") is a genus of tyrannosaurid theropod dinosaur that lived in western North America during the Late Cretaceous Period, more than 70 million years ago. The type species, A. sarcophagus, was apparently restricted in range to the modern-day Canadian province of Alberta, after which the genus is named. Scientists disagree on the content of the genus, with some recognizing Gorgosaurus libratus as a second species.As a tyrannosaurid, Albertosaurus was a bipedal predator with tiny, two-fingered hands and a massive head with dozens of large, sharp teeth. It may have been at the top of the food chain in its local ecosystem. Although relatively large for a theropod, Albertosaurus was much smaller than its more famous relative Tyrannosaurus, probably weighing less than 2 metric tons.
Since the first discovery in 1884, fossils of more than thirty individuals have been recovered, providing scientists with a more detailed knowledge of Albertosaurus anatomy than is available for most other tyrannosaurids. The discovery of 22 individuals at one site provides evidence of pack behaviour and allows studies of ontogeny and population biology which are impossible with lesser-known dinosaurs.
Contents
Description
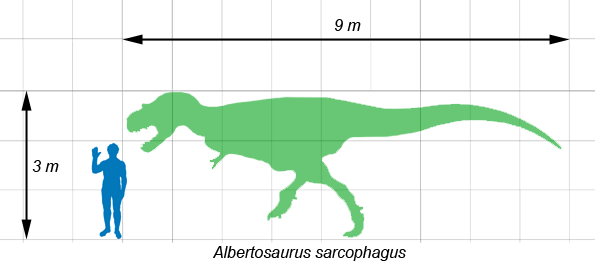
 Albertosaurus with a human for scale
Albertosaurus with a human for scale
Albertosaurus was smaller than some other tyrannosaurids, such as Tarbosaurus and Tyrannosaurus. Typical adults of Albertosaurus measured up to 9 metres (30 ft) long,[1][2] while rare individuals of great age could grow to over 10 metres (33 ft) in length.[3] Several independent mass estimates, obtained by different methods, suggest that an adult Albertosaurus weighed between 1.3 tonnes (1.4 short tons)[4] and 1.7 tonnes (1.9 tons).[5]
All tyrannosaurids, including Albertosaurus, shared a similar body appearance. Typically for a theropod, Albertosaurus was bipedal and balanced the heavy head and torso with a long tail. However, tyrannosaurid forelimbs were extremely small for their body size and retained only two digits. The hind limbs were long and ended in a four-toed foot. The first digit, called the hallux, was short and only the other three contacted the ground, with the third (middle) digit longer than the rest.[2] Albertosaurus may have been able to reach speeds of 14−21 kilometres per hour (8−13 miles per hour).[6]
Skull and teeth
 Reconstruction
ReconstructionThe massive skull of Albertosaurus, perched on a short, S-shaped neck, was approximately 1 metre (3.3 ft) long in the largest adults.[7] Wide openings in the skull (fenestrae) reduced the weight of the head while also providing space for muscle attachment and sensory organs. Its long jaws contained more than 60 banana-shaped teeth; larger tyrannosaurids possessed fewer teeth. Unlike most theropods, Albertosaurus and other tyrannosaurids were heterodont, with teeth of different forms depending on their position in the mouth. The premaxillary teeth at the tip of the upper jaw were much smaller than the rest, more closely packed, and D-shaped in cross section.[2] Above the eyes were short bony crests that may have been brightly coloured in life and used in courtship to attract a mate.[8]
 Skull cast
Skull castWilliam L. Abler observed in 2001 that Albertosaurus tooth serrations are so thin as to functionally be a crack in the tooth.[9] However, at the base of this crack is a round void called an ampulla which would have functioned to distribute force over a larger surface area, hindering the ability of the "crack" formed by the serration to propagate through the tooth.[9] An examination of other ancient predators, a phytosaur and Dimetrodon found similarly crack-like serrations, but no adaptations for preventing crack propagation.[9] Tyrannosaurid teeth were used as holdfasts for pulling meat off a body, rather than knife-like cutting functions.[9] Tooth wear patterns hint that complex head shaking behaviours may have been involved in tyrannosaur feeding.[9] When a tyrannosaur would have pulled back on a piece of meat, the force would tend to push the tip of tooth toward the front of the mouth and the anchored root would experience tension on the posterior side and compression from the front.[9] This would typically incline the tooth to crack formation on the posterior side of the tooth, but the ampullae at the base of the already crack-like serrations would tend to diffuse potential crack-forming forces.[9] This form resembles techniques used by guitar makers to "impart alternating regions of flexibility and rigidity to a stick of wood."[9] The use of a drill to create an "ampulla" of sorts and prevent the propagation of cracks through an important material is also used to protect airplane surfaces.[9] Abler demonstrated that a plexiglass bar with kerfs and drilled holes was more than 25% stronger than one with only regularly placed incisions.[9]
Classification and systematics
Cladogram of Tyrannosauridae highlighting the position of AlbertosaurusTyrannosauridae Tyrannosaurinae
Albertosaurinae Albertosaurus
Albertosaurus is a member of the theropod family Tyrannosauridae, in the subfamily Albertosaurinae. Its closest relative is the slightly older Gorgosaurus libratus (sometimes called Albertosaurus libratus; see below).[10] These two species are the only described albertosaurines; other undescribed species may exist.[11] Thomas Holtz found Appalachiosaurus to be an albertosaurine in 2004,[2] but his more recent unpublished work locates it just outside Tyrannosauridae,[12] in agreement with other authors.[13]
The other major subfamily of tyrannosaurids is the Tyrannosaurinae, including Daspletosaurus, Tarbosaurus and Tyrannosaurus. Compared with these robust tyrannosaurines, albertosaurines had slender builds, with proportionately smaller skulls and longer bones of the lower leg (tibia) and feet (metatarsals and phalanges).[7][10]
Discovery and naming
Naming
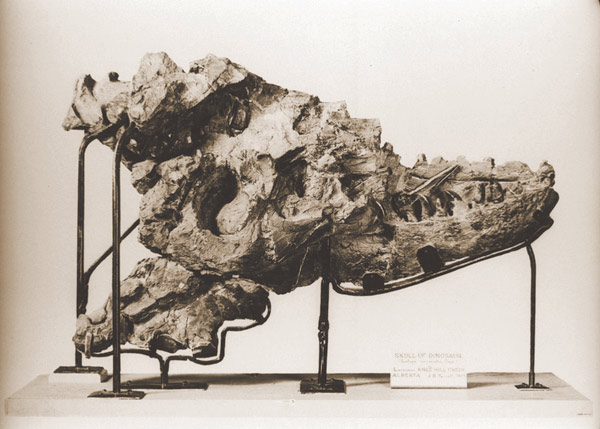
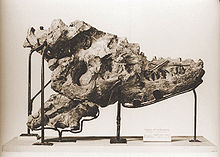 Type specimen
Type specimenAlbertosaurus was named by Henry Fairfield Osborn in a one-page note at the end of his 1905 description of Tyrannosaurus rex.[14]:265 The name honours Alberta, the present-day Canadian province in which the first remains were found. The generic name also incorporates the Greek term σαυρος/sauros ("lizard"), the most common suffix in dinosaur names. The type species is A. sarcophagus, which means "flesh-eater" and has the same etymology as the funeral container with which it shares its name: a combination of the Ancient Greek words σαρξ/sarx ("flesh") and Φαγειν/phagein ("to eat").[14] More than thirty specimens of all ages are known to science.[3][11]
Early discoveries
 The Red Deer River near Drumheller, Alberta. Almost three-quarters of all Albertosaurus remains have been discovered alongside the river, in outcrops like the ones on either side of this picture.
The Red Deer River near Drumheller, Alberta. Almost three-quarters of all Albertosaurus remains have been discovered alongside the river, in outcrops like the ones on either side of this picture.The type specimen is a partial skull, collected in 1884 from an outcrop of the Horseshoe Canyon Formation alongside the Red Deer River, in present-day Alberta. This specimen and a smaller skull associated with some skeletal material were recovered by expeditions of the Geological Survey of Canada, led by the famous geologist Joseph B. Tyrrell. The two skulls were assigned to the preexisting species Laelaps incrassatus by Edward Drinker Cope in 1892,[15] although the name Laelaps was preoccupied by a genus of mite and had been changed to Dryptosaurus in 1877 by Othniel Charles Marsh. Cope refused to recognize the new name created by his archrival Marsh, so it fell to Lawrence Lambe to change Laelaps incrassatus to Dryptosaurus incrassatus when he described the remains in detail in 1904.[16] Shortly later, Osborn pointed out that D. incrassatus was based on generic tyrannosaurid teeth, so the two Horseshoe Canyon skulls could not be confidently referred to that species. The Horseshoe Canyon skulls also differed markedly from the remains of D. aquilunguis, type species of Dryptosaurus, so Osborn created the new name Albertosaurus sarcophagus for them in 1905. He did not describe the remains in any great detail, citing Lambe's complete description the year before.[14] Both specimens (CMN 5600 and 5601) are stored in the Canadian Museum of Nature in Ottawa.
Dry Island bonebed
In 1910, American paleontologist Barnum Brown uncovered the remains of a large group of Albertosaurus at another quarry alongside the Red Deer River. Because of the large number of bones and the limited time available, Brown's party did not collect every specimen, but made sure to collect remains from all of the individuals they could identify in the bonebed. Among the bones deposited in the American Museum of Natural History collections in New York City are seven sets of right metatarsals, along with two isolated toe bones that did not match any of the metatarsals in size. This indicated the presence of at least nine individuals in the quarry. The Royal Tyrrell Museum of Palaeontology rediscovered the bonebed in 1997 and resumed fieldwork at the site, which is now located inside Dry Island Buffalo Jump Provincial Park.[17] Further excavation from 1997 to 2005 turned up the remains of 13 more individuals of various ages, including a diminutive two-year-old and a very old individual estimated at over 10 metres (33 ft) in length. None of these individuals are known from complete skeletons, and most are represented by remains in both museums.[3][4]
Gorgosaurus libratus
In 1913, paleontologist Charles H. Sternberg recovered another tyrannosaurid skeleton from the slightly older Dinosaur Park Formation in Alberta. Lawrence Lambe named this dinosaur Gorgosaurus libratus in 1914.[18] Other specimens were later found in Alberta and the US state of Montana. Finding few differences to separate the two genera, Dale Russell declared the name Gorgosaurus a junior synonym of Albertosaurus, which had been named first, and G. libratus was renamed Albertosaurus libratus in 1970. This addition extended the temporal range of the genus Albertosaurus backwards by several million years and its geographic range southwards by hundreds of kilometres.[1]
In 2003, Philip J. Currie compared several tyrannosaurid skulls and came to the conclusion that the two species are more distinct than previously thought. The decision to use one or two genera is rather arbitrary, as the two species are sister taxa, more closely related to each other than to any other species. Recognizing this, Currie nevertheless recommended that Albertosaurus and Gorgosaurus be retained as separate genera, as they are no more similar than Daspletosaurus and Tyrannosaurus, which are almost always separated. In addition, several albertosaurine specimens have been recovered from Alaska and New Mexico, and Currie suggested that the Albertosaurus-Gorgosaurus situation may be clarified once these are described fully.[11] Most authors have followed Currie's recommendation,[2][4][19] but some have not.[13]
Other discoveries
William Parks described a new species, Albertosaurus arctunguis, based on a partial skeleton excavated near the Red Deer River in 1928,[20] but this species has been considered identical to A. sarcophagus since 1970.[1] Parks' specimen (ROM 807) is housed in the Royal Ontario Museum in Toronto. Six more skulls and skeletons have since been discovered in Alberta and are housed in various Canadian museums. Fossils have also been reported from the American states of Montana, New Mexico, and Wyoming, but these probably do not represent A. sarcophagus and may not even belong to the genus Albertosaurus.[2][11]
Albertosaurus megagracilis was based on a small tyrannosaurid skeleton from the Hell Creek Formation of Montana.[21] It was renamed Dinotyrannus in 1995,[22] but is now thought to represent a juvenile Tyrannosaurus rex.[7]
Paleobiology
Growth pattern
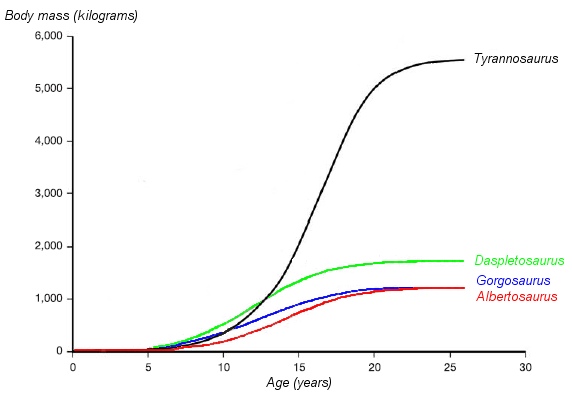
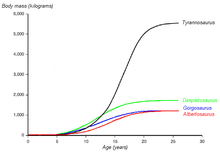 A graph showing the hypothesized growth curves (body mass versus age) of four tyrannosaurids, with Albertosaurus drawn in red[4]
A graph showing the hypothesized growth curves (body mass versus age) of four tyrannosaurids, with Albertosaurus drawn in red[4]Most age categories of Albertosaurus are represented in the fossil record. Using bone histology, the age of an individual animal at the time of death can often be determined, allowing growth rates to be estimated and compared with other species. The youngest known Albertosaurus is a two-year-old discovered in the Dry Island bonebed, which would have weighed about 50 kilograms (110 lb) and measured slightly more than 2 metres (7 ft) in length. The 10 metre (33 ft) specimen from the same quarry is the oldest and largest known, at 28 years of age. When specimens of intermediate age and size are plotted on a graph, an S-shaped growth curve results, with the most rapid growth occurring in a four-year period ending around the sixteenth year of life, a pattern also seen in other tyrannosaurids. The growth rate during this phase was 122 kilograms (268 lb) per year, based on an adult 1.3 tonnes (1.4 short tons). Other studies have suggested higher adult weights; this would affect the magnitude of the growth rate but not the overall pattern. Tyrannosaurids similar in size to Albertosaurus had similar growth rates, although the much larger Tyrannosaurus rex grew almost five times faster (601 kilograms [1325 lb] per year) at its peak.[3] The end of the rapid growth phase suggests the onset of sexual maturity in Albertosaurus, although growth continued at a slower rate throughout the animals' lives.[3][4] Sexual maturation while still actively growing appears to be a shared trait among small[23] and large[24] dinosaurs as well as in large mammals such as humans and elephants.[24] This pattern of relatively early sexual maturation differs strikingly from the pattern in birds, which delay their sexual maturity until after they have finished growing.[19][24]
Life history
Most known Albertosaurus individuals were aged 14 years or more at the time of death. Juvenile animals are rarely found as fossils for several reasons, mainly preservation bias, where the smaller bones of younger animals were less likely to be preserved by fossilization than the larger bones of adults, and collection bias, where smaller fossils are less likely to be noticed by collectors in the field.[25] Young Albertosaurus are relatively large for juvenile animals, but their remains are still rare in the fossil record compared with adults. It has been suggested that this phenomenon is a consequence of life history, rather than bias, and that fossils of juvenile Albertosaurus are rare because they simply did not die as often as adults did.[3]
A hypothesis of Albertosaurus life history postulates that hatchlings died in large numbers, but have not been preserved in the fossil record due to their small size and fragile construction. After just two years, juveniles were larger than any other predator in the region aside from adult Albertosaurus, and more fleet of foot than most of their prey animals. This resulted in a dramatic decrease in their mortality rate and a corresponding rarity of fossil remains. Mortality rates doubled at age twelve, perhaps the result of the physiological demands of the rapid growth phase, and then doubled again with the onset of sexual maturity between the ages of fourteen and sixteen. This elevated mortality rate continued throughout adulthood, perhaps due to high physiological demands, stress and injuries received during intraspecific competition for mates and resources, and eventually, the ever-increasing effects of senescence. The higher mortality rate in adults may explain their more common preservation. Very large animals were rare because few individuals survived long enough to attain such sizes. High infant mortality rates, followed by reduced mortality among juveniles and a sudden increase in mortality after sexual maturity, with very few animals reaching maximum size, is a pattern observed in many modern large mammals, including elephants, African buffalo, and rhinoceros. The same pattern is also seen in other tyrannosaurids. The comparison with modern animals and other tyrannosaurids lends support to this life history hypothesis, but bias in the fossil record may still play a large role, especially since more than two-thirds of all Albertosaurus specimens are known from one locality.[3][19]
In 2009, researchers hypothesized that smooth-edged holes found in the fossil jaws of tyrannosaurid dinosaurs such as Albertosaurus were caused by a parasite similar to Trichomonas gallinae which infects birds.[26] They suggested that tyrannosaurids transmitted the infection by biting each other, and that the infection impaired their ability to eat food.[26]
Pack behaviour
The Dry Island bonebed discovered by Barnum Brown and his crew contains the remains of 22 Albertosaurus, the most individuals found in one locality of any Cretaceous theropod, and the second-most of any large theropod dinosaur behind the Allosaurus assemblage at the Cleveland Lloyd Dinosaur Quarry in Utah. The group seems to be composed of one very old adult; eight adults between 17 and 23 years old; seven sub-adults undergoing their rapid growth phases at between 12 and 16 years old; and six juveniles between the ages of 2 and 11 years, who had not yet reached the growth phase.[3]
 Albertosaurus models, Royal Tyrell Museum.
Albertosaurus models, Royal Tyrell Museum.The near-absence of herbivore remains and the similar state of preservation common to the many individuals at the Albertosaurus bonebed quarry led Currie to conclude that the locality was not a predator trap like the La Brea Tar Pits in California, and that all of the preserved animals died at the same time. Currie claims this as evidence of pack behaviour.[17] Other scientists are skeptical, observing that the animals may have been driven together by drought, flood or for other reasons.[3][25][27]
There is plentiful evidence for gregarious behaviour among herbivorous dinosaurs, including ceratopsians and hadrosaurs.[28] However, only rarely are so many dinosaurian predators found at the same site. Small theropods like Deinonychus,[29] Coelophysis and Megapnosaurus (Syntarsus) rhodesiensis[30] have been found in aggregations, as have larger predators like Allosaurus and Mapusaurus.[31] There is some evidence of gregarious behaviour in other tyrannosaurids as well. Fragmentary remains of smaller individuals were found alongside "Sue," the Tyrannosaurus mounted in the Field Museum of Natural History in Chicago, and a bonebed in the Two Medicine Formation of Montana contains at least three specimens of Daspletosaurus, preserved alongside several hadrosaurs.[32] These findings may corroborate the evidence for social behaviour in Albertosaurus, although some or all of the above localities may represent temporary or unnatural aggregations.[17] Others have speculated that instead of social groups, at least some of these finds represent Komodo dragon-like mobbing of carcasses, where aggressive competition leads to some of the predators being killed and cannibalized.[25]
Currie also offers speculation on the pack-hunting habits of Albertosaurus. The leg proportions of the smaller individuals were comparable to those of ornithomimids, which were probably among the fastest dinosaurs. Younger Albertosaurus were probably equally fleet-footed, or at least faster than their prey. Currie hypothesized that the younger members of the pack may have been responsible for driving their prey towards the adults, who were larger and more powerful, but also slower.[17] Juveniles may also have had different lifestyles than adults, filling predator niches between the enormous adults and the smaller contemporaneous theropods, the largest of which were two orders of magnitude smaller than adult Albertosaurus in mass.[2] A similar situation is observed in modern Komodo dragons, with hatchlings beginning life as small insectivores before growing to become the dominant predators on their islands.[33] However, as the preservation of behaviour in the fossil record is exceedingly rare, these ideas cannot readily be tested.
Paleopathology
In 2001, Bruce Rothschild and others published a study examining evidence for stress fractures and tendon avulsions in theropod dinosaurs and the implications for their behavior. They found that only one of the 319 Albertosaurus foot bones checked for stress fractures actually had them and none of the four hand bones did. The scientists found that stress fractures were "significantly" less common in Albertosaurus than in the carnosaur Allosaurus.[34] ROM 807, the holotype of A. arctunguis (now referred to A. sarcophagus), had a 2.5 by 3.5 cm deep hole in the iliac blade, although the describer of the species did not recognize this as pathological. The specimen also contains some exostosis on the fourth left metatarsal. Two of the five Albertosaurus sarcophagus specimens with humeri in 1970 were reported by Dale Russel as having pathological damage to them.[35]
Paleoecology

All identifiable fossils of Albertosaurus sarcophagus are known from the Horseshoe Canyon Formation in Alberta. This geologic formation dates to the early Maastrichtian stage of the Late Cretaceous Period, 73 to 70 Ma (million years ago). Immediately below this formation is the Bearpaw Shale, a marine formation representing a section of the Western Interior Seaway. The seaway was receding as the climate cooled and sea levels subsided towards the end of the Cretaceous, exposing land that had previously been underwater. It was not a smooth process, however, and the seaway would periodically rise to cover parts of the region throughout Horseshoe Canyon times before finally receding altogether in the years after. Due to the changing sea levels, many different environments are represented in the Horseshoe Canyon Formation, including offshore and near-shore marine habitats and coastal habitats like lagoons, estuaries and tidal flats. Numerous coal seams represent ancient peat swamps. Like most of the other vertebrate fossils from the formation, Albertosaurus remains are found in deposits laid down in the deltas and floodplains of large rivers during the later half of Horseshoe Canyon times.[36]
The fauna of the Horseshoe Canyon Formation is well-known, as vertebrate fossils, including those of dinosaurs, are quite common. Sharks, rays, sturgeons, bowfins, gars and the gar-like Aspidorhynchus made up the fish fauna. Mammals included multituberculates and the marsupial Didelphodon. The saltwater plesiosaur Leurospondylus has been found in marine sediments in the Horseshoe Canyon, while freshwater environments were populated by turtles, Champsosaurus, and crocodilians like Leidyosuchus and Stangerochampsa. Dinosaurs dominate the fauna, especially hadrosaurs, which make up half of all dinosaurs known, including the genera Edmontosaurus, Saurolophus and Hypacrosaurus. Ceratopsians and ornithomimids were also very common, together making up another third of the known fauna. Along with much rarer ankylosaurians and pachycephalosaurs, all of these animals would have been prey for a diverse array of carnivorous theropods, including troodontids, dromaeosaurids, and caenagnathids. Adult Albertosaurus were the apex predators in this environment, with intermediate niches possibly filled by juvenile albertosaurs.[36]
References
- ^ a b c Russell, Dale A. (1970). "Tyrannosaurs from the Late Cretaceous of western Canada". National Museum of Natural Sciences Publications in Paleontology 1: 1–34.
- ^ a b c d e f g Holtz, Thomas R. (2004). "Tyrannosauroidea". In Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 111–136. ISBN 0-520-24209-2.
- ^ a b c d e f g h i Erickson, Gregory M.; Currie, Philip. J.; Inouye, Brian D.; & Wynn, Alice A. (2006). "Tyrannosaur life tables: an example of nonavian dinosaur population biology". Science 313 (5784): 213–217. doi:10.1126/science.1125721. PMID 16840697. http://www.nceas.ucsb.edu/jai/podcast_publications/inouye_science_dinosaur.pdf. Retrieved 2010-08-29.
- ^ a b c d e Erickson, Gregory M., GM; Makovicky, Peter J.; Currie, Philip J.; Norell, Mark A.; Yerby, Scott A.; & Brochu, Christopher A. (2004). "Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs". Nature 430 (7001): 772–775. doi:10.1038/nature02699. PMID 15306807. http://www.biology.ualberta.ca/courses.hp/biol506.hp/pdfs/Erickson'04_Nature.pdf. Retrieved 2010-08-29.
- ^ Christiansen, Per; & Fariña, Richard A. (2004). "Mass prediction in theropod dinosaurs". Historical Biology 16 (2-4): 85–92. doi:10.1080/08912960412331284313.
- ^ Thulborn, Richard A. (1982). "Speeds and gaits of dinosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology 38 (3-4): 227−256. doi:10.1016/0031-0182(82)90005-0.
- ^ a b c Currie, Philip J. (2003). "Allometric growth in tyrannosaurids (Dinosauria: Theropoda) from the Upper Cretaceous of North America and Asia". Canadian Journal of Earth Sciences 40 (4): 651–665. doi:10.1139/e02-083. http://www.ingentaconnect.com/content/nrc/cjes/2003/00000040/00000004/art00014.
- ^ "Albertosaurus." In: Dodson, Peter; Britt, Brooks; Carpenter, Kenneth; Forster, Catherine A.; Gillette, David D.; Norell, Mark A.; Olshevsky, George; Parrish, J. Michael; & Weishampel, David B. The Age of Dinosaurs. Lincolnwood, IL: Publications International, Ltd., 1993. pp. 106-107. ISBN 0-7853-0443-6.
- ^ a b c d e f g h i j Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84-89. In: Mesozioc Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- ^ a b Currie, Philip J.; Hurum, Jørn H; & Sabath, Karol (2003). "Skull structure and evolution in tyrannosaurid phylogeny" (PDF). Acta Palaeontologica Polonica 48 (2): 227–234. http://app.pan.pl/archive/published/app48/app48-227.pdf. Retrieved 2009-05-03.
- ^ a b c d Currie, Philip J. (2003). "Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta" (PDF). Acta Palaeontologica Polonica 48 (2): 191–226. http://app.pan.pl/archive/published/app48/app48-191.pdf.
- ^ Holtz, Thomas R. (2005-09-20). "RE: Burpee Conference (LONG)". http://dml.cmnh.org/2005Sep/msg00345.html. Retrieved 2007-06-18.
- ^ a b Carr, Thomas D.; Williamson, Thomas E.; & Schwimmer, David R. (2005). "A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama". Journal of Vertebrate Paleontology 25 (1): 119–143. doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2.
- ^ a b c Osborn, Henry F. (1905). "Tyrannosaurus and other Cretaceous carnivorous dinosaurs". Bulletin of the American Museum of Natural History 21: 259–265. doi:10.1111/j.1468-5965.2007.00735_17.x. hdl:2246/1464.
- ^ Cope, Edward D. (1892). "On the skull of the dinosaurian Laelaps incrassatus Cope". American Philosophical Society, Proceedings 30: 240–245.
- ^ Lambe, Lawrence M. (1904). "On Dryptosaurus incrassatus (Cope) from the Edmonton Series of the Northwest Territory". Contributions to Canadian Palaeontology 3: 1–27. http://www.archive.org/details/LambeL.M.1904.OnDryptosaurusIncrassatuscopeFromTheEdmonton. Retrieved 2010-08-29.
- ^ a b c d Currie, Philip J. (1998). "Possible evidence of gregarious behavior in tyrannosaurids" (PDF). Gaia 15: 271–277. http://www.mnhn.ul.pt/geologia/gaia/21.pdf. Retrieved 2009-05-03. (not printed until 2000)
- ^ Lambe, Lawrence M. (1914). "On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon". Ottawa Naturalist 28: 13–20.
- ^ a b c Ricklefs, Robert E. (2007). "Tyrannosaur ageing". Biology Letters 3 (2): 214–217. doi:10.1098/rsbl.2006.0597. PMC 2375931. PMID 17284406. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2375931.
- ^ Parks, William A. (1928). "Albertosaurus arctunguis, a new species of therapodous dinosaur from the Edmonton Formation of Alberta". University of Toronto Studies, Geological Series 25: 1–42.
- ^ Paul, Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. ISBN 978-0-671-61946-6.
- ^ Olshevsky, George. (1995). "(The origin and evolution of the tyrannosaurids.)" (in Japanese). Kyoryugaku Saizensen (Dino Frontline) 9: 92–119.
- ^ Erickson, Gregory M.; Curry Rogers, Kristi; Varricchio, David J.; Norell, Mark.; & Xu, Xing (2007). "Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition". Biology Letters 3 (5): 558–561. doi:10.1098/rsbl.2007.0254. PMC 2396186. PMID 17638674. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2396186.
- ^ a b c Lee, Andrew H.; Werning, Sarah (2008). "Sexual maturity in growing dinosaurs does not fit reptilian growth models". PNAS 105 (2): 582–587. doi:10.1073/pnas.0708903105. PMC 2206579. PMID 18195356. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2206579.
- ^ a b c Roach, Brian T.; & Brinkman, Daniel T. (2007). "A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs". Bulletin of the Peabody Museum of Natural History 48 (1): 103–138. doi:10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ^ a b Wolff, Ewan D. S.; Salisbury, Steven W.; Horner, John R.; & Varricchio, David J. (2009). Hansen, Dennis Marinus. ed. "Common avian infection plagued the tyrant dinosaurs". PLoS ONE 4 (9): e7288. doi:10.1371/journal.pone.0007288. PMC 2748709. PMID 19789646. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2748709.
- ^ Eberth, David A.; & McCrea, Richard T. (2001). "Were large theropods gregarious?". Journal of Vertebrate Paleontology 21 (Supplement to 3 - Abstracts of Papers, 61st Annual Meeting of the Society of Vertebrate Paleontology): 46A.(published abstract only)
- ^ Horner, John R. (1997). "Behavior". In Currie, Philip J.; & Padian, Kevin (eds.). Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 45–50. ISBN 0-12-226810-5.
- ^ Maxwell, W. Desmond; & Ostrom, John H. (1995). "Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations". Journal of Vertebrate Paleontology 15 (4): 707–712. doi:10.1080/02724634.1995.10011256.
- ^ Raath, Michael A. (1990). "Morphological variation in small theropods and its meaning in systematics: evidence from Syntarsus rhodesiensis". In Carpenter, Kenneth & Currie, Philip J.. Dinosaur Systematics: Approaches and Perspectives. Cambridge: Cambridge University Press. pp. 91–105. ISBN 0-521-43810-1.
- ^ Coria, Rodolfo A.; & Currie, Philip J. (2006). "A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina" (PDF). Geodiversitas 28 (1): 71–118. http://www.mnhn.fr/museum/front/medias/publication/7653_g06n1a4.pdf. Retrieved 2009-05-03.
- ^ Currie, Philip J.; Trexler, David; Koppelhus, Eva B.; Wicks, Kelly; & Murphy, Nate. (2005). "An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA)". In Carpenter, Kenneth (ed.).. The Carnivorous Dinosaurs. Bloomington: Indiana University Press. pp. 313–324. ISBN 978-0-253-34539-4.
- ^ Auffenberg, Walter (2000). The Behavioral Ecology of the Komodo Monitor. Gainesville: University Press of Florida. ISBN 978-0-8130-0621-5.
- ^ Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331-336.
- ^ Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.
- ^ a b Eberth, David A. (1997). "Edmonton Group". In Currie, Philip J. & Padian, Kevin (eds.). Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 199–204. ISBN 0-12-226810-5.
External links
- American Museum of Natural History, Division of Paleontology. Search results for "Albertosaurus" in the Collections Database. Sample entries:
- Bureau of Land Management, New Mexico State Office. Operation Dino-Lift. Summarizes the excavation and transport by helicopter of a possible Albertosaurus skeleton; however, per Carr & Williamson (2010), specimen may later have been assigned to Bistahieversor.
- Dykens, Margaret; & Gillette, Lynett. Fossil Mysteries Field Guide: Albertosaurus. San Diego Natural History Museum.
- Geological Survey of Canada. Past lives: Chronicles of Canadian Paleontology. Tyrrell and Albertosaurus. About the discovery of the holotype skull of A. sarcophagus. 2008-02-07.
- Hanson, Michael. A. sarcophagus. (Skeletal drawing from The Grave Yard: A Skeletal Reconstructions Database, 2007.)
- Mortimer, Mickey. Albertosaurus Osborn, 1905. (List of specimens from The Theropod Database.)
Categories:- Cretaceous dinosaurs
- Dinosaurs of North America
- Tyrannosaurs






Wikimedia Foundation. 2010.