- DNA glycosylase
-
DNA glycosylases are a family of enzymes involved in base excision repair, classified under EC number EC 3.2.2. Base excision repair is the mechanism by which damaged bases in DNA are removed and replaced. DNA glycosylases catalyze the first step of this process. They remove the damaged nitrogenous base while leaving the sugar-phosphate backbone intact, creating an apurinic/apyrimidinic site, commonly referred to as an AP site. This is accomplished by flipping the damaged base out of the double helix followed by cleavage of the N-glycosidic bond. Glycosylases were first discovered in bacteria, and have since been found in all kingdoms of life. In addition to their role in base excision repair DNA glycosylase enzymes have been implicated in the repression of gene silencing in A. thaliana, N. tabacum and other plants by active demethylation. 5-methylcytosine residues are excised and replaced with unmethylated cytosines allowing acces to the chromatin structure of the enzymes and proteins necessary for trancription and subsequent translation[1][2].
Contents
Monofunctional vs. bifunctional glycosylases
There are two main classes of glycosylases: monofunctional and bifunctional. Monofunctional glycosylases have only glycosylase activity, whereas bifunctional glycosylases also possess AP lyase activity that permits them to cut the phosphodiester bond of DNA, creating a single-strand break without the need for an AP endonuclease. β-Elimination of an AP site by a glycosylase-lyase yields a 3' α,β-unsaturated aldehyde adjacent to a 5' phosphate, which differs from the AP endonuclease cleavage product.[3] Some glycosylase-lyases can further perform δ-elimination, which converts the 3' aldehyde to a 3' phosphate.
Biochemical mechanism
The first crystal structure of a DNA glycosylase was obtained for E. coli Nth.[4] This structure revealed that the enzyme flips the damaged base out of the double helix into an active site pocket in order to excise it. Other glycosylases have since been found to follow the same general paradigm, including human UNG pictured below. To cleave the N-glycosidic bond, monofunctional glycosylases use an activated water molecule to attack carbon 1 of the substrate. Bifunctional glycosylases, instead, use an amine residue as a nucleophile to attack the same carbon, going through a Schiff base intermediate.
Types of glycosylases
Crystal structures of many glycosylases have been solved. Based on structural similarity, glycosylases are grouped into four superfamilies. The UDG and AAG families contain small, compact glycosylases, whereas the MutM/Fpg and HhH-GPD families comprise larger enzymes with multiple domains.[3]
A wide variety of glycosylases have evolved to recognize different damaged bases. The table below summarizes the properties of known glycosylases in commonly studied model organisms.
Glycosylases in bacteria, yeast and humans[5][6] E. coli Yeast (S. cerevisiae) Human Type Substrates AlkA Mag1 MPG monofunctional 3-meA, hypoxanthine UDG Ung1 UNG monofunctional uracil Fpg Ogg1 hOGG1 bifunctional 8-oxoG, FapyG Nth Ntg1 hNTH1 bifunctional Tg, hoU, hoC, urea, FapyG Ntg2 Nei Not present hNEIL1 bifunctional Tg, hoU, hoC, urea, FapyG, FapyA hNEIL2 AP site, hoU hNEIL3 unknown MutY Not present hMYH monofunctional A:8-oxoG Not present Not present hSMUG1 monofunctional U, hoU, hmU, fU Not present Not present TDG monofunctional T:G mispair Not present Not present MBD4 monofunctional T:G mispair DNA glycosylases can be grouped into the following categories based on their substrate(s):
Uracil DNA glycosylases
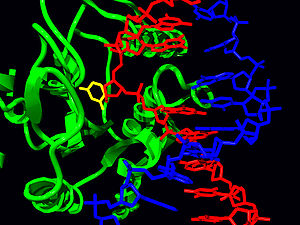 Structure of the base-excision repair enzyme uracil-DNA glycosylase. The uracil residue is shown in yellow.
Structure of the base-excision repair enzyme uracil-DNA glycosylase. The uracil residue is shown in yellow.
Uracil DNA glycosylases remove uracil from DNA, which can arise either by spontaneous deamination of cytosine or by the misincorporation of dU opposite dA during DNA replication. The prototypical member of this family is E. coli UDG, which was among the first glycosylases discovered. Four different uracil-DNA glycosylase activities have been identified in mammalian cells, including UNG, SMUG1, TDG, and MBD4. They vary in substrate specificity and subcellular localization. SMUG1 prefers single-stranded DNA as substrate, but also removes U from double-stranded DNA. In addition to unmodified uracil, SMUG1 can excise 5-hydroxyuracil, 5-hydroxymethyluracil and 5-formyluracil bearing an oxidized group at ring C5.[7] TDG and MBD4 are strictly specific for double-stranded DNA. TDG can remove thymine glycol when present opposite guanine, as well as derivatives of U with modifications at carbon 5. Current evidence suggests that, in human cells, TDG and SMUG1 are the major enzymes responsible for the repair of the U:G mispairs caused by spontaneous cytosine deamination, whereas uracil arising in DNA through dU misincorporation is mainly dealt with by UNG. MBD4 is thought to correct T:G mismatches that arise from deamination of 5-methylcytosine to thymine in CpG sites.[8] MBD4 mutant mice develop normally and do not show increased cancer susceptibility or reduced survival. But they acquire more C T mutations at CpG sequences in epithelial cells of the small intestine.[9]
The structure of human UNG in complex with DNA revealed that, like other glycosylases, it flips the target nucleotide out of the double helix and into the active site pocket.[10] UDG undergoes a conformational change from an ‘‘open’’ unbound state to a ‘‘closed’’ DNA-bound state.[11]
Glycosylases of oxidized bases
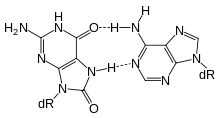 8-oxoG (syn) in a Hoogsteen base pair with dA (anti)
8-oxoG (syn) in a Hoogsteen base pair with dA (anti)A variety of glycosylases have evolved to recognize oxidized bases, which are commonly formed by reactive oxygen species generated during cellular metabolism. The most abundant lesions formed at guanine residues are 2,6-diamino-4-hydroxy-5-formamidopyrimidine (FapyG) and 8-oxoguanine. Due to mispairing with adenine during replication, 8-oxoG is highly mutagenic, resulting in G to T transversions. Repair of this lesion is initiated by the bifunctional DNA glycosylase OGG1, which recognizes 8-oxoG paired with C. hOGG1 is a bifunctional glycosylase that belongs to the helix-hairpin-helix (HhH) family. MYH recognizes adenine mispaired with 8-oxoG but excises the A, leaving the 8-oxoG intact. OGG1 knockout mice do not show an increased tumor incidence, but accumulate 8-oxoG in the liver as they age.[12] A similar phenotype is observed with the inactivation of MYH, but simultaneous inactivation of both MYH and OGG1 causes 8-oxoG accumulation in multiple tissues including lung and small intestine.[13] In humans, mutations in MYH are associated with increased risk of developing colon polyps and colon cancer. In addition to OGG1 and MYH, human cells contain three additional DNA glycosylases, NEIL1, NEIL2, and NEIL3. These are homologous to bacterial Nei, and their presence likely explains the mild phenotypes of the OGG1 and MYH knockout mice.
Glycosylases of alkylated bases
This group includes E. coli AlkA and related proteins in higher eukaryotes. These glycosylases are monofunctional and recognize methylated bases, such as 3-methyladenine.
References
- ^ Aguis, F.; Kapoor, A. and Zhu. J-K. (2006). "Role of the Arabidopsis DNA glycosylase/lyase ROS1 in active DNA demethylation". Proc. Natl. ACad. Sci. 103 (31): 11796–11801.
- ^ Choi, C-S.; Sano, H. (2007). "Identification of tobacco genes encoding proteins possessing removal activity of 5-methylcytosines from intact tobacco DNA". Plant Biotechnology 24: 339–344.
- ^ a b Fromme JC, Banerjee A, Verdine GL (February 2004). "DNA glycosylase recognition and catalysis". Curr. Opin. Struct. Biol. 14 (1): 43–9. doi:10.1016/j.sbi.2004.01.003. PMID 15102448.
- ^ Kuo CF, McRee DE, Fisher CL, O'Handley SF, Cunningham RP, Tainer JA (October 1992). "Atomic structure of the DNA repair [4Fe-4S] enzyme endonuclease III". Science 258 (5081): 434–40. doi:10.1126/science.1411536. PMID 1411536.
- ^ Ide H, Kotera M (April 2004). "Human DNA glycosylases involved in the repair of oxidatively damaged DNA". Biol. Pharm. Bull. 27 (4): 480–5. doi:10.1248/bpb.27.480. PMID 15056851.
- ^ Alseth I, Osman F, Korvald H, et al. (2005). "Biochemical characterization and DNA repair pathway interactions of Mag1-mediated base excision repair in Schizosaccharomyces pombe". Nucleic Acids Res. 33 (3): 1123–31. doi:10.1093/nar/gki259. PMC 549418. PMID 15722486. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=549418.
- ^ Matsubara M, Tanaka T, Terato H, Ohmae E, Izumi S, Katayanagi K, Ide H. (2004). Mutational analysis of the damage-recognition and catalytic mechanism of human SMUG1 DNA glycosylase. Nucleic Acids Res. 2004; 32(17): 5291–5302.
- ^ Wu P, Qiu C, Sohail A, Zhang X, Bhagwat, AS, Xiaodong C. (2003). Mismatch Repair in Methylated DNA. STRUCTURE AND ACTIVITY OF THE MISMATCH-SPECIFIC THYMINE GLYCOSYLASE DOMAIN OF METHYL-CpG-BINDING PROTEIN MBD4. 5285-5291.
- ^ Wong E, Yang K, Kuraguchi M, Werling U, Avdievich E, Fan K, Fazzari M, Jin B, Brown, M.C, Lipkin M, Edelmann W. (1995). Mbd4 inactivation increases C→T transition mutations and promotes gastrointestinal tumor formation. PNAS 99(23): 14937-14942.
- ^ Mol CD, Arvai AS, Slupphaug G, Kavli B, Alseth I, Krokan HE, Tainer JA. (1995). Crystal structure and mutational analysis of human uracil-DNA glycosylase. Cell 80(6):869-878.
- ^ Slupphaug G, Mol CD, Kavli B, Arvai AS, Krokan HE, Tainer JA. (1996). A nucleotide-flipping mechanism from the structure of human uracil–DNA glycosylase bound to DNA. 384: 87-92.
- ^ Klungland A, Rosewell I, Hollenbach S, Larsen E, Daly G, Epe A, Seeberg E, Lindahl T, Barnes D. E. (1999). Accumulation of premutagenic DNA lesions in mice defective in removal of oxidative base damage. PNAS 96(23): 13300–13305.
- ^ Russo M.T, De luca G, Degan P, Parlanti E, Dogliotti E Barnes D.E, Lindahl T, Yang H, Miller J. H, Bignami M. (2004). Accumulation of the Oxidative Base Lesion 8-Hydroxyguanine in DNA of Tumor-Prone Mice Defective in Both the Myh and Ogg1 DNA Glycosylases. Cancer Res 64(13): 4411-4414.
External links
Hydrolase: sugar hydrolases (EC 3.2) 3.2.1: Glycoside hydrolases Cellulase · Alpha-glucosidase (Acid, Neutral AB, Neutral C) · Beta-glucosidase (cytosolic) · Debranching enzymeOtherAmylase (Alpha-Amylase) · Chitinase · Lysozyme · Neuraminidase (NEU1, NEU2, NEU3, NEU4, Bacterial neuraminidase, Viral neuraminidase) · Galactosidases (Alpha, Beta) · alpha-Mannosidase · Glucuronidase · Hyaluronidase · Pullulanase · Glucosylceramidase (lysosomal, non-lysosomal) · Galactosylceramidase · Alpha-N-acetylgalactosaminidase (NAGA) · Alpha-N-acetylglucosaminidase · Fucosidase · Hexosaminidase (HEXA, HEXB) · Iduronidase · Maltase-glucoamylase · Heparanase (HPSE2)3.2.2: Hydrolysing
N-Glycosyl compoundsDNA glycosylases: Oxoguanine glycosylaseCategories:- DNA repair
- EC 3.2.2


Wikimedia Foundation. 2010.