- Carbohydrate-binding module
-
CBM_1 
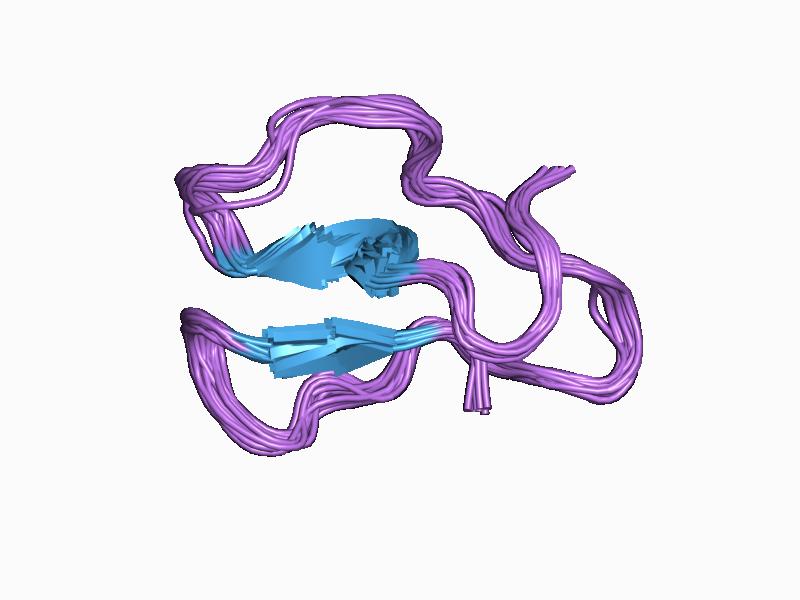
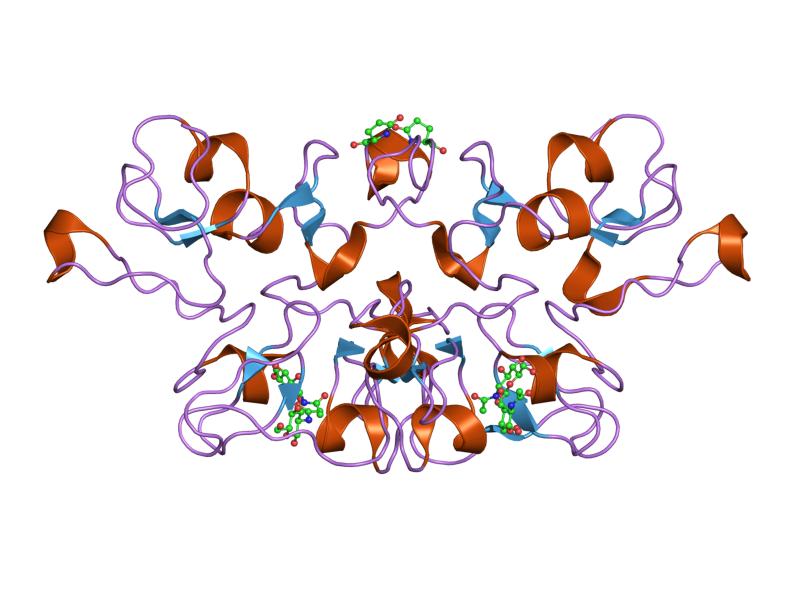
three-dimensional structures of three engineered cellulose-binding domains of cellobiohydrolase i from trichoderma reesei, nmr, 18 structures Identifiers Symbol CBM_1 Pfam PF00734 InterPro IPR000254 PROSITE PDOC00486 SCOP 1cel CAZy CBM1 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_2 
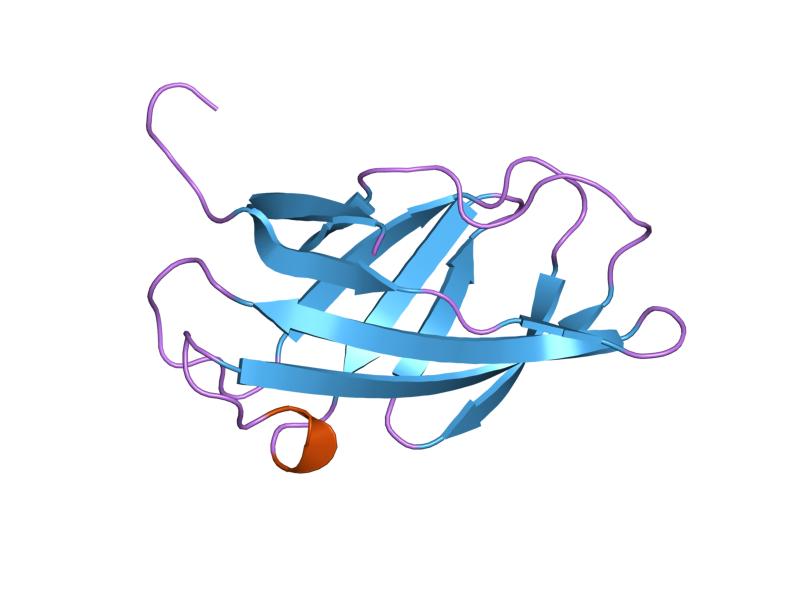
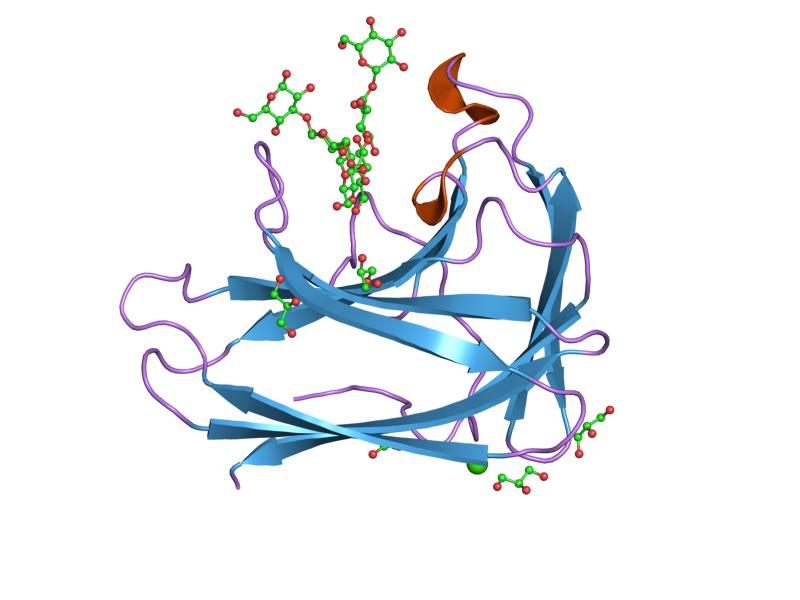
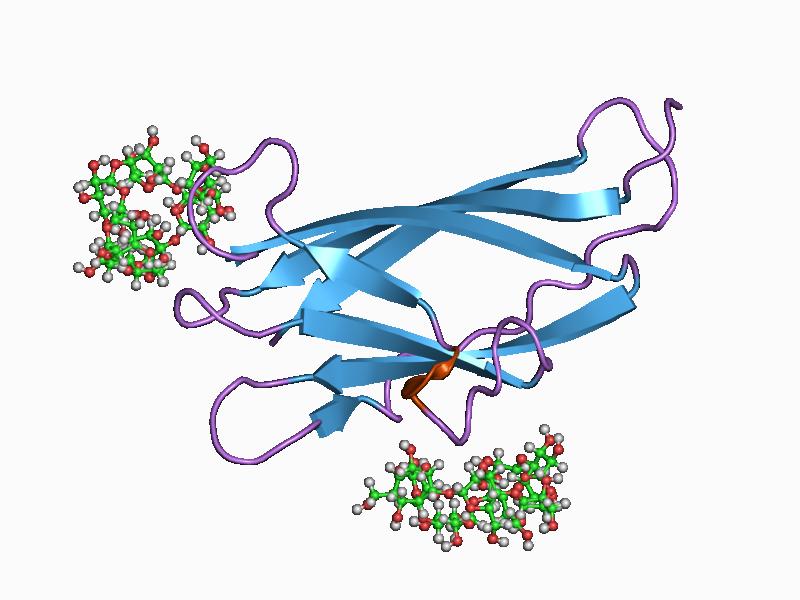
solution structure of a cellulose binding domain from cellulomonas fimi by nuclear magnetic resonance spectroscopy Identifiers Symbol CBM_2 Pfam PF00553 Pfam clan CL0203 InterPro IPR001919 PROSITE PDOC00485 SCOP 1exg CAZy CBM2 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_3 
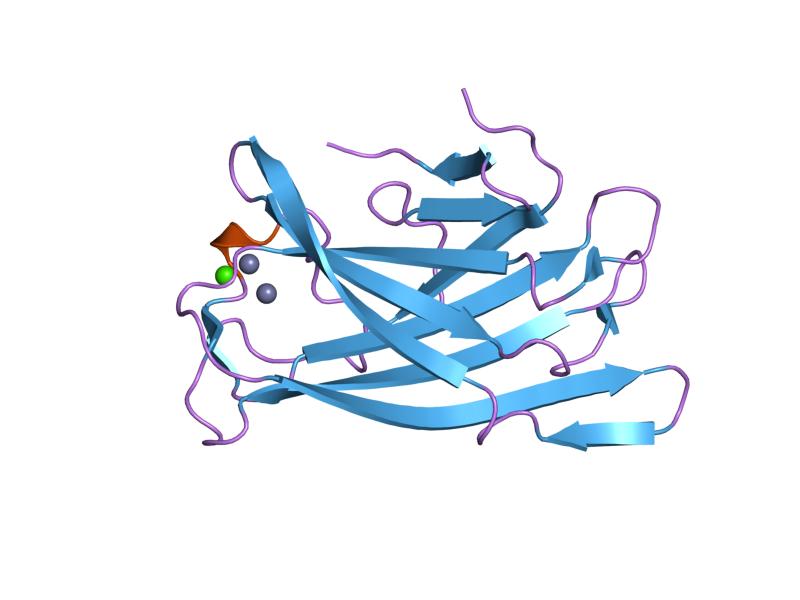
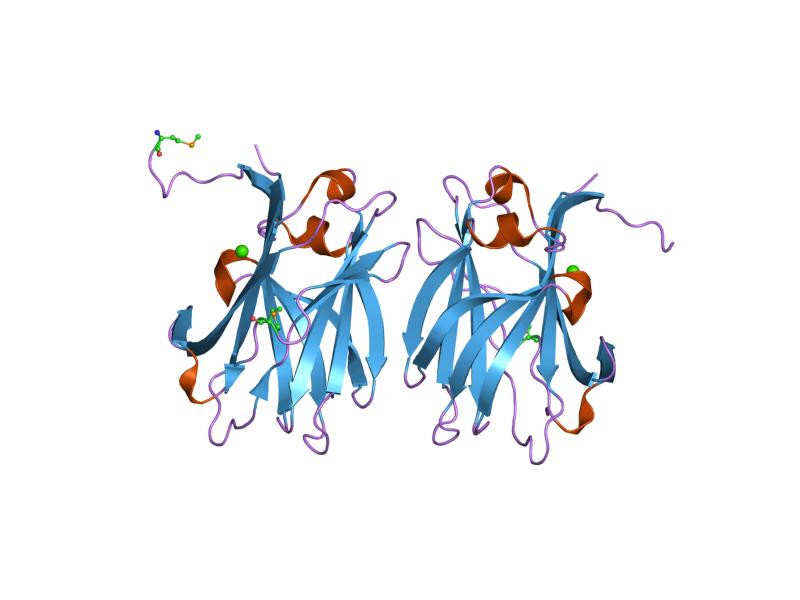
crystal structure of a family iiia cbd from clostridium cellulolyticum Identifiers Symbol CBM_3 Pfam PF00942 Pfam clan CL0203 InterPro IPR001956 SCOP 1nbc CAZy CBM3 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_5/12 
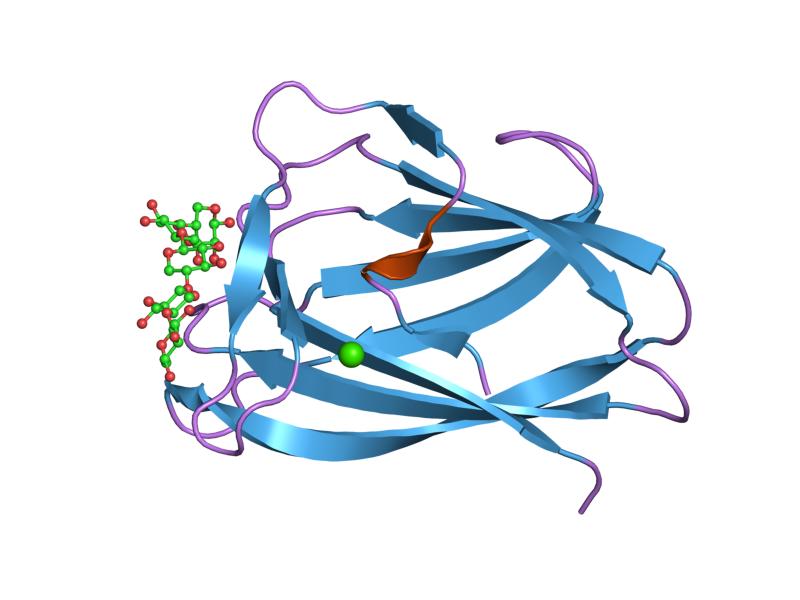
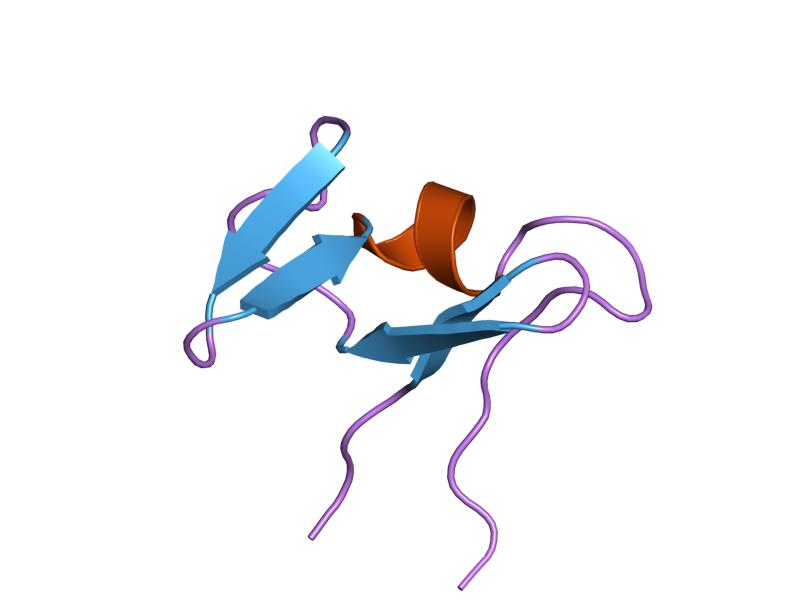
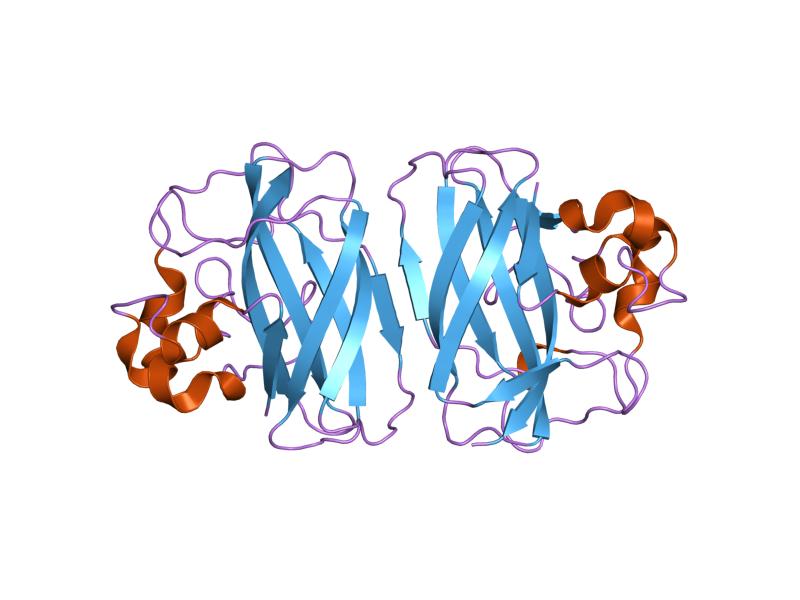
interactions of a family 18 chitinase with the designed inhibitor hm508, and its degradation product, chitobiono-delta-lactone Identifiers Symbol CBM_5_12 Pfam PF02839 InterPro IPR003610 SCOP 1ed7 CAZy CBM12 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_6 
cbm6ct from clostridium thermocellum in complex with xylopentaose Identifiers Symbol CBM_6 Pfam PF03422 Pfam clan CL0202 InterPro IPR005084 SCOP 1gmm CAZy CBM6 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_4/9 
cbm4 structure and function Identifiers Symbol CBM_4_9 Pfam PF02018 Pfam clan CL0202 InterPro IPR003305 SCOP 1ulp CAZy CBM22 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_10 
solution structure of type x cbm Identifiers Symbol CBM_10 Pfam PF02013 InterPro IPR002883 SCOP 1qld CAZy CBM10 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_11 
family 11 carbohydrate-binding module of cellulosomal cellulase lic26a-cel5e of clostridium thermocellum Identifiers Symbol CBM_11 Pfam PF03425 Pfam clan CL0202 InterPro IPR005087 CAZy CBM11 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_14 Identifiers Symbol CBM_14 Pfam PF01607 Pfam clan CL0155 InterPro IPR002557 SCOP 1dqc CAZy CBM14 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_15 
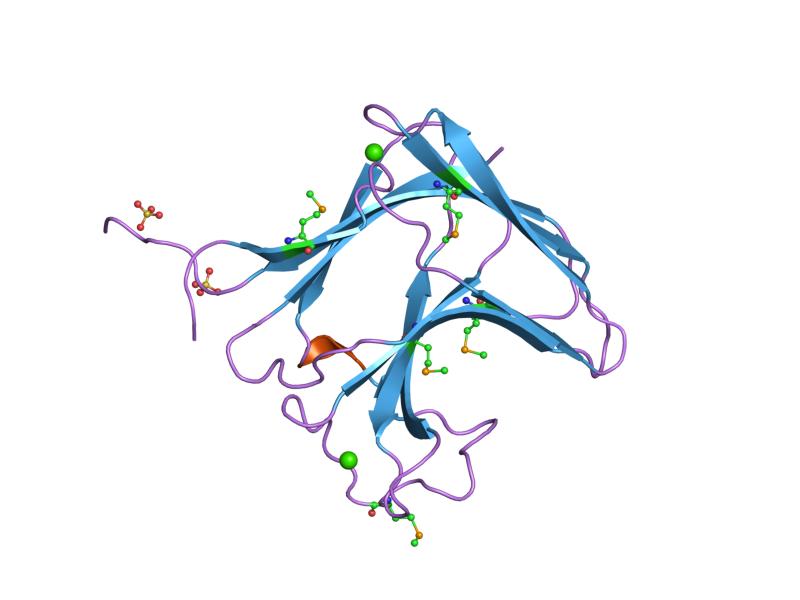
xylan-binding module cbm15 Identifiers Symbol CBM_15 Pfam PF03426 Pfam clan CL0202 InterPro IPR005088 SCOP 1gny CAZy CBM15 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_17/28 
structure of fam17 carbohydrate binding module from clostridium cellulovorans Identifiers Symbol CBM_17_28 Pfam PF03424 Pfam clan CL0202 InterPro IPR005086 SCOP 1g0c CAZy CBM28 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary Chitin_bind_1 (CBM18) 
crystal structure analysis of crosslinked-wga3/glcnacbeta1,4glcnac complex Identifiers Symbol Chitin_bind_1 Pfam PF00187 InterPro IPR001002 PROSITE PDOC00025 SCOP 1wgt CAZy CBM18 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_19 Identifiers Symbol CBM_19 Pfam PF03427 Pfam clan CL0155 InterPro IPR005089 CAZy CBM19 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_20 
glucoamylase, granular starch-binding domain complex with cyclodextrin, nmr, minimized average structure Identifiers Symbol CBM_20 Pfam PF00686 Pfam clan CL0369 InterPro IPR002044 SCOP 1cdg CAZy CBM20 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_21 Identifiers Symbol CBM_21 Pfam PF03370 InterPro IPR005036 CAZy CBM21 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_25 Identifiers Symbol CBM_25 Pfam PF03423 InterPro IPR005085 CAZy CBM25 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM27 
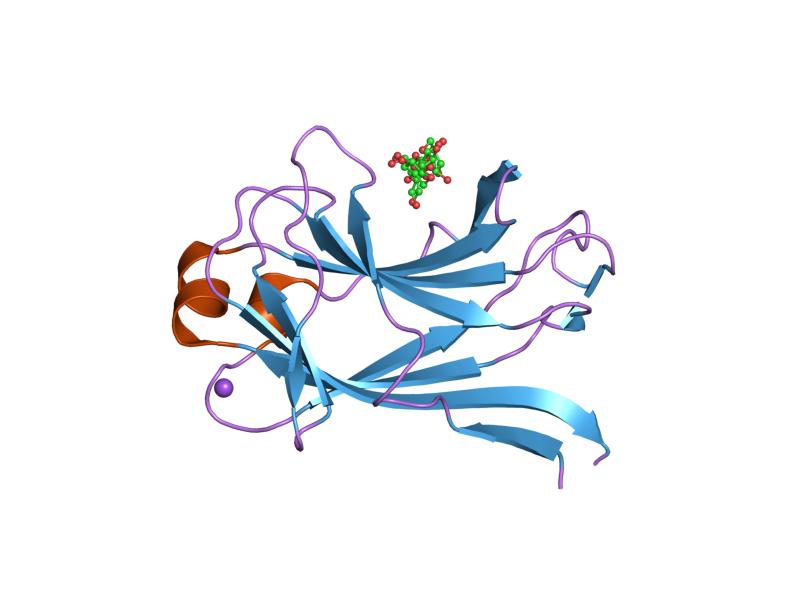
structural and thermodynamic dissection of specific mannan recognition by a carbohydrate-binding module, tmcbm27 Identifiers Symbol CBM27 Pfam PF09212 InterPro IPR015295 SCOP 1oh4 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary Chitin_bind_3 (CBM33) 
crystal structure of the serratia marcescens chitin-binding protein cbp21 y54a mutant. Identifiers Symbol Chitin_bind_3 Pfam PF03067 InterPro IPR004302 CAZy CBM33 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM_48 
crystal structure of glycosyltrehalose trehalohydrolase from sulfolobus solfataricus Identifiers Symbol CBM_48 Pfam PF02922 Pfam clan CL0369 InterPro IPR004193 SCOP 1bf2 CAZy CBM48 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary CBM49 Identifiers Symbol CBM49 Pfam PF09478 Pfam clan CL0203 InterPro IPR019028 Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary In molecular biology, a carbohydrate-binding module (CBM) is a protein domain found in carbohydrate-active enzymes (for example glycoside hydrolases). The majority of these domains have carbohydrate-binding activity. Some of these domains are found on cellulosomal scaffoldin proteins. CBMs were previously known as cellulose-binding domains.[1] CBMs are classified into numerous families, based on amino acid sequence similarity. There are currently (June 2011) 64 families of CBM in the CAZy database.[2]
CBMs of microbial glycoside hydrolases play a central role in the recycling of photosynthetically fixed carbon through their binding to specific plant structural polysaccharides.[3] CBMs can recognise both crystalline and amorphous cellulose forms.[4] CBMs are the most common non-catalytic modules associated with enzymes active in plant cell-wall hydrolysis. Many putative CBMs have been identified by amino acid sequence alignments but only a few representatives have been show experimentally to have a carbohydrate-binding function.[5]
Contents
CBM1
Carbohydrate-binding module family 1 (CBM1) consists of 36 amino acids. This domain contains 4 conserved cysteine residues which are involved in the formation of two disulfide bonds.
CBM2
Carbohydrate-binding module family 2 (CBM2) contains two conserved cysteines - one at each extremity of the domain - which have been shown [6] to be involved in a disulfide bond. There are also four conserved tryptophans, two of which are involved in cellulose binding.[7][8][9]
CBM3
Carbohydrate-binding module family 3 (CBM3) is involved in cellulose binding [10] and is found associated with a wide range of bacterial glycosyl hydrolases. The structure of this domain is known; it forms a beta sandwich.[11]
CBM4
Carbohydrate-binding module family 4 (CBM4) includes the two cellulose-binding domains, CBD(N1) and CBD(N2), arranged in tandem at the N terminus of the 1,4-beta-glucanase, CenC, from Cellulomonas fimi. These homologous CBMs are distinct in their selectivity for binding amorphous and not crystalline cellulose.[12] Multidimensional heteronuclear nuclear magnetic resonance (NMR) spectroscopy was used to determine the tertiary structure of the 152 amino acid N-terminal cellulose-binding domain from C. fimi 1,4-beta-glucanase CenC (CBDN1). The tertiary structure of CBDN1 is strikingly similar to that of the bacterial 1,3-1,4-beta-glucanases, as well as other sugar-binding proteins with jelly-roll folds.[13] CBM4 and CBM9 are closely related.
CBM5
Carbohydrate-binding module family 5 (CBM5) binds chitin.[14] CBM5 and CBM12 are distantly related.
CBM6
Carbohydrate-binding module family 6 (CBM6) is unusual in that is contains two substrate-binding sites, cleft A and cleft B. Cellvibrio mixtus endoglucanase 5A contains two CBM6 domains, the CBM6 domain at the C-terminus displays distinct ligand binding specificities in each of the sustrate-binding clefts. Both cleft A and cleft B can bind cello-oligosaccharides, laminarin preferentially binds in cleft A, xylooligosaccharides only bind in cleft A and beta1,4,-beta1,3-mixed linked glucans only bind in cleft B.[15]
CBM9
Carbohydrate-binding module family 9 (CBM9) binds to crystalline cellulose.[16] CBM4 and CBM9 are closely related.
CBM10
Carbohydrate-binding module family 10 (CBM10) is found in two distinct sets of proteins with different functions. Those found in aerobic bacteria bind cellulose (or other carbohydrates); but in anaerobic fungi they are protein binding domains, referred to as dockerin domains. The dockerin domains are believed to be responsible for the assembly of a multiprotein cellulase/hemicellulase complex, similar to the cellulosome found in certain anaerobic bacteria.[17][18]
In anaerobic bacteria that degrade plant cell walls, exemplified by Clostridium thermocellum, the dockerin domains of the catalytic polypeptides can bind equally well to any cohesin from the same organism. More recently, anaerobic fungi, typified by Piromyces equi, have been suggested to also synthesise a cellulosome complex, although the dockerin sequences of the bacterial and fungal enzymes are completely different.[19] For example, the fungal enzymes contain one, two or three copies of the dockerin sequence in tandem within the catalytic polypeptide. In contrast, all the C. thermocellum cellulosome catalytic components contain a single dockerin domain. The anaerobic bacterial dockerins are homologous to EF hands (calcium-binding motifs) and require calcium for activity whereas the fungal dockerin does not require calcium. Finally, the interaction between cohesin and dockerin appears to be species specific in bacteria, there is almost no species specificity of binding within fungal species and no identified sites that distinguish different species.
The of dockerin from P. equi contains two helical stretches and four short beta-strands which form an antiparallel sheet structure adjacent to an additional short twisted parallel strand. The N- and C-termini are adjacent to each other.[19]
CBM11
Carbohydrate-binding module family 11 (CBM11) is found in a number of bacterial cellulases. One example is the CBM11 of Clotridium thermocellum Cel26A-Cel5E, this domain has been shown to bind both β-1,4-glucan and β-1,3-1,4-mixed linked glucans.[20] CBM11 has beta-sandwich structure with a concave side forming a substrate-binding cleft. [20]
CBM12
Carbohydrate-binding module family 12 (CBM12) comprises two beta-sheets, consisting of two and three antiparallel beta strands respectively. It binds chitin via the aromatic rings of tryptophan residues.[14] CBM5 and CBM12 are distantly related.
CBM14
Carbohydrate-binding module family 14 (CBM14) is also known as the peritrophin-A domain. It is found in chitin binding proteins, particularly the peritrophic matrix proteins of insects and animal chitinases.[21][22][23] Copies of the domain are also found in some baculoviruses. It is an extracellular domain that contains six conserved cysteines that probably form three disulfide bridges. Chitin binding has been demonstrated for a protein containing only two of these domains.[21]
CBM15
Carbohydrate-binding module family 15 (CBM15), found in bacterial enzymes, has been shown to bind to xylan and xylooligosaccharides. It has a beta-jelly roll fold, with a groove on the concave surface of one of the beta-sheets.[24]
CBM17
Carbohydrate-binding module family 17 (CBM17) appears to have a very shallow binding cleft that may be more accessible to cellulose chains in non-crystalline cellulose than the deeper binding clefts of family 4 CBMs.[25] Sequence and structural conservation in families CBM17 and CBM28 suggests that they have evolved through gene duplication and subsequent divergence.[4] CBM17 does not compete with CBM28 modules when binding to non-crystalline cellulose. Different CBMs have been shown to bind to different sirtes in amorphous cellulose, CBM17 and CBM28 recognise distinct non-overlapping sites in amorphous cellulose.[26]
CBM18
Carbohydrate-binding module family 18 (CBM18) (also known as chitin binding 1 or chitin recognition protein) is found in a number of plant and fungal proteins that bind N-acetylglucosamine (e.g. solanaceous lectins of tomato and potato, plant endochitinases, the wound-induced proteins: hevein, win1 and win2, and the Kluyveromyces lactis killer toxin alpha subunit).[27] The domain may occur in one or more copies and is thought to be involved in recognition or binding of chitin subunits.[28][29] In chitinases, as well as in the potato wound-induced proteins, this 43-residue domain directly follows the signal sequence and is therefore at the N terminus of the mature protein; in the killer toxin alpha subunit it is located in the central section of the protein.
CBM19
Carbohydrate-binding module family 19 (CBM19), found in fungal chitinases, binds chitin.[30]
CBM20
Carbohydrate-binding module family 20 (CBM20) binds to starch.[31][32]
CBM21
Carbohydrate-binding module family 21 (CBM21), found in many eukaryotic proteins involved in glycogen metabolism, binds to glycogen.[33]
CBM25
Carbohydrate-binding module family 25 (CBM25) binds alpha-glucooligosaccharides, particularly those containing alpha-1,6 linkages, and granular starch.[34]
CBM27
Carbohydrate-binding module family 27 (CBM27) binds to beta-1,4-mannooligosaccharides, carob galactomannan, and konjac glucomannan, but not to cellulose (insoluble and soluble) or soluble birchwood xylan. CBM27 adopts a beta sandwich structure comprising 13 beta strands with a single, small alpha-helix and a single metal atom.[35]
CBM28
Carbohydrate-binding module family 28 (CBM28) does not compete with CBM17 modules when binding to non-crystalline cellulose. Different CBMs have been shown to bind to different sirtes in amorphous cellulose, CBM17 and CBM28 recognise distinct non-overlapping sites in amorphous cellulose. CBM28 has a "beta-jelly roll" topology, which is similar in structure to the CBM17 domains. Sequence and structural conservation in families CBM17 and CBM28 suggests that they have evolved through gene duplication and subsequent divergence.[4][26]
CBM33
Carbohydrate-binding module family 33 (CBM33) is a chitin-binding domain.[36] It has a budded fibronectin type III fold consisting of two beta-sheets, arranged as a beta-sheet sandwich and a bud consisting of three short helices, located between beta-strands 1 and 2. It binds chitin via conserved polar amino acids.[37] This domain is found in isolation in baculoviral spheroidin and spindolin proteins.
CBM48
Carbohydrate-binding module family 48 (CBM48) is often found in enzymes containing glycosyl hydrolase family 13 catalytic domains. It is found in a range of enzymes that act on branched substrates ie. isoamylase, pullulanase and branching enzyme. Isoamylase hydrolyses 1,6-alpha-D-glucosidic branch linkages in glycogen, amylopectin and dextrin; 1,4-alpha-glucan branching enzyme functions in the formation of 1,6-glucosidic linkages of glycogen; and pullulanase is a starch-debranching enzyme. CBM48 binds glycogen.[38][39][40][41]
CBM49
Carbohydrate-binding module family 49 (CBM49) is found at the C-terminal of cellulases and in vitro binding studies have shown it to binds to crystalline cellulose.[42]
References
- ^ Gilkes NR, Henrissat B, Kilburn DG, Miller RC, Warren RA (June 1991). "Domains in microbial beta-1, 4-glycanases: sequence conservation, function, and enzyme families". Microbiol. Rev. 55 (2): 303–15. PMC 372816. PMID 1886523. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=372816.
- ^ Cantarel, B. L.; Coutinho, P. M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. (2009). "The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics". Nucleic Acids Research 37 (Database issue): D233–D238. doi:10.1093/nar/gkn663. PMC 2686590. PMID 18838391. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2686590.
- ^ Szabo L, Jamal S, Xie H, Charnock SJ, Bolam DN, Gilbert HJ, Davies GJ (December 2001). "Structure of a family 15 carbohydrate-binding module in complex with xylopentaose. Evidence that xylan binds in an approximate 3-fold helical conformation". J. Biol. Chem. 276 (52): 49061–5. doi:10.1074/jbc.M109558200. PMID 11598143.
- ^ a b c Jamal S, Nurizzo D, Boraston AB, Davies GJ (May 2004). "X-ray crystal structure of a non-crystalline cellulose-specific carbohydrate-binding module: CBM28". J. Mol. Biol. 339 (2): 253–8. doi:10.1016/j.jmb.2004.03.069. PMID 15136030.
- ^ Roske Y, Sunna A, Pfeil W, Heinemann U (July 2004). "High-resolution crystal structures of Caldicellulosiruptor strain Rt8B.4 carbohydrate-binding module CBM27-1 and its complex with mannohexaose". J. Mol. Biol. 340 (3): 543–54. doi:10.1016/j.jmb.2004.04.072. PMID 15210353.
- ^ Gilkes NR, Claeyssens M, Aebersold R, Henrissat B, Meinke A, Morrison HD, Kilburn DG, Warren RA, Miller RC (December 1991). "Structural and functional relationships in two families of beta-1,4-glycanases". Eur. J. Biochem. 202 (2): 367–77. doi:10.1111/j.1432-1033.1991.tb16384.x. PMID 1761039.
- ^ Meinke A, Gilkes NR, Kilburn DG, Miller RC, Warren RA (December 1991). "Bacterial cellulose-binding domain-like sequences in eucaryotic polypeptides". Protein Seq. Data Anal. 4 (6): 349–53. PMID 1812490.
- ^ Simpson PJ, Xie H, Bolam DN, Gilbert HJ, Williamson MP (December 2000). "The structural basis for the ligand specificity of family 2 carbohydrate-binding modules". J. Biol. Chem. 275 (52): 41137–42. doi:10.1074/jbc.M006948200. PMID 10973978.
- ^ Xu, G. Y.; Ong, E.; Gilkes, N. R.; Kilburn, D. G.; Muhandiram, D. R.; Harris-Brandts, M.; Carver, J. P.; Kay, L. E. et al. (1995). "Solution structure of a cellulose-binding domain from Cellulomonas fimi by nuclear magnetic resonance spectroscopy". Biochemistry 34 (21): 6993–7009. doi:10.1021/bi00021a011. PMID 7766609.
- ^ Poole DM, Morag E, Lamed R, Bayer EA, Hazlewood GP, Gilbert HJ (December 1992). "Identification of the cellulose-binding domain of the cellulosome subunit S1 from Clostridium thermocellum YS". FEMS Microbiol. Lett. 78 (2-3): 181–6. PMID 1490597.
- ^ Tormo J, Lamed R, Chirino AJ, Morag E, Bayer EA, Shoham Y, Steitz TA (November 1996). "Crystal structure of a bacterial family-III cellulose-binding domain: a general mechanism for attachment to cellulose". EMBO J. 15 (21): 5739–51. PMC 452321. PMID 8918451. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=452321.
- ^ Brun E, Johnson PE, Creagh AL, Tomme P, Webster P, Haynes CA, McIntosh LP (March 2000). "Structure and binding specificity of the second N-terminal cellulose-binding domain from Cellulomonas fimi endoglucanase C". Biochemistry 39 (10): 2445–58. doi:10.1021/bi992079u. PMID 10704194.
- ^ Johnson PE, Joshi MD, Tomme P, Kilburn DG, McIntosh LP (November 1996). "Structure of the N-terminal cellulose-binding domain of Cellulomonas fimi CenC determined by nuclear magnetic resonance spectroscopy". Biochemistry 35 (45): 14381–94. doi:10.1021/bi961612s. PMID 8916925.
- ^ a b Akagi, K. -I.; Watanabe, J.; Hara, M.; Kezuka, Y.; Chikaishi, E.; Yamaguchi, T.; Akutsu, H.; Nonaka, T. et al. (2006). "Identification of the Substrate Interaction Region of the Chitin-Binding Domain of Streptomyces griseus Chitinase C". Journal of Biochemistry 139 (3): 483–493. doi:10.1093/jb/mvj062. PMID 16567413.
- ^ Henshaw, J. L.; Bolam, D. N.; Pires, V. M.; Czjzek, M.; Henrissat, B.; Ferreira, L. M.; Fontes, C. M.; Gilbert, H. J. (2004). "The Family 6 Carbohydrate Binding Module CmCBM6-2 Contains Two Ligand-binding Sites with Distinct Specificities". Journal of Biological Chemistry 279 (20): 21552–21559. doi:10.1074/jbc.M401620200. PMID 15004011.
- ^ Winterhalter, C.; Heinrich, P.; Candussio, A.; Wich, G.; Liebl, W. (1995). "Identification of a novel cellulose-binding domain within the multidomain 120 kDa xylanase XynA of the hyperthermophilic bacterium Thermotoga maritima". Molecular microbiology 15 (3): 431–444. doi:10.1111/j.1365-2958.1995.tb02257.x. PMID 7783614.
- ^ Millward-Sadler SJ, Davidson K, Hazlewood GP, Black GW, Gilbert HJ, Clarke JH (November 1995). "Novel cellulose-binding domains, NodB homologues and conserved modular architecture in xylanases from the aerobic soil bacteria Pseudomonas fluorescens subsp. cellulosa and Cellvibrio mixtus". Biochem. J. 312 ( Pt 1): 39–48. PMC 1136224. PMID 7492333. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1136224.
- ^ Fanutti C, Ponyi T, Black GW, Hazlewood GP, Gilbert HJ (December 1995). "The conserved noncatalytic 40-residue sequence in cellulases and hemicellulases from anaerobic fungi functions as a protein docking domain". J. Biol. Chem. 270 (49): 29314–22. PMID 7493964.
- ^ a b Raghothama S, Eberhardt RY, Simpson P, Wigelsworth D, White P, Hazlewood GP, Nagy T, Gilbert HJ, Williamson MP (September 2001). "Characterization of a cellulosome dockerin domain from the anaerobic fungus Piromyces equi". Nat. Struct. Biol. 8 (9): 775–8. doi:10.1038/nsb0901-775. PMID 11524680.
- ^ a b Carvalho, A. L.; Goyal, A.; Prates, J. A.; Bolam, D. N.; Gilbert, H. J.; Pires, V. M.; Ferreira, L. M.; Planas, A. et al. (2004). "The Family 11 Carbohydrate-binding Module of Clostridium thermocellum Lic26A-Cel5E Accommodates -1,4- and -1,3-1,4-Mixed Linked Glucans at a Single Binding Site". Journal of Biological Chemistry 279 (33): 34785–34793. doi:10.1074/jbc.M405867200. PMID 15192099.
- ^ a b Shen Z, Jacobs-Lorena M (July 1998). "A type I peritrophic matrix protein from the malaria vector Anopheles gambiae binds to chitin. Cloning, expression, and characterization". J. Biol. Chem. 273 (28): 17665–70. PMID 9651363.
- ^ Elvin CM, Vuocolo T, Pearson RD, East IJ, Riding GA, Eisemann CH, Tellam RL (April 1996). "Characterization of a major peritrophic membrane protein, peritrophin-44, from the larvae of Lucilia cuprina. cDNA and deduced amino acid sequences". J. Biol. Chem. 271 (15): 8925–35. PMID 8621536.
- ^ Casu R, Eisemann C, Pearson R, Riding G, East I, Donaldson A, Cadogan L, Tellam R (August 1997). "Antibody-mediated inhibition of the growth of larvae from an insect causing cutaneous myiasis in a mammalian host". Proc. Natl. Acad. Sci. U.S.A. 94 (17): 8939–44. doi:10.1073/pnas.94.17.8939. PMC 22971. PMID 9256413. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=22971.
- ^ Szabo, L.; Jamal, S.; Xie, H.; Charnock, S. J.; Bolam, D. N.; Gilbert, H. J.; Davies, G. J. (2001). "Structure of a Family 15 Carbohydrate-binding Module in Complex with Xylopentaose. EVIDENCE THAT XYLAN BINDS IN AN APPROXIMATE 3-FOLD HELICAL CONFORMATION". Journal of Biological Chemistry 276 (52): 49061–49065. doi:10.1074/jbc.M109558200. PMID 11598143.
- ^ Notenboom V, Boraston AB, Chiu P, Freelove AC, Kilburn DG, Rose DR (December 2001). "Recognition of cello-oligosaccharides by a family 17 carbohydrate-binding module: an X-ray crystallographic, thermodynamic and mutagenic study". J. Mol. Biol. 314 (4): 797–806. doi:10.1006/jmbi.2001.5153. PMID 11733998.
- ^ a b Jamal, S.; Nurizzo, D.; Boraston, A. B.; Davies, G. J. (2004). "X-ray Crystal Structure of a Non-crystalline Cellulose-specific Carbohydrate-binding Module: CBM28". Journal of Molecular Biology 339 (2): 253–258. doi:10.1016/j.jmb.2004.03.069. PMID 15136030.
- ^ Wright HT, Sandrasegaram G, Wright CS (September 1991). "Evolution of a family of N-acetylglucosamine binding proteins containing the disulfide-rich domain of wheat germ agglutinin". J. Mol. Evol. 33 (3): 283–94. PMID 1757999.
- ^ Butler AR, O'Donnell RW, Martin VJ, Gooday GW, Stark MJ (July 1991). "Kluyveromyces lactis toxin has an essential chitinase activity". Eur. J. Biochem. 199 (2): 483–8. doi:10.1111/j.1432-1033.1991.tb16147.x. PMID 2070799.
- ^ Lerner DR, Raikhel NV (June 1992). "The gene for stinging nettle lectin (Urtica dioica agglutinin) encodes both a lectin and a chitinase". J. Biol. Chem. 267 (16): 11085–91. PMID 1375935.
- ^ Kuranda, M. J.; Robbins, P. W. (1991). "Chitinase is required for cell separation during growth of Saccharomyces cerevisiae". The Journal of biological chemistry 266 (29): 19758–19767. PMID 1918080.
- ^ Penninga, D.; Van Der Veen, B. A.; Knegtel, R. M.; Van Hijum, S. A.; Rozeboom, H. J.; Kalk, K. H.; Dijkstra, B. W.; Dijkhuizen, L. (1996). "The raw starch binding domain of cyclodextrin glycosyltransferase from Bacillus circulans strain 251". The Journal of biological chemistry 271 (51): 32777–32784. PMID 8955113.
- ^ Oyama, T.; Kusunoki, M.; Kishimoto, Y.; Takasaki, Y.; Nitta, Y. (1999). "Crystal structure of beta-amylase from Bacillus cereus var. Mycoides at 2.2 a resolution". Journal of biochemistry 125 (6): 1120–1130. PMID 10348915.
- ^ Armstrong, C. G.; Doherty, M. J.; Cohen, P. T. (1998). "Identification of the separate domains in the hepatic glycogen-targeting subunit of protein phosphatase 1 that interact with phosphorylase a, glycogen and protein phosphatase 1". The Biochemical journal 336 ( Pt 3): 699–704. PMC 1219922. PMID 9841883. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1219922.
- ^ Boraston, A. B.; Healey, M.; Klassen, J.; Ficko-Blean, E.; Lammerts Van Bueren, A.; Law, V. (2005). "A Structural and Functional Analysis of -Glucan Recognition by Family 25 and 26 Carbohydrate-binding Modules Reveals a Conserved Mode of Starch Recognition". Journal of Biological Chemistry 281 (1): 587–598. doi:10.1074/jbc.M509958200. PMID 16230347.
- ^ Boraston AB, Revett TJ, Boraston CM, Nurizzo D, Davies GJ (June 2003). "Structural and thermodynamic dissection of specific mannan recognition by a carbohydrate binding module, TmCBM27". Structure 11 (6): 665–75. doi:10.1016/S0969-2126(03)00100-X. PMID 12791255.
- ^ Schnellmann, J.; Zeltins, A.; Blaak, H.; Schrempf, H. (1994). "The novel lectin-like protein CHB1 is encoded by a chitin-inducible Streptomyces olivaceoviridis gene and binds specifically to crystalline alpha-chitin of fungi and other organisms". Molecular microbiology 13 (5): 807–819. doi:10.1111/j.1365-2958.1994.tb00473.x. PMID 7815940.
- ^ Vaaje-Kolstad, G.; Houston, D. R.; Riemen, A. H.; Eijsink, V. G.; Van Aalten, D. M. (2005). "Crystal Structure and Binding Properties of the Serratia marcescens Chitin-binding Protein CBP21". Journal of Biological Chemistry 280 (12): 11313–11319. doi:10.1074/jbc.M407175200. PMID 15590674.
- ^ Katsuya, Y.; Mezaki, Y.; Kubota, M.; Matsuura, Y. (1998). "Three-dimensional structure of Pseudomonas isoamylase at 2.2 Å resolution1". Journal of Molecular Biology 281 (5): 885–897. doi:10.1006/jmbi.1998.1992. PMID 9719642.
- ^ Wiatrowski, H. A.; Van Denderen, B. J.; Berkey, C. D.; Kemp, B. E.; Stapleton, D.; Carlson, M. (2004). "Mutations in the gal83 glycogen-binding domain activate the snf1/gal83 kinase pathway by a glycogen-independent mechanism". Molecular and cellular biology 24 (1): 352–361. PMC 303368. PMID 14673168. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=303368.
- ^ Polekhina, G.; Gupta, A.; Michell, B. J.; Van Denderen, B.; Murthy, S.; Feil, S. C.; Jennings, I. G.; Campbell, D. J. et al. (2003). "AMPK beta subunit targets metabolic stress sensing to glycogen". Current biology : CB 13 (10): 867–871. doi:10.1016/S0960-9822(03)00292-6. PMID 12747837.
- ^ Hudson, E. R.; Pan, D. A.; James, J.; Lucocq, J. M.; Hawley, S. A.; Green, K. A.; Baba, O.; Terashima, T. et al. (2003). "A novel domain in AMP-activated protein kinase causes glycogen storage bodies similar to those seen in hereditary cardiac arrhythmias". Current biology : CB 13 (10): 861–866. doi:10.1016/S0960-9822(03)00249-5. PMID 12747836.
- ^ Urbanowicz BR, Catala C, Irwin D, Wilson DB, Ripoll DR, Rose JK (April 2007). "A tomato endo-beta-1,4-glucanase, SlCel9C1, represents a distinct subclass with a new family of carbohydrate binding modules (CBM49)". J. Biol. Chem. 282 (16): 12066–74. doi:10.1074/jbc.M607925200. PMID 17322304.
External links
This article includes text from the public domain Pfam and InterPro IPR000254
This article includes text from the public domain Pfam and InterPro IPR002883
This article includes text from the public domain Pfam and InterPro IPR005087
This article includes text from the public domain Pfam and InterPro IPR002557
This article includes text from the public domain Pfam and InterPro IPR005088
This article includes text from the public domain Pfam and InterPro IPR005086
This article includes text from the public domain Pfam and InterPro IPR005089
This article includes text from the public domain Pfam and InterPro IPR001919
This article includes text from the public domain Pfam and InterPro IPR002044
This article includes text from the public domain Pfam and InterPro IPR005036
This article includes text from the public domain Pfam and InterPro IPR005085
This article includes text from the public domain Pfam and InterPro IPR015295
This article includes text from the public domain Pfam and InterPro IPR001956
This article includes text from the public domain Pfam and InterPro IPR004193
This article includes text from the public domain Pfam and InterPro IPR019028
This article includes text from the public domain Pfam and InterPro IPR003305
This article includes text from the public domain Pfam and InterPro IPR003610
This article includes text from the public domain Pfam and InterPro IPR005084
This article includes text from the public domain Pfam and InterPro IPR001002
Categories:- Protein domains


Wikimedia Foundation. 2010.