- Phloem
-
In vascular plants, phloem is the living tissue that carries organic nutrients (known as photosynthate), in particular, glucose, a sugar, to all parts of the plant where needed. In trees, the phloem is the innermost layer of the bark, hence the name, derived from the Greek word φλόος (phloos) "bark". The phloem is concerned mainly with the transport of soluble organic material made during photosynthesis. This is called translocation.
Contents
Structure
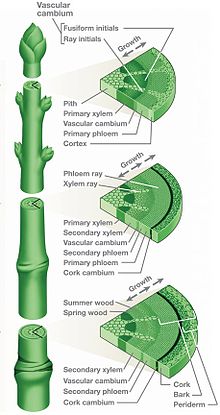 Multiple cross-sections of a stem showing phloem and companion cells[1]
Multiple cross-sections of a stem showing phloem and companion cells[1]
Phloem tissue consists of less specialized and nucleate parenchyma cells, sieve-tube cells, and companion cells (in addition albuminous cells, fibres and sclereids).
Sieve tubes
The sieve-tube cells lack a nucleus, have very few vacuoles, but contain other organelles such as ribosomes. The sieve tube is an elongated rank of individual cells, called sieve-tube members, arranged end to end. The endoplasmic reticulum is concentrated at the lateral walls. Sieve-tube members are joined end to end to form a tube that conducts food materials throughout the plant. The end walls of these cells have many small pores and are called sieve plates and have enlarged plasmodesmata.
Companion cells
The survival of sieve-tube members depends on a close association with the companion cells. All of the cellular functions of a sieve-tube element are carried out by the (much smaller) companion cell, a typical plant cell, except the companion cell usually has a larger number of ribosomes and mitochondria. This is because the companion cell is more metabolically active than a 'typical' plant cell. The cytoplasm of a companion cell is connected to the sieve-tube element by plasmodesmata.
There are three types of companion cell.
- Ordinary companions cells, which have smooth walls and few or no plasmodesmata connections to cells other than the sieve tube.
- Transfer cells, which have much folded walls that are adjacent to non-sieve cells, allowing for larger areas of transfer. They are specialised in scavenging solutes from those in the cell walls that are actively pumped requiring energy.
- Intermediary cells, which have smooth walls and numerous plasmodesmata connecting them to other cells.
The first two types of cell collect solutes through apoplastic (cell wall) transfers, whilst the third type can collect solutes via the symplast through the plasmodesmata connections.
Function
Unlike xylem (which is composed primarily of dead cells), the phloem is composed of still-living cells that transport sap. The sap is a water-based solution, but rich in sugars made by the photosynthetic areas. These sugars are transported to non-photosynthetic parts of the plant, such as the roots, or into storage structures, such as tubers or bulbs.
The Pressure flow hypothesis was a hypothesis proposed by Ernst Münch in 1930 that explained the mechanism of phloem translocation.[2] A high concentration of organic substance inside cells of the phloem at a source, such as a leaf, creates a diffusion gradient that draws water into the cells. Movement occurs by bulk flow; phloem sap moves from sugar sources to sugar sinks by means of turgor pressure gradient. A sugar source is any part of the plant that is producing or releasing sugar.
During the plant's growth period, usually during the spring, storage organs such as the roots are sugar sources, and the plant's many growing areas are sugar sinks. The movement in phloem is multidirectional, whereas, in xylem cells, it is unidirectional (upward).
After the growth period, when the meristems are dormant, the leaves are sources, and storage organs are sinks. Developing seed-bearing organs (such as fruit) are always sinks. Because of this multi-directional flow, coupled with the fact that sap cannot move with ease between adjacent sieve-tubes, it is not unusual for sap in adjacent sieve-tubes to be flowing in opposite directions.
While movement of water and minerals through the xylem is driven by negative pressures (tension) most of the time, movement through the phloem is driven by positive hydrostatic pressures. This process is termed translocation, and is accomplished by a process called phloem loading and unloading. Cells in a sugar source "load" a sieve-tube element by actively transporting solute molecules into it. This causes water to move into the sieve-tube element by osmosis, creating pressure that pushes the sap down the tube. In sugar sinks, cells actively transport solutes out of the sieve-tube elements, producing the exactly opposite effect.
Some plants however appear not to load phloem by active transport. In these cases a mechanism known as the polymer trap mechanism was proposed by Robert Turgeon.[3] In this case small sugars such as sucrose move into intermediary cells through narrow plasmodesmata, where they are polymerised to raffinose and other larger oligosaccharides. Now they are unable to move back, but can proceed through wider plasmodesmata into the sieve tube element.
The symplastic phloem loading (polymer trap mechanism above) is confined mostly to plants in tropical rain forests and is seen as more primitive. The actively-transported apoplastic phloem loading is viewed as more advanced, as it is found in the later-evolved plants, and particularly in those in temperate and arid conditions. This mechanism may, therefore, have allowed plants to colonise the cooler locations.
Organic molecules such as sugars, amino acids, certain hormones, and even messenger RNAs are transported in the phloem through sieve tube elements.
Girdling
Main article: GirdlingBecause phloem tubes sit on the outside of the xylem in most plants, a tree or other plant can be effectively killed by stripping away the bark in a ring on the trunk or stem. With the phloem destroyed, nutrients cannot reach the roots, and the tree/plant will die. Trees located in areas with animals such as beavers are vulnerable since beavers chew off the bark at a fairly precise height. This process is known as girdling, and can be used for agricultural purposes. For example, enormous fruits and vegetables seen at fairs and carnivals are produced via girdling. A farmer would place a girdle at base of a large branch, and remove all but one fruit/vegetable from that branch. Thus, all the sugars manufactured by leaves on that branch have no sinks to go to but the one fruit/vegetable, which thus expands to many times normal size.
Origin
The phloem originates, and grows outwards from, meristematic cells in the vascular cambium. Phloem is produced in phases. Primary phloem is laid down by the apical meristem and develops from the procambium. Secondary phloem is laid down by the vascular cambium to the inside of the established layer(s) of phloem.
In some eudicot families (Apocynaceae, Convolvulaceae, Cucurbitaceae, Solanaceae, Myrtaceae, Asteraceae), phloem also develops on the inner side of the vascular cambium; in this case, a distinction between external phloem and internal phloem or intraxylary phloem is made. Internal phloem is mostly primary, and begins differentiation later than the external phloem and protoxylem, though it is not without exceptions. In some other families (Amaranthaceae, Nyctaginaceae, Salvadoraceae), the cambium also periodically forms inward strands or layers of phloem, embedded in the xylem: Such phloem strands are called included phloem or interxylary phloem.[4]
Nutritional use
Phloem of pine trees has been used in Finland as a substitute food in times of famine, and even in good years in the northeast, where supplies of phloem from earlier years helped stave off starvation somewhat in the great famine of the 1860s. Phloem is dried and milled to flour (pettu in Finnish) and mixed with rye to form a hard dark bread. Recently, pettu has again become available as a curiosity, and some have made claims of health benefits.
See also
References
- ^ Winterborne J, 2005. Hydroponics - Indoor Horticulture
- ^ Münch, E (1930). "Die Stoffbewegunen in der Pflanze". Verlag von Gustav Fischer, Jena: 234.
- ^ Turgeon, R (1991). "Symplastic phloem loading and the sink-source transition in leaves: a model". In VL Bonnemain, S Delrot, J Dainty, WJ Lucas, (eds). Recent Advances Phloem Transport and Assimilate Compartmentation.
- ^ Evert, Ray F. Esau's Plant Anatomy. John Wiley & Sons, Inc, 2006, p. 357-358.
Categories:

Wikimedia Foundation. 2010.