- Angraecum sesquipedale
-
Angraecum sesquipedale 
Angraecum sesquipedale Scientific classification Kingdom: Plantae (unranked): Angiosperms (unranked): Monocots Order: Asparagales Family: Orchidaceae Subfamily: Epidendroideae Tribe: Vandaeae Subtribe: Angraecinae Alliance: Angraecum Genus: Angraecum Species: A. sesquipedale Binomial name Angraecum sesquipedale
ThouarsVarieties - A. s. var. angustifolium
- A. s. var. sesquipedale
Synonyms Angraecum sesquipedale[1] (also called Darwin's orchid, Christmas orchid, Star of Bethlehem orchid, and King of the Angraecums) is an epiphytic orchid in the genus Angraecum that is endemic to Madagascar. The orchid was first discovered by the French botanist Louis-Marie Aubert du Petit-Thouars in 1798, but was not described until 1822.[2] It is noteworthy for its long spur and its association with the naturalist Charles Darwin who surmised that the flower was pollinated by a then undiscovered moth with a proboscis whose length was then unprecedented. His prediction had gone unverified until 21 years after his death when the moth was discovered and his conjecture vindicated. The story of its postulated pollinator has come to be seen as one of the celebrated predictions of the theory of evolution.
Contents
Description
Angraecum sesquipedale is a monocot with monopodial growth and can get to a height of 1 meter (3.3 ft).[3] Its growth habit is rather similar to species in the genus Aerides.
 A. sesquipedale flowers
A. sesquipedale flowers
The leaves are dark green with a bit of a grayish tone and leathery with a bilobed tip. They are usually around 20–40 centimeters (7.9–15.7 in) long and 6–8 cm (2.4–3.1 in) wide.[3] The roots are dark gray, thick, and emerge from the orchid's stem. There tends to be few roots and they attach to the bark of the trees quite strongly.[4] Each of the succulent roots can extend along the trunk of the tree for several meters.[3]
There is also a variations of this species namely A. sesquipedale var. angustifolium.[5][6] A. sesquipedale var. angustifolium tends to be smaller than A. sesquipedale and has narrower leaves.[7] The chromosome number of A. sesquipedale is 2n=42.[8][9] A. sesquipedale has also previously gone by the synonyms Aeranthes sesquipedalis Lindl. (1824),[10] Macroplectrum sesquipedale Pfitzer (1889),[11] Angorchis sesquepedalis Kuntze (1891),[12] and Mystacidium sesquipedale Rolfe (1904).[13]
Habitat
It is often found in lowlands in Madagascar at altitudes below 400–500 ft, near the east coast of the island, and on trees that are at the edge of forests.[14][15] Usually it is attached to trees with fewer leaves and to areas of the branch or trunk that are driest.[16] This allows the plant to obtain a great deal of light and air movement. Larger plants are usually found growing within 12–20 ft from the ground, whereas smaller plants are often found higher up in the canopy.[16] Rarely A. sesquipedale is also found growing as a lithophyte and sometimes even as a semi-terrestrial.[17][18] The orchid lives in an environment with heavy rainfall, up to 150 in (380 cm) per year.[19] There is no dry season so the growing season is continuous.
Flower
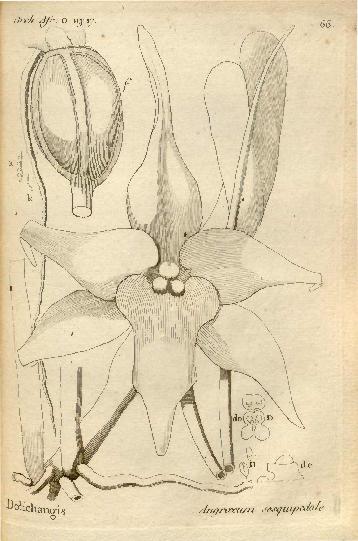
 Original drawing included in first publication of A. sesquipedale by Thoars.
Original drawing included in first publication of A. sesquipedale by Thoars.Star-like waxy flowers are produced on 30 cm (11.8 in) inflorescences arising from the stem from June to September in the wild with most flowers wilting by August.[14] When cultivated in Europe however, the plant flowers between December and January.[20] This flowering habit is what lends the orchid several of its common names, such as "Comet orchid" due to the shape of its flower and "Christmas orchid" due to the timing of its flowering. Each flower opens up with a green color, but eventually turns white with tones of light green. The amount of green in each flower can vary from plant to plant.[21] It is claimed that the age-dependent color change is more pronounced in wild A. sesquipedale than in greenhouse-grown plants.[22] The sepals tend to stay green for a longer time than the petals. As the flower ages further it goes from white to yellowish and then from orange to brown as it begins to wilt. As the wilting progresses the dorsal sepal bends down and then the lateral sepals bend inward with the lip remaining fairly stationary. Finally the entire flower closes in on itself.[23] A distinctive feature of the flowers is that they have a long green spur attached. The spur of the flower is 27–43 cm (10.6–16.9 in) from its tip to the tip of the flower's lip.[24] The specific epithet sesquipedale is Latin for "one-and-a-half-feet-long," referring to the distance between the end of the spur and the very top of the dorsal sepal.[14][25] At the end of the spur is a small amount of nectar usually about 40–300 µl in volume. In general, longer spurs tend to have greater concentrations of nectar. This nectar fills the spur up to within 7 to 25 cm (2.8 to 9.8 in) from the bottom of the spur.[22] The nectar has been found to contain the sugars fructose, sucrose, glucose, and raffinose.[26] The flowers produce an extremely intense spicy scent that can easily fill a room; this fragrance is only present during the night and is reminiscent of lily and some nocturnally flowering Nicotiana species.[27] The scent has been found to be composed of approximately 39 different chemical constituents with its greatest concentration consisting of isovaleraldoxime, methyl benzoate, benzyl alcohol, isovaleronitrile, benzyl benzoate, phenylethyl alcohol, isovaleraldehyde, and phenylacetaldoxime.[28] Usually 1 to 5 flowers are produced at a time.[21][29][30]
Reproduction
Coevolution model
This orchid is well-known within the botany community, especially for its association with the naturalist Charles Darwin. After being sent several flowers of A. sesquipedale by James Bateman,[31] Darwin noted the defining characteristic of the species, its extremely long spur.[32] From his observations, Darwin surmised, in his 1862 publication On the various contrivances by which British and foreign orchids are fertilized by insects, and on the good effects of intercrossing, that there must be a pollinator moth with a proboscis long enough to reach the nectar at the end of the spur. He arrived at this conclusion after attempting in vain to remove the pollinia of the flower using needles and bristles. Only after placing a cylinder with a diameter of 1⁄10 of an inch down the full length of the spur was he able to detach the pollinia upon retracting it. The viscidium attached to the cylinder as he removed it. Darwin surmised that in the moth's attempt at getting the nectar at the end of the spur the moth would get the pollinarium attached to itself.[33] The next orchid it visited would then be pollinated in the same manner.[34] For some time after this prediction the notion of a pollinator with a 35 cm long proboscis was ridiculed and generally not believed to exist.[35][36] After Darwin's publication, George Campbell published a book in 1867 titled, The Reign of Law, in which he attempted to argue that the complexity of this species implied that it was created by a supernatural being.[37] However, in 1903, such a moth was discovered in Madagascar by Lionel Walter Rothschild and Karl Jordan.[38][39] This confirmed Darwin's prediction. The moth was named Xanthopan morganii praedicta. The subspecific epithet praedicta was given in honor of the fact that Darwin predicted its existence, though the subspecies was later determined to be invalid (it is identical to the mainland form of the species).
 Original drawing included in first publication of A. sesquipedale by Thoars.
Original drawing included in first publication of A. sesquipedale by Thoars.In 1873 W. A. Forbes wrote an article in the journal Nature asking readers if they knew of the moth predicted by Darwin.[40] A reply to the question was first made that same year by Hermann Müller. He announced that his brother Fritz Müller had discovered a moth with a proboscis of 30–33 cm (11.8–13.0 in) long, but it was discovered in Brazil and so was not a candidate for pollinating A. sesquipedale.[41][42] Although Darwin learned of Müller's finding he did not live to see the discovery of Xanthopan morgani.[43][44] Even after the 1903 discovery however, news of Xanthopan morganii praedicta didn't immediately disseminate. A second inquiry into the existence of the pollinator moth was made in the January 30, 1907 issue of the journal Nature by E. W. Swanton. Presumably still unaware of Rothschild and Jordan's discovery, Alfred Russel Wallace responded stating that he didn't know of a suitable pollinator in Madagascar, but that he had heard of one from East Africa with a long enough proboscis.[45]
Both Darwin and Alfred Russel Wallace had suggested that the evolutionary basis for how the odd relationship between the sphinx moth and A. sesquipedale evolved over time could be understood by considering one orchid with a long spur and another with a short spur.[34][46] If a moth goes to fertilize a flower with a short spur its proboscis would easily reach all the way to the bottom of the spur and it would get the nectar. However, since the proboscis of the moth is longer than the spur of the flower; the head of the moth would not touch the flower obtaining the pollinarium and so the flower would not be fertilized.[47] The orchid with the longer spur on the other hand would be able to be fertilized since the entire length of the proboscis fits within the spur and thus allowing the head of the moth to touch the flower and become connected to the pollinarium. As a result, over time plants with longer spurs would be more likely to reproduce and so become more prevalent in the population.[48] In this way A. sesquipedale has evolved to have a very long spur. The moth too would evolve to have a longer and longer proboscis in the following way. If a moth goes to fertilize an A. sesquipedale flower and the spur is longer than its proboscis then it will not be able to reach all of the nectar. As such, moths with too short of a proboscis would not be able to get as much food as those moths with a longer proboscis who could reach all of the nectar. Due to this arrangement moths with longer proboscis would become more physically fit to reproduce due to their ability to get more nectar and so such moths would become more prevalent in the population. This can result in a seasawing effect by which both organisms produce a mechanism that leads the other to increase the others spur and proboscis.[49] There are however certain properties that no doubt prevent this mechanism from continuing indefinitely. For example, the risk such a long proboscis poses to a moth could be a factor that would prevent the spur of A. sesquipedale from becoming indefinitely long. If moths with proboscises that were too long substantially risked their lives due to being easier prey, then such moths could only afford to evolve a proboscis to a certain length. This would in turn restrict the length of the orchid's spur, since moths would not want to visit flowers whose spurs were too long since they would not be able to reach the nectar.[50]
There was also another explanation for why the spur of A. sesquipedale grew so long proposed by Thomas Belt in his 1874 book The Naturalist in Nicaragua.[51] Belt suggested that the spur grew long in order to prevent other moths with shorter proboscises from drinking the nectar. Darwin took up this explanation briefly in a footnote of the second edition of his famous orchid book, explaining that although this explanation was no doubt true, it cannot account for the lengthening spur.[52]
 An illustration by Thomas William Wood, based on Alfred Russel Wallace's description, showing a moth pollinating A. sesquipedale. This was Wallace's conception of the event since it was drawn in 1867 before the moth was even discovered.[53]
An illustration by Thomas William Wood, based on Alfred Russel Wallace's description, showing a moth pollinating A. sesquipedale. This was Wallace's conception of the event since it was drawn in 1867 before the moth was even discovered.[53]The fertilization of A. sesquipedale has been observed to proceed as follows. The moth approaches the flower to ascertain by scent whether or not it is the correct orchid species. Then the moth backs up over a foot and unrolls its proboscis, then flies forward, inserting it into a cleft in the rostellum which leads to the spur while gripping the labellum. After the moth has finished drinking the nectar, which usually takes about 6 seconds,[22] it instinctively raises its head while removing its proboscis from the spur, and in doing so causes the viscidium to adhere to its proboscis usually about 4 to 9 mm (0.16 to 0.35 in) from its base.[22] Attached to the viscidium via the caudicle is the pollinia. Upon removing its proboscis from the flower, the pollinarium stalk will be straight and parallel with the moth's proboscis. Then after leaving the orchid the caudicle will eventually dry out, causing its angle relative to the moth's proboscis to change by 90° so that it is at the correct angle to attach to the stigma of the next orchid the moth visits. The moth then repeats this process at another A. sesquipedale orchid and simultaneously fertilizes it. Once the flower has been fertilized, it quickly stops producing its powerful scent.[23]
Pollinator shift model
An alternative path by which A. sesquipedale could have evolved that differed with Darwin and Wallace's explanation was proposed by Lutz Thilo Wasserthal in 1997.[22] According to Wasserthal, hawk moths could have evolved long proboscises as a predatory avoidance strategy from heteropodid spiders. Since such spiders have been known to jump at hovering moths in an attempt at eating them, hawk moths would be at risk when visiting flowers if such a spider was nearby. Based on this reasoning moths with longer tongues would be less at risk when pollinating flowers since they would be farther away and thus a more challenging target for jumping spiders. As a result nature would select for hawk moths with longer and longer proboscis. The flowers of A. sesquipedale on the other hand would be evolving longer spurs since flowers with longer spurs are more likely to become fertilized by long tongued moths. In other words, the flowers evolve long spurs to fit the pollinators and not the reverse. It has also been observed that the moths will swing side to side when feeding, presumably to evade jumping spiders.[54] Possible problems with this hypothesis is that active predation by spiders on hawk moths visiting flowers has not been observed.[55] It has also been suggested that flying predators such as bats and birds are the more likely predators to hawk moths. Whether or not the pollinator shift model or the coevolution model, or even a little bit of both are correct is currently the subject of debate.[56]
Related species
In 2008, Micheneau et al. performed a cladistic analysis of the Angraecoid species. The following is a portion of the resultant Bayesian tree they obtained, showing part of clade 2.[57]
0.58 1.0 1.0 0.92 1.0 0.97 1.0 A. didlen
A. aff. rutenbergianum
A. pseudodidien
A. obesum
A. liliodorum
A. cornigerum
A. borbonicum
A. cucullatum
A. sesquipedale
The cladogram indicates that A. cucullatum is the most closely related species to A. sesquipedale.
Cultivation
 A. sesquipedale in bud
A. sesquipedale in budAngraceum sesquipedale was first brought to the United Kingdom in 1855 to be grown outside of its natural environment by William Ellis. Subsequently, Ellis achieved the first flowering of the plant in cultivation in 1857.[35] Angraecum sesquipedale has been attributed as having a nicer appearance when grown in cultivation than when found in the wild, since wild specimens appear as a long stem surrounded by a few struggling leaves.[58] Additionally, A. sesquipedale is seldom grown in private collections, despite its enormous importance to Darwin's concept of co-evolution and subsequently the fields of botany and evolutionary biology.
 A. sesquipedale flower
A. sesquipedale flowerIt's often recommended that A. sesquipedale be grown under warm to intermediate conditions and given as much light as possible without burning the leaves.[59] The choice of growing the plant in intermediate or warm housing conditions can affect the timing of the flowering.[14][20] In order to stimulate heavy flower production its important that the light intensity be greatest between September and November.[14] The number of flower spikes present during flowering is dependent on the number of new leaf-pairs formed during the preceding spring and summer, since each newly formed leaf-pair produce one spike and rarely two.[14] Angraecum sesquipedale is commonly found to have a slow growth habit, but the orchid can be expected to produce flowers even before it has reached an adult size.[60] Angraecum sesquipedale is notorious for having sensitive roots. The roots of mature plants are best left undisturbed as much as possible and as a result its prudent to be especially careful during repotting. Young plants however are less susceptible to such root problems. Disturbing the roots can cause the plant to sulk for 2 to 4 years or even to cause it to die. When a mature plant is disturbed it frequently looses many of its lower leaves and reverts to only producing one or two flowers at a time.[14] To avoid these problems its commonly advised that the orchid be planted in a coarse medium such as fir bark, crock, or charcoal to minimize disturbances to the roots.[14] Also planting it in a basket or large pot is best since this allows the orchid to grow for many years before having to have its roots disturbed.[3]
Hybrids
The first Angraecum hybrid was created by John Seden, an employee of Veitch Nurseries, and exhibited for the first time on January 10, 1899.[61] It was named A. Veitchii, but it also commonly goes by the name King of the Angraceum hybrids. The cross was between A. sesquipedale and A. eburneum.[61][62][63] The flowers somewhat resemble those of A. leonis. Another common cross involving A. sesquipedale is A. Crestwood, which is a cross between A. Veitchii and A. sesquipedale.
Interspecific hybrids
- Angraecum Appalachian Star (Angraecum sesquipedale x Angraecum praestans)[64]
- Angraecum Crestwood (Angraecum Veitchii x Angraecum sesquipedale)[65]
- Angraecum Dianne's Darling (Angraecum sesquipedale x Angraecum Alabaster)[64]
- Angraecum Lemförde White Beauty (Angraecum magdalenae x Angraecum sesquipedale)[66]
- Angraecum Malagasy (Angraecum sesquipedale x Angraecum sororium)[65]
- Angraecum Memoria Mark Aldridge (Angraecum sesquipedale x Angraecum eburneum subsp. superbum)[65]
- Angraecum North Star (Angraecum sesquipedale x Angraecum leonis)[65]
- Angraecum Ol Tukai (Angraecum comorense x Angraecum sesquipedale)[65]
- Angraecum Orchidglade (Angraecum sesquipedale x Angraecum giryamae)[67]
- Angraecum Rose Ann Carroll (Angraecum eichlerianum x Angraecum sesquipedale)[67]
- Angraecum Sesquibert (Angraecum sesquipedale x Angraecum humbertii)[65]
- Angraecum Sesquivig (Angraecum viguieri x Angraecum sesquipedale)[65]
- Angraecum Star Bright (Angraecum sesquipedale x Angraecum didieri)[65]
- Angraecum Veitchii (Angraecum eburneum x Angraecum sesquipedale)[62]
Intergeneric hybrids
- Angranthes Paille en Queue (Angraecum sesquipedale x Aeranthes arachnites)[65]
- Angranthes Sesquimosa (Aeranthes ramosa x Angraecum sesquipedale)[65]
- Chouara Kaohsiung Dream (Angraecum sesquipedale x Doritaenopsis Ruey Lih Beauty)[65]
- Eurygraecum Lydia (Angraecum sesquipedale x Eurychone rothschildiana)[66]
- Vandaecum Enzomondo Amore (Vanda Rothschildiana x Angraecum sesquipedale)[65]
Notes
- ^ The name Sesquipedale is pronounced /ˌsɛskwɨpɨˈdeɪliː/
- ^ Petit-Thouars 1822, tab. 66
- ^ a b c d Stewart & Campbell 1970, p. 50
- ^ Ellis 1858, pp. 40–41
- ^ Bosser & Morat 1972, pp. 76–78
- ^ Garay 1973, p. 514
- ^ Stewart, Hermans & Campbell 2006, p. 42
- ^ Jones 1967, p. 151
- ^ The Arends source quotes 2n=38 in contradiction with the Jones source.
- ^ Edwards, & Lindley 1824, 817
- ^ Schimper 1903, p. 131
- ^ Kuntze 1891, p. 651
- ^ Rolfe 1904, pp. 46–47
- ^ a b c d e f g h Hillerman 1975, pp. 104–105
- ^ Kluge, Vinson & Ziegler 1997, pp. 46–47
- ^ a b Fowlie 1969, p. 209
- ^ Ellis 1858, pp. 177–178
- ^ Mayr 2005, p. 13
- ^ There is a contradiction in the literature as the source from F.E. Hillerman states that the plant is exclusively only found along a narrow strip on the east coast of the island that gets between 80-100 inches of rainfall a year.
- ^ a b If grown in warm house conditions the plant will flower around Christmas day, while if grown in intermediate house conditions it will flower around mid-January or later.
- ^ a b Ellis 1858, pp. 41–42
- ^ a b c d e Wasserthal 1997, pp. 343–359
- ^ a b Arditti 1979, p. 570
- ^ In a paper published by Wasserthal in 1997 he quoted a spur length of 33.3±4.6 cm (N=15). The spur was measured from the rostellum at the spur entrance to the end of the spur lumen.
- ^ According to F.E. Hillerman some mature species can in fact reach these proportions.
- ^ Jeffrey, Arditti & Koopowitz 1970, p. 192
- ^ Kaiser 1993, p. 246
- ^ Kaiser 1993, p. 247
- ^ Warner & Williams 1897, plate 518
- ^ The Warner source quotes 1–4 flowers whereas the Ellis source quotes 3–5 flowers.
- ^ The orchid flowers were sent unidentified and were later identified by James Bateman's son Robert Bateman in a letter to Darwin. Letter 3357 — Bateman, Robert to Darwin, C. R., [1862], Darwin Correspondence Project.
- ^ Darwin 1997, Letter 3421 Darwin to Hooker 30 Jan. 1862
- ^ Lecoufle 1982, p. 99
- ^ a b Darwin 1862, pp. 197–203
- ^ a b van der Cingel 2001, p. 155
- ^ Rodríguez-Gironés & Santamaría 2007, p. 455
- ^ Campbell 2005, pp. 44–52
- ^ Rothschild & Jordan 1903, plate XXII
- ^ The J.A. Fowlie source claims that the moth was first discovered by a missionary in the 1880's and subsequently the moth found its way into Humbold's collection in 1882. All other sources however accredit Rothschild and Jordan as the discoverers.
- ^ Forbes 1873, p. 121
- ^ Müller 1873, p. 223
- ^ Müller 1878, p. 221
- ^ Thompson 1994, p. 26
- ^ Beatty 2006, pp. 632–633
- ^ Just 1959, p. 5
- ^ Wallace 1867, pp. 474–477
- ^ Cox & Grubb 1991, p. 220
- ^ Lecoufle 1982, pp. 99–102
- ^ Wallace 2009, pp. 146–149
- ^ Kritsky 1991, pp. 206–210
- ^ Belt 1985, p. 146
- ^ Rodríguez-Gironés & Llandres 2008, p. 1
- ^ Darwin 2005, Letter 5637 Wallace to Darwin 1 Oct. 1867
- ^ Kitching 2002, p. 473
- ^ Anders Nilsson 1998, pp. 250–269
- ^ Johnson & Anderson 2010, pp. 34–38
- ^ Micheneau et al. 2008, p. 914
- ^ Bean 1891, p. 437
- ^ F.E. Hillerman stated in an Orchid Digest article that 3,000-4,000 footcandles isn't too much.
- ^ F.E. Hillerman stated in a Orchid Digest article that the plant can begin flowering after only four years after having first sowed the seeds, which would correspond with a height of 6 inches.
- ^ a b Seden et al. 1899, p. 31
- ^ a b Pridgeon 2006, p. 31
- ^ There is some ambiguity in the literature as to the parentage of this hybrid. Some sources claim that the seed parent is A. eburneum subsp. superbum, while the majority of the sources like the Royal Horticultural Society claims it's simply A. eburneum and that (A. eburneum subsp. superbum x A. sesquipedale) should be called Angcm. Memoria Mark Aldridge.
- ^ a b Frowine 2005, pp. 100–101
- ^ a b c d e f g h i j k l The Royal Horticultural Society,The International Orchid Register.
- ^ a b LaCroix & LaCroix 2003, p. 78
- ^ a b LaCroix & LaCroix 2003, p. 83
References
- Anders Nilsson, L. (1 July 1998). "Deep flowers for long tongues". Trends in Ecology and Evolution 13 (7): 259–260. doi:10.1016/S0169-5347(98)01359-7. PMID 21238293.
- Arditti, Joseph (1979). Woolhouse, H. W.. ed. "Aspects of the Physiology of Orchids". Advances in Botanical Research (Academic Press Inc.) 7: 422–638. ISBN 978-0-12-005907-2. http://books.google.com/?id=IIHO10OyyZ0C&pg=PA421&dq=Angraecum+%27Veitchii%27#v=onepage&q=arditti&f=false.
- Arends, J. C.; Van der Laan, F. M. (1983). "Cytotaxonomy of the monopodial orchids of the African and Malagasy regions". Genetica (Springer Netherlands) 62 (2): 81–94. doi:10.1007/BF00116630. ISSN 0016-6707.
- Bean, William Jackson (9 May 1891). "Garden Flora: Angræcums". The Garden an Illustrated Weekly Journal of Horticulture in all its Branches (London) 39: 436–437.
- Beatty, John (December 2006). "Chance Variation: Darwin on Orchids". Philosophy of Science (The University of Chicago Press) 73 (5): 629–641. doi:10.1086/518332.
- Belt, Thomas (1 September 1985) [First published 1874]. The Naturalist in Nicaragua. University Of Chicago Press. ISBN 978-0-226-04219-0.
- Bosser, Jean; Morat, Philippe (1972). "Contribution à l'étude des Orchidaceae de Madagascar : 18". Adansonia.Série 2 12 (1): 73–78.
- Campbell, George; George Douglas Campbell, Duke of Argyll (22 December 2005) [First published 1867]. The Reign of Law. Scholarly Publishing Office. ISBN 978-1-4255-5627-3.
- Cox, Paul Alan; Grubb, Peter J. (29 August 1991). "Abiotic Pollination: An Evolutionary Escape for Animal-Pollinated Angiosperms [and Discussion]". Philosophical Transactions: Biological Sciences (The Royal Society) 333 (1267): 217–224. doi:10.1098/rstb.1991.0070.
- Topham, Jonathan R.; Frederick Burkhardt, Joy Harvey, Duncan M. Porter (13 June 1997). The Correspondence of Charles Darwin. 10. Cambridge University Press. ISBN 978-0-521-59032-7.
- Burkhardt, Frederick; Frederick Burkhardt, Sydney Smith (2006-04-03). The Correspondence of Charles Darwin. 15. Cambridge University Press. ISBN 978-0-521-85931-8.
- Darwin, Charles (25 June 2004) [First published 1862]. On the various contrivances by which British and foreign orchids are fertilised by insects, and on the good effects of intercrossing. Kessinger Publishing. ISBN 978-1-4179-2156-0. http://darwin-online.org.uk/content/frameset?viewtype=text&itemID=F800&pageseq=1.
- Edwards, Sydenham; Lindley, John (1824). The Botanical Register: Consisting of Coloured Figures of Exotic Plants, Cultivated in British Gardens; with their History and Mode of Treatment. 10. London. http://www.biodiversitylibrary.org/item/9046#1.
- Ellis, William (1858). Three visits to Madagascar during the years 1853-1854-1856: including a journey to the capital : with notices of the natural history of the country and of the present civilization of the people. John Murray. http://books.google.com/?id=s_AMAAAAIAAJ&printsec=frontcover&dq=Three+visits+to+Madagascar+during+the+years+1853-1854-1856:+including+a+journey+to+the+capital+:+with+notices+of+the+natural+history+of+the+country+and+of+the+present+civilization+of+the+people.#v=onepage&q&f=false.
- Forbes, William Alexander (12 June 1873). "Fertilisation of Orchids". Nature 8 (189): 121. doi:10.1038/008121b0.
- Fowlie, Jack A. (1969). "The Curious Tryst of the Star Orchid of Madagascar (Angraecum sesquipedale)". Orchid Digest 33 (7): 209–210.
- Frowine, Steven A. (15 October 2005). Fragrant Orchids: A Guide to Selecting, Growing, and Enjoying. Timber Press. ISBN 978-0-88192-739-9.
- Garay, Leslie A. (1973). "Systematics of the Genus Angraecum (Orchidaceae)". Kew Bulletin (Springer) 28 (3): 495–516. doi:10.2307/4108894. JSTOR 4108894.
- Hillerman, Fred E. (1975). "Angraecum sesquipedale: the Comet Orchid". Orchid Digest 39 (3): 104–105.
- Jeffrey, David C.; Arditti, Joseph; Koopowitz, Harold (January 1970). "Sugar Content in Floral and Extrafloral Exudates of Orchids: Pollination, Myrmecology and Chemotaxonomy Implication". New Phytologist (Blackwell Publishing) 69 (1): 187–195. doi:10.1111/j.1469-8137.1970.tb04062.x.
- Johnson, Steven D.; Anderson, Bruce (March 2010). "Coevolution Between Food-Rewarding Flowers and Their Pollinators". Evolution: Education and Outreach (Springer) 3 (1): 32–39. doi:10.1007/s12052-009-0192-6.
- Jones, Keith (1967). "The Chromosomes of Orchids: II: Vandeae Lindl". Kew Bulletin (Springer) 21 (1): 151–156. doi:10.2307/4108458. JSTOR 4108458.
- Just, Theodor Karl (November 1959). "Darwin, an Orchid, and a Moth". Chicago Natural History Museum Bulletin 30 (11).
- Sugisawa, Hiroshi (5 May 1993). "18". Bioactive Volatile Compounds from Plants. An American Chemical Society Publication. pp. 240–268. doi:10.1021/bk-1993-0525.ch018. ISBN 978-0-8412-2639-5.
- Kitching, Ian J. (29 July 2002). "The phylogenetic relationships of Morgan's Sphinx, Xanthopan morganii (Walker), the tribe Acherontiini, and allied long-tongued hawkmoths (Lepidoptera: Sphingidae, Sphinginae)". Zoological Journal of the Linnean Society (The Linnean Society of London) 135 (4): 471–527. doi:10.1046/j.1096-3642.2002.00021.x.
- Kluge, Manfred; Vinson, B.; Ziegler, Hubert (1997). "Ecophysiological Studies on Orchids of Madagascar: Incidence and Plasticity of Crassulacean Acid Metabolism in Species of the Genus Angraecum Bory". Plant Ecology (Springer) 135 (1): 43–57.
- Kritsky, Gene (4 November 1991). "Darwin's Madagascan Hawk Moth Prediction". American Entomologist (Entomological Society of America) 37 (4): 206–210.
- Kuntze, Otto (1891) (in German). Revisio Generum Plantarum. 2. http://www.biodiversitylibrary.org/item/7554#1.
- I. F. La Croix; Eric La Croix (1 January 2003). African Orchids in the Wild and in Cultivation. Timber Press. ISBN 978-0-88192-405-3.
- Lecoufle, Marcel (1982). "The Angraecum sesquipedale and Pollinization". Orchid Digest 46 (3): 99–102.
- Mayr, Hubert (2005). "Beautiful Angraecum and Aerangis: An Expedition to Madagascar". Orchid Digest 69 (1): 8–15.
- Micheneau, Claire; Carlsward, Barbara S.; Fay, Michael F.; Bytebier, Benny; Pailler, Thierry; Chase, Mark W. (March 2008). "Phylogenetics and biogeography of Mascarene angraecoid orchids (Vandeae, Orchidaceae)". Molecular Phylogenetics and Evolution 46 (3): 908–922. doi:10.1016/j.ympev.2007.12.001. PMID 18272406.
- Müller, Hermann (17 July 1873). "Probosces capable of sucking the Nectar of Anagræcum sesquipedale". Nature 8 (194): 223. doi:10.1038/008223a0.
- Müller, Hermann (17 January 1878). "Macrosilia cluentius". Nature 17 (429): 221. doi:10.1038/017221b0.
- Petit-Thouars, Louis-Marie Aubert du (1822) (in French). Histoire particulière des plantes orchidées recueillies sur les trois iles australes d'Afrique, du France, de Bourbon et de Madagascar. L'auteur. http://books.google.com/?id=E1IaAAAAYAAJ&printsec=frontcover&dq=Histoire+Particuli%C3%A8re+des+Plantes+Orchid%C3%A9es+Recueillies+sur+les+trois+Isles+Australs+d%27Afrique.#v=onepage&q&f=false.
- Pridgeon, Alec M.; Alasdair Morrison (1 April 2006). The Illustrated Encyclopedia of Orchids. Timber Press. ISBN 978-0-88192-801-3.
- Rodríguez-Gironés, Miguel A.; Llandres, Ana L.; Rogers, Naomi (August 2008). Rogers, Naomi. ed. "Resource Competition Triggers the Co-Evolution of Long Tongues and Deep Corolla Tubes". PLoS ONE 3 (8): 1–8. doi:10.1371/journal.pone.0002992.
- Rodríguez-Gironés, Miguel A.; Santamaría, Luis (September 2007). "Resource Competition, Character Displacement, and the Evolution of Deep Corolla Tubes". The American Naturalist (The University of Chicago Press) 170 (3): 455–464. doi:10.1086/520121. ISSN 00030147. PMID 17879195.
- Rolfe, Robert Allen (February 1904). "The Genus Mystacidium". The Orchid Review 12 (134): 46–47.
- Rothschild, Lionel Walter; Jordan, Karl (1903). A revision of the lepidopterous family Sphingidae. 9. Hazell, Watson & Viney, Ld..
- Schimper, Andreas Franz Wilhelm (1903). Groom, Percy; Balfour, Isaac Bayley. eds. Plant-geography upon a physiological basis. Clarendon Press. http://books.google.com/?id=I_MKAAAAIAAJ&printsec=frontcover&dq=Plant-geography+upon+a+physiological+basis#v=onepage&q&f=false.
- Seden, John; Veitch, James H.; Lindley, John; Paxton, Joseph (14 January 1899). "Societies: Orchid Committee". The Gardener's Chronicle, third series 25.
- Stewart, Joyce; Campbell, Bob (1970). Orchids of Tropical Africa. A.S. Barnes & Co.. ISBN 978-0-498-07555-1.
- Joyce Stewart, Joyce; Johan Hermans, Bob Ian Martin Campbell (1 August 2006). Angraecoid Orchids: Species from the African Region. Timber Press. ISBN 978-0-88192-788-7.
- Thompson, John N. (15 November 1994). The Coevolutionary Process. University of Chicago Press. ISBN 978-0-226-79760-1.
- Wallace, Alfred Russel (October 1867). "Creation by Law". Quarterly Journal of Science (John Churchill and Sons) 4: 471–488. http://books.google.com/?id=7a8VAAAAYAAJ&printsec=frontcover&dq=Quarterly+Journal+of+Science#v=onepage&q&f=false.
- Wallace, Alfred Russel (16 August 2009) [First published 1891]. Natural Selection and Tropical Nature. General Books LLC. ISBN 978-0-217-73022-8.
- Warner, Robert; Williams, Henry (1897). Orchid album: comprising coloured figures and descriptions of new, rare and beautiful orchidaceous plants. 11. B. S. Williams & Son.
- Wasserthal, L. T. (1997). "The Pollinators of the Malagsy Star Orchids Angraecum sesquipedale, A. sororium, and A. compactum and the Evolution of Extremely Long Spurs by Pollinator Shift". Botanica Acta (Georg Thieme Verlag) 110 (5): 343–359.
- Whittall, Justen B.; Hodges, Scott A. (7 June 2007). "Pollinator shifts drive increasingly long nectar spurs in columbine flowers". Nature 447 (7145): 706–709. doi:10.1038/nature05857. PMID 17554306.
- van der Cingel, Nelis A. (1 June 2001). An Atlas of Orchid Pollination. Taylor & Francis. ISBN 978-90-5410-486-5.
External links
External identifiers for Angraecum sesquipedale NCBI 78697 Also found in: Wikispecies Categories:- Angraecum
- Orchid species
- Plants and pollinators
- House plants
- Plants described in 1822
- Epiphytes






Wikimedia Foundation. 2010.