- Ammonite
-
For other uses, see Ammonite (disambiguation).
Ammonites
Temporal range: 400–65.5 Ma Devonian - Cretaceous
Artist's reconstruction of Asteroceras Scientific classification 
Kingdom: Animalia Phylum: Mollusca Class: Cephalopoda Subclass: †Ammonoidea
Zittel, 1884Orders and Suborders See text
Ammonite, as a zoological or paleontological term, refers to any member of the Ammonoidea an extinct subclass within the Molluscan class Cephalopoda which are more closely related to living coleoids (i.e. octopuses, squid, and cuttlefish) than they are to shelled nautiloids such as living species of Nautilus .
Ammonites are excellent index fossils, and it is often possible to link the rock layer in which they are found to specific geological time periods. Their fossil shells usually take the form of planispirals, although there were some helically-spiraled and non-spiraled forms (known as heteromorphs).
The name ammonite, from which the scientific term is derived, was inspired by the spiral shape of their fossilized shells, which somewhat resemble tightly coiled rams' horns. Pliny the Elder (d. 79 AD. near Pompeii) called fossils of these animals ammonis cornua ("horns of Ammon") because the Egyptian god Ammon (Amun) was typically depicted wearing ram's horns.[1] Often the name of an ammonite genus ends in -ceras, which is Greek (κέρας) for "horn".
Contents
Diagnostic characters
Ammonites (subclass Ammonoidea) can be distinguishes by their septa, the dividing walls that separate the chambers in the phragmocone, by the nature of their sutures where the septa joint the outer shell wall, and in general by their siphuncles.
Septa
Ammonoid septa characteristically have bulges and indentations and are to varying degrees convex from the front, distinguishing them from nautiloid septa which are typically simple concave dish-shaped structures. The topology of the septa, especially around the rim, results in the various suture patterns found.
Suture patterns
Three major types of suture patterns are found in the Ammonoidea:
- Goniatitic - numerous undivided lobes and saddles; typically 8 lobes around the conch. This pattern is characteristic of the Paleozoic ammonoids.
- Ceratitic - lobes have subdivided tips, giving them a saw-toothed appearance, and rounded undivided saddles. This suture pattern is characteristic of Triassic ammonoids and appears again in the Cretaceous "pseudoceratites".
- Ammonitic - lobes and saddles are much subdivided (fluted); subdivisions are usually rounded instead of saw-toothed. Ammonoids of this type are the most important species from a biostratigraphical point of view. This suture type is characteristic of Jurassic and Cretaceous ammonoids but extends back all the way to the Permian.
Siphuncle
The siphuncle in most ammonoids by far is a narrow tubular structure that runs along the outer rim, known as the venter, connecting the chambers of the phragmocone to the body or living chamber. This distinguishes them from living nautiloides (Nautilus and Allonautilus) and typical Nautilida. However the very earliest nautiloids from the Late Cambrian and Ordovician typically had ventral siphuncles, although often proportionally larger than those in ammonites and more internally structured.
Classification of the Ammonoidea
See also: List of ammonitesOriginating from within the bactritoid nautiloids, the ammonoid cephalopods first appeared in the Devonian (circa 400 million years ago) and became extinct at the close of the Cretaceous (65.5 Ma) along with the dinosaurs. The classification of ammonoids is based in part on the ornamentation and structure of the septa comprising their shells' gas chambers; by these and other characteristics we can divide subclass Ammonoidea into three orders and eight known suborders. While nearly all nautiloids show gently curving sutures, the ammonoid suture line (the intersection of the septum with the outer shell) is variably folded, forming saddles (or peaks) and lobes (or valleys).
Orders and suborders
 An ammonitic ammonoid with the body chamber missing, showing the septal surface (especially at right) with its undulating lobes and saddles.
An ammonitic ammonoid with the body chamber missing, showing the septal surface (especially at right) with its undulating lobes and saddles.
 Iridescent ancient ammonite fossil on display at the American Museum of Natural History, New York City, around 2.5 feet in diameter.
Iridescent ancient ammonite fossil on display at the American Museum of Natural History, New York City, around 2.5 feet in diameter.The Ammonoidea can be divided into eight orders, listed here starting with the most primitive and going to the more derived.
- Anarcestida, Devonian
- Clymeniida, Upper Devonian
- Goniatitida, Middle Devonian - Upper Permian
- Prolecanitida, Upper Devonian - Upper Triassic
- Ceratitida, Permian - Triassic
- Phylloceratida, Triassic - Cretaceous
- Lytoceratida, Jurassic - Cretaceous
- Ammonitida, Lower Jurassic - Upper Cretaceous
In some classifications these are left as suborders, included in only three orders: Goniatitida, Ceratitida, and Ammonitida.
Taxonomy of the Treatise
The Treatise on Invertebrate Paleontology (Part L, 1957) divides the Ammonoidea, regarded simply as an order, into eight suborders, the Anarcestina, Clymeniina, Goniatitina, and Prolecanitina from the Paleozoic; the Ceratitina from the Triassic; and the Ammonitina, Lytoceratina, and Phylloceratina from the Jurassic and Cretaceous. In subsequent taxonomies these are sometimes regarded as orders within the subclass Ammonoidea.
Life
 Jeletzkytes, a Cretaceous ammonite from the USA
Jeletzkytes, a Cretaceous ammonite from the USA Asteroceras, a Jurassic ammonite from England
Asteroceras, a Jurassic ammonite from EnglandBecause ammonites and their close relatives are extinct, little is known about their way of life. Their soft body parts are very rarely preserved in any detail. Nonetheless, much has been worked out by examining ammonoid shells and by using models of these shells in water tanks.
Many ammonoids probably lived in the open water of ancient seas, rather than at the sea bottom. This is suggested by the fact that their fossils are often found in rocks that were laid down under conditions where no bottom-dwelling life is found. Many of them (such as Oxynoticeras) are thought to have been good swimmers with flattened, discus-shaped, streamlined shells, although some ammonoids were less effective swimmers and were likely to have been slow-swimming bottom-dwellers. Synchrotron analysis of an aptychophoran ammonite revealed remains of isopod and mollusc larva in its buccal cavity, indicating that at least this kind of ammonite fed on plankton.[2] Fossilized ammonoids have been found showing tooth marks from such attacks. They may have avoided predation by squirting ink, much like modern cephalopods; ink is occasionally preserved in fossil specimens.[3]
The soft body of the creature occupied the largest segments of the shell at the end of the coil. The smaller earlier segments were walled off and the animal could maintain its buoyancy by filling them with gas. Thus the smaller sections of the coil would have floated above the larger sections.[4]
Shell anatomy and diversity
Basic shell anatomy
 A variety of ammonite forms, from Ernst Haeckel's 1904 Kunstformen der Natur (Art Forms of Nature).
A variety of ammonite forms, from Ernst Haeckel's 1904 Kunstformen der Natur (Art Forms of Nature). Baculites ammonite from the Late Cretaceous of Wyoming. The original aragonite of the outer conch and inner septa has dissolved away, leaving this articulated internal mold.
Baculites ammonite from the Late Cretaceous of Wyoming. The original aragonite of the outer conch and inner septa has dissolved away, leaving this articulated internal mold.The chambered part of the ammonite shell is called a phragmocone. The phragmocone contains a series of progressively larger chambers, called camerae (sing. camera) that are divided by thin walls called septa (sing. septum). Only the last and largest chamber, the body chamber, was occupied by the living animal at any given moment. As it grew, it added newer and larger chambers to the open end of the coil. A thin living tube called a siphuncle passed through the septa, extending from the ammonite's body into the empty shell chambers. Through a hyperosmotic active transport process, the ammonite emptied water out of these shell chambers. This enabled it to control the buoyancy of the shell and thereby rise or descend in the water column.
A primary difference between ammonites and nautiloids is that the siphuncle of ammonites (excepting Clymeniina) runs along the ventral periphery of the septa and camerae (i.e., the inner surface of the outer axis of the shell), while the siphuncle of nautiloids runs more or less through the center of the septa and camerae.
Sexual dimorphism
 Discoscaphites iris, Owl Creek Formation (Upper Cretaceous), Ripley, Mississippi.
Discoscaphites iris, Owl Creek Formation (Upper Cretaceous), Ripley, Mississippi.One feature found in shells of the modern Nautilus is the variation in the shape and size of the shell according to the sex of the animal, the shell of the male being slightly smaller and wider than that of the female. This sexual dimorphism is thought to be an explanation for the variation in size of certain ammonite shells of the same species, the larger shell (called a macroconch) being female, and the smaller shell (called a microconch) being male. This is thought to be because the female required a larger body size for egg production. A good example of this sexual variation is found in Bifericeras from the early part of the Jurassic period of Europe.
It is only in relatively recent years that the sexual variation in the shells of ammonites has been recognized. The macroconch and microconch of one species were often previously mistaken for two closely related but different species occurring in the same rocks. However, these "pairs" were so consistently found together that it became apparent that they were in fact sexual forms of the same species.
Variations in shape
The majority of ammonite species feature a shell that is a planispiral flat coil, but other species feature a shell that is nearly straight (as in baculites). Still other species' shells are coiled helically, superficially like that of a large gastropod (as in Turrilites and Bostrychoceras). Some species' shells are even initially uncoiled, then partially coiled, and finally straight at maturity (as in Australiceras). These partially uncoiled and totally uncoiled forms began to diversify mainly during the early part of the Cretaceous and are known as heteromorphs.
Perhaps the most extreme and bizarre looking example of a heteromorph is Nipponites, which appears to be a tangle of irregular whorls lacking any obvious symmetrical coiling. However, upon closer inspection the shell proves to be a three-dimensional network of connected "U" shapes. Nipponites occurs in rocks of the upper part of the Cretaceous in Japan and the USA.
Ammonites vary greatly in the ornamentation (surface relief) of their shells. Some may be smooth and relatively featureless, except for growth lines, and resemble that of the modern Nautilus. In others various patterns of spiral ridges and ribs or even spines are shown. This type of ornamentation of the shell is especially evident in the later ammonites of the Cretaceous.
Aptychus
Main article: Aptychus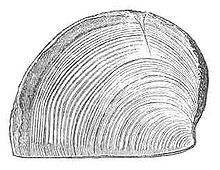 A drawing of an aptychus named "Trigonellites latus" from the Kimmeridge Clay Formation in England
A drawing of an aptychus named "Trigonellites latus" from the Kimmeridge Clay Formation in EnglandSome ammonites have been found in association with a single horny plate or a pair of calcitic plates. In the past it was assumed that these plates served to close the opening of the shell in much the same way as an operculum, however more recently it has been postulated that they were instead a jaw apparatus.[5][6][7][8]
The plates are collectively termed the aptychus or aptychi in the case of a pair of plates, and anaptychus in the case of a single plate. The paired aptychi were symmetrical to one another and equal in size and appearance.
Anaptychi are relatively rare as fossils. They are found representing ammonites from the Devonian period through those of the Cretaceous period.
Calcified aptychi only occur in ammonites from the Mesozoic era. They are almost always found detached from the shell, and are only very rarely preserved in place. Still, sufficient numbers have been found closing the apertures of fossil ammonite shells as to leave no doubt as to their identity as part of the anatomy of an ammonite.
Large numbers of detached aptychi occur in certain beds of rock (such as those from the Mesozoic in the Alps). These rocks are usually accumulated at great depths. The modern Nautilus lacks any calcitic plate for closing its shell, and only one extinct nautiloid genus is known to have borne anything similar. Nautilus does, however, have a leathery head shield (the hood) which it uses to cover the opening when it retreats inside.
There are many forms of aptychus, varying in shape and the sculpture of the inner and outer surfaces, but because they are so rarely found in position within the shell of the ammonite it is often unclear to which species of ammonite one kind of aptychus belongs. A number of aptychi have been given their own genus and even species names independent of their unknown owners' genus and species, pending future discovery of verified occurrences within ammonite shells.
Soft-part anatomy
Although ammonites do occur in exceptional lagerstatten such as the Solnhofen limestone, their soft part record is surprisingly bleak. Beyond a tentative ink sac and possible digestive organs, no soft parts are known at all.[9] They likely bore a radula and beak, a marginal siphuncle, and ten arms.[10] They operated by direct development with sexual reproduction, were carnivorous and had a crop for food storage. It is unlikely that any ammonoids dwelt in fresh or brackish water.[11]
Size
 2-metre (6.5-foot) Parapuzosia seppenradensis cast in Germany
2-metre (6.5-foot) Parapuzosia seppenradensis cast in GermanyFew of the ammonites occurring in the lower and middle part of the Jurassic period reach a size exceeding 23 centimetres (9 inches) in diameter. Much larger forms are found in the later rocks of the upper part of the Jurassic and the lower part of the Cretaceous, such as Titanites from the Portland Stone of Jurassic of southern England, which is often 53 centimetres (2 feet) in diameter, and Parapuzosia seppenradensis of the Cretaceous period of Germany, which is one of the largest known ammonites, sometimes reaching 2 metres (6.5 feet) in diameter. The largest documented North American ammonite is Parapuzosia bradyi from the Cretaceous with specimens measuring 137 centimetres (4.5 feet) in diameter, although a new 2.3-metre (7.5-foot) British Columbian specimen, if authentic, would appear to trump even the European champion.[12]
Distribution
 A specimen of Hoploscaphites from the Pierre Shale of South Dakota. Much of the original shell, including the nacre, has survived.
A specimen of Hoploscaphites from the Pierre Shale of South Dakota. Much of the original shell, including the nacre, has survived.Starting from the mid-Devonian, ammonoids were extremely abundant, especially as ammonites during the Mesozoic era. Many genera evolved and ran their course quickly, becoming extinct in a few million years. Due to their rapid evolution and widespread distribution, ammonoids are used by geologists and paleontologists for biostratigraphy. They are excellent index fossils, and it is often possible to link the rock layer in which they are found to specific geological time periods.
Due to their free-swimming and/or free-floating habits, ammonites often happened to live directly above seafloor waters so poor in oxygen as to prevent the establishment of animal life on the seafloor. When upon death the ammonites fell to this seafloor and were gradually buried in accumulating sediment, bacterial decomposition of these corpses often tipped the delicate balance of local redox conditions sufficiently to lower the local solubility of minerals dissolved in the seawater, notably phosphates and carbonates. The resulting spontaneous concentric precipitation of minerals around a fossil is called a concretion and is responsible for the outstanding preservation of many ammonite fossils.
When ammonites are found in clays their original mother-of-pearl coating is often preserved. This type of preservation is found in ammonites such as Hoplites from the Cretaceous Gault clay of Folkestone in Kent, England.
The Cretaceous Pierre Shale formation of the United States and Canada is well known for the abundant ammonite fauna it yields, including Baculites, Placenticeras, Scaphites, Hoploscaphites, and Jeletzkytes, as well as many uncoiled forms. Many of these also have much or all of the original shell, as well as the complete body chamber, still intact. Many Pierre Shale ammonites, and indeed many ammonites throughout earth history, are found inside concretions.
 An iridescent ammonite from Madagascar.
An iridescent ammonite from Madagascar.Other fossils, such as many found in Madagascar and Alberta (Canada), display iridescence. These iridescent ammonites are often of gem quality (ammolite) when polished. In no case would this iridescence have been visible during the animal's life; additional shell layers covered it.
The majority of ammonoid specimens, especially those of the Paleozoic era, are preserved only as internal molds; that it to say, the outer shell (composed of aragonite)[13] has been lost during the fossilization process. It is only in these internal-mold specimens that the suture lines can be observed; in life the sutures would have been hidden by the outer shell.
The ammonoids as a group continued through several major extinction events, although it appears that often only a few species survived. Each time, however, this handful of species diversified into a multitude of forms. Ammonite fossils became less abundant during the latter part of the Mesozoic, with none surviving into the Cenozoic era. The last surviving lineages disappeared, along with the dinosaurs, 65 million years ago in the Cretaceous-Tertiary extinction event. The reason why no ammonites survived the extinction event at the end of the Cretaceous, whereas some nautiloid cousins survived, might be due to differences in ontogeny. If their extinction was due to a bolide strike, plankton around the globe could have been severely diminished, thereby dooming ammonite reproduction during its planktonic stage.
Extinction
 Ammonites in the permanent collection of The Children’s Museum of Indianapolis
Ammonites in the permanent collection of The Children’s Museum of IndianapolisThe extinction of the ammonites along with other marine animals and of course, non-avian dinosaurs, has been attributed to a bolide impact, marking the end of the Cretaceous Period. Regardless of what effect an impact may have had, many of these groups, including ammonoids, were already in serious decline. Previously ammonoid cephalopods barely survived several earlier major extinction events, often with only a few species surviving from which a multitude of forms diversified.
Eight or so species from only two families made it almost to the end of the Cretaceous, the order having gone through a more or less steady decline since the middle of the period.[citation needed] Six other families made it well into the upper Maastrichtian (uppermost stage of the Cretaceous) but were extinct well before the end. All told, 11 families entered the Maastrichtian, a decline from the 19 families known from the Cenomanian in the middle of the Cretaceous.
One reason given for their demise is that Cretaceous ammonites, being closely related to coleoids, had a similar reproductive strategy in which a huge number of eggs is laid in a single batch at the end of the life span. These, along with juvenile ammonites, are thought to have been part of the plankton at the surface of the ocean where they were killed off by the effects of an impact. Nautiloids, exemplified by modern nautiluses, are thought on the other hand to have had a reproductive strategy in which eggs were laid in smaller batches many times during the life span and on the sea floor well away from any direct effects of such a bolide strike, and thus survived.
Mythology
In medieval Europe, fossilised ammonites were thought to be petrified coiled snakes, and were called "snakestones" or, more commonly in medieval England, "serpentstones". They were considered to be evidence for the actions of saints such as Hilda of Whitby, a myth referenced in Sir Walter Scott's Marmion,[14] and Saint Patrick, and were held to have healing or oracular powers. Traders would occasionally carve the head of a snake onto the empty, wide end of the ammonite fossil, and then sell them to the public. In other cases the snake's head would be simply painted on.[15] Ammonites from the Gandaki river in Nepal are known as saligrams, and are believed by Hindus to be a concrete manifestation of God or Vishnu.[16]
Terminological note
The words ammonite and ammonoid are both used quite loosely in common parlance to refer to any member of subclass Ammonoidea. However, in stricter usage the term ammonite is reserved for members of suborder Ammonitina (or sometimes even order Ammonitida).
See also
- Ammolite - a gemstone formed from fossil ammonite shells.
- Belemnoidea
- Coleoidea
- Fossils and the geological timescale
- Nautiloidea
References
- ^ NH 37.40.167
- ^ The Role of Ammonites in the Mesozoic Marine Food Web Revealed by Jaw Preservation, Isabelle Kruta, Neil Landman, Isabelle Rouget, Fabrizio Cecca, Paul Tafforeau, SCIENCE, JANUARY 2011 VOL 331
- ^ Doguzhaeva, Larisa A.; Royal H. Mapes; Herbert Summesberger; and Harry Mutvei (2007). "The Preservation of Body Tissues, Shell, and Mandibles in the Ceratitid Ammonoid Austrotrachyceras (Late Triassic), Austria". In N.H.Landman et al.. Cephalopods Present and Past: New Insights and Fresh Perspectives. Dordrecht: Springer. pp. 221–238. doi:10.1007/978-1-4020-6806-5_11. ISBN 978-1-4020-6806-5.
- ^ "Introduction to Ammonoidea". The Geology of Portsdown Hill. http://www.bbm.me.uk/portsdown/PH_232_Ammonites.htm. Retrieved 2007-04-26.
- ^ Morton, N. 1981. Aptychi: the myth of the ammonite operculum. Lethaia 14(1): 57–61. doi:10.1111/j.1502-3931.1981.tb01074.x
- ^ Morton, N. & M. Nixon 1987. Size and function of ammonite aptychi in comparison with buccal masses of modem cephalopods. Lethaia 20(3): 231–238. doi:10.1111/j.1502-3931.1987.tb02043.x
- ^ Lehmann, U. & C. Kulicki 1990. Double function of aptychi (Ammonoidea) as jaw elements and opercula. Lethaia 23: 325–331. doi:10.1111/j.1502-3931.1990.tb01365.x
- ^ Seilacher, A. 1993. Ammonite aptychi; how to transform a jaw into an operculum? American Journal of Science 293: 20–32. doi:10.2475/ajs.293.A.20
- ^ Wippich, M. G. E.; Lehmann, J. (2004). "Allocrioceras from the Cenomanian (mid-Cretaceous) of the Lebanon and its bearing on the palaeobiological interpretation of heteromorphic ammonites". Palaeontology 47 (5): 1093–1107. doi:10.1111/j.0031-0239.2004.00408.x.
- ^ Landman, Neil H; Tanabe, Kazushige; Davis, Richard Arnold (1996). Ammonoid paleobiology. ISBN 9780306452222. http://books.google.ca/books?id=hKbkB4MzUIkC&pg=PA14.
- ^ Landman, Neil H; Tanabe, Kazushige; Davis, Richard Arnold (1996). Ammonoid paleobiology. ISBN 9780306452222. http://books.google.ca/books?id=hKbkB4MzUIkC&pg=PA17.
- ^ "Ammonites". Hanman's Fossil Replicas and Minerals. Archived from the original on 2003-02-10. http://web.archive.org/web/20030210130400/http://www.hanmansfossils.com/catalogs/fossils/ammonites/ammonites.shtml.
- ^ "Diagenesis of aragonite from Upper Cretaceous ammonites: a geochemical case-study". Sedimentology 28 (3): 423–438. 1981. Bibcode 1981Sedim..28..423B. doi:10.1111/j.1365-3091.1981.tb01691.x.
- ^ Lovett, Edward (September 1905). "The Whitby Snake-Ammonite Myth". Folk-Lore 16 (3): 333–4. http://en.wikisource.org/wiki/Folk-Lore._Volume_16/The_Whitby_Snake-Ammonite_Myth.
- ^ Cadbury, D. the Dinosaur Hunters. (Fourth Estate, 2000) (ISBN 1-85702-963-1), p.7
- ^ "Fossils: myths, mystery, and magic". The Independent (London). 2007-02-12. http://news.independent.co.uk/sci_tech/article2259490.ece. Retrieved 2010-04-23.
- Neal L. Larson, Steven D Jorgensen, Robert A Farrar and Peter L Larson. Ammonites and the other Cephalopods of the Pierre Seaway. Geoscience Press, 1997.
- Lehmann, Ulrich. The Ammonites: Their life and their world. Cambridge University Press, New York, 1981. Translated from German by Janine Lettau.
- Monks, Neale and Palmer, Phil. Ammonites. Natural History Museum, 2002.
- Walker, Cyril and Ward, David. Fossils. Dorling, Kindersley Limited, London, 2002.
- A Broad Brush History of the Cephalopoda by Dr. Neale Monks, from The Cephalopod Page.
- Ammonite maturity, pathology and old age By Dr. Neale Monks, from The Cephalopod Page. Essay about the life span of Ammonites.
- Cretaceous Fossils Taxonomic Index for Order Ammonoitida
- Deeply Buried Sediments Tell Story of Sudden Mass Extinction
External links
- Descriptions and pictures of ammonite fossils
- goniat.org, a palaezoic ammonoid database
- paleozoic.org: gallery of ammonite photographs
- photos of ammonites at Lyme Regis, UK
- TaxonConcept's data on cretaceous ammonites
- The ammonites of Peacehaven - photos of giant cretaceous ammonites in Southern England
- tonmo.com: The octopus news magazine online, Cephalopod fossil articles.
Notable fossil cephalopods (listed by first occurrence) Cenozoic The belemnite-cuttlefish-like Belosaepia
Evolutionary history of cephalopodsMesozoic - Advanced nautiloids: Nautilaceae • True ammonites
- Early coleoids: Proteroctopus, Styletoctopus, Keuppia, Palaeoctopus, Paleocirroteuthis
- Ceratitid ammonoids • late Nautilida
- Advanced belemnoids: Diplobelids: Diplobelus • Belemnites: Belemnites, Belemnopsis, Belemnotheutis
Palaeozoic - First appearance of long-lasting lineages: Goniatite and ceratite ammonoids • Probable coleoid ancestor Bactritida • Earliest belemnoids: Aulacocerids: Aulacoceras • Phragmoteuthids: Phragmoteuthis
- Early forms: the earliest cephalopod group Ellesmerocerida • Orthocones: Endocerida, Orthocerida, Actinocerida • Brevicones: Ascocerida, Oncocerida • Earliest coiled cephalopods: Tarphycerida • Spirulid?: Shimanskya
- Octopus: Pohlsepia
Cambrian - Earliest unambiguous cephalopods: Plectronocerida (Plectronoceras)
- Monoplacophoran-like ancestral forms: Knightoconus
- Possible, nude, "stem-group cephalopods": Nectocarididae
Categories:- Extinct cephalopods
- Ammonites
- Fossils












Wikimedia Foundation. 2010.