- Trypanosoma brucei
Taxobox
color = khaki
image_width = 240px
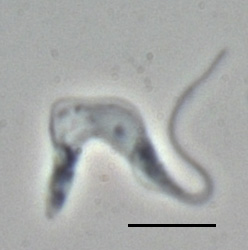
image_caption = "Trypanosoma brucei brucei" TREU667 (Bloodstream form, phase contrast picture. Black bar indicates 10 µm.)
name = "Trypanosoma brucei"
regnum =Protist a
phylum =Euglenozoa
classis = Kinetoplastea
ordo = Trypanosomatida
genus = "Trypanosoma "
species = "T. brucei"
binomial = "Trypanosoma brucei"
binomial_authority = Plimmer & Bradford, 1899
subdivision_ranks = Subspecies
subdivision = "T. b. brucei" "T. b. gambiense" "T. b. rhodesiense""Trypanosoma brucei" is a parasitic
protist species that causesAfrican trypanosomiasis (orsleeping sickness ) in humans andnagana in animals inAfrica . There are 3 sub-species of "T. brucei"; "T. b. brucei", "T. b. gambiense" and "T. b. rhodesiense".These obligate parasites have two hosts - an
insect vector andmammalian host. Because of the large difference between these hosts the trypanosome undergoes complex changes during its life cycle to facilitate its survival in the insect gut and the mammalian bloodstream. It also features a unique and notable variable surface glycoprotein (VSG) coat in order to avoid the host's immune system. There is an urgent need for the development of new drug therapies as current treatments can prove fatal to the patient as well as the trypanosomes.The trypanosome
cytoskeleton is the subject of considerable research. The cytoskeleton, as the structure behindmitosis , locomotion and surface binding, is vital for viability and so is a target of interest for drug development. Much research on "Trypanosoma brucei" is first done onCrithidia fasciculata a highly similar organism that is not dangerous to humans.The infection: Trypanosomiasis
The insect vector for "T. brucei" is the
tsetse fly . The parasite lives in the midgut of the fly (procyclic form), until it migrates to the salivary glands for injection to the mammalian host on binding. The parasite lives within the bloodstream (bloodstream form) where it can reinfect the fly vector after biting. Later during a "T. brucei" infection the parasite may migrate to other areas of the host. A "T. brucei" infection may be transferred human to human via bodily fluid exchange, primarily blood transfer.There are three different sub-species of "T. brucei", which cause different variants of trypanosomiasis.
*"T. brucei gambiense" - Causes slow onset chronic trypanosomiasis in humans. Most common in central and western Africa, where humans are thought to be the primary reservoir [Barrett et al. The trypanosomiases. The Lancet (2003) vol. 362 (9394) pp. 1469-1480 [http://www.ncbi.nlm.nih.gov/pubmed/14602444] ] .
*"T. brucei rhodesiense" - Causes fast onset acute trypanosomiasis in humans. Most common in southern and eastern Africa, where game animals and livestock are thought to be the primary reservoir [Barrett et al. The trypanosomiases. The Lancet (2003) vol. 362 (9394) pp. 1469-1480 [http://www.ncbi.nlm.nih.gov/pubmed/14602444] ] .
*"T. brucei brucei" - Causes animal African trypanosomiasis, along with several other species oftrypanosoma . "T. b. brucei" is not human infective due to its susceptibility to lysis by human apolipoprotein L1 [Vanhamme L, Paturiaux-Hanocq F, Poelvoorde P, et al. (2003). "Apolipoprotein L-I is the trypanosome lytic factor of human serum.". Nature 422 (6927): 83-7. [http://www.ncbi.nlm.nih.gov/pubmed/12621437] ] . However, as it shares many features with "T. b. gambiense" and "T. b. rhodesiense" (such as antigenic variation) it is used as a model for human infections in laboratory and animal studies.The cell structure
The structure of the cell is fairly typical of
eukaryote s, seeeukaryotic cell . All major organelles are seen, including the nucleus,mitochondria ,endoplasmic reticulum ,Golgi apparatus etc. Unusual features include the single large mitochondria with a condensedmitochondrial DNA structure, and its association with thebasal body of theflagellum , unusually thecytoskeleton organisation mechanism of the cell. The cell surface of the bloodstream form features a dense coat of variable surface glycoproteins (VSGs) which is replaced by an equally dense coat of procyclins when the parasite differentiates into the procylic in the tsetse fly midgut.Trypanosomatids show specific cellular forms:
*Amastigote - Basal body anterior of nucleus, with a short, essentially non-functional, flagellum.
*Promastigote - Basal body anterior of nucleus, with a long detached flagellum.
*Epimastigote - Basal body anterior of nucleus, with a long flagellum attached along the cell body.
*Trypomastigote - Basal body posterior of nucleus, with a long flagellum attached along the cell body.These names are derived from the Greek "mastig-" meaning
whip , referring to the trypanosome's whip-like flagellum."T. brucei" is found as a trypomastigote in the slender, stumpy, procyclic and metacyclic forms. The procylic form differentiates to the proliferitive epimastigote form in the salivary glands of the insect. Unlike Leishmania, the promastigote and the amastigote form does not form part of the "T.brucei" life cycle.
The genome
The
genome of "T. brucei" is made up of:
*11 largechromosome s of 1 to 6 megabase pairs.
*6 intermediate chromosomes of 300 to 600 kilobase pairs.
*Around 100 mini chromosomes of around 50 to 100 kilobase pairs. These may be present in multiple copies perhaploid genome.The large chromosomes contain mostgene s, while the small chromosomes tend to carry genes involved inantigen ic variation, including the VSG genes. The genome has been sequenced and is available online www.genedb.org.The mitochondrial genome is found condensed into the
kinetoplast , an unusual feature unique to thekinetoplastea class. It and thebasal body of theflagellum are strongly associated via a cytoskeletal structure.VSG surface coat
"Main section: The VSG coat"
The surface of the trypanosome is covered by a dense coat of Variable Surface Glycoprotein (VSG), which allows persistence of an infecting trypanosome population in the host. See below.
The cytoskeleton
The
cytoskeleton is predominantly made up ofmicrotubules , forming a subpellicular corset. The microtubules lie parallel to each other along the long axis of the cell, with the number of microtubules at any point roughly proportional to the circumference of the cell at that point. As the cell grows (including for mitosis) additional microtubules grow between the existing tubules, leading to semiconservative inheritance of the cytoskeleton. The microtubules are orientated + at the posterior and - at the anterior.Microfilament andintermediate filament s also play an important role in the cytoskeleton, but these are generally overlooked.Flagellar structure
The trypanosome flagellum has two main structures. It is made up of a typical flagellar axoneme which lies parallel to the paraflagellar rod, a lattice structure of proteins unique to the
kinetoplastida ,euglenoid s anddinoflagellate s.The microtubules of the flagellar
axoneme lie in the normal 9+2 arrangement, orientated with the + at the anterior end and the - in the basal body. The a cytoskeletal structure extends from the basal body to the kinetoplast. The flagellum is bound to the cytoskeleton of the main cell body by four specialised microtubules, which run parallel and in the same direction to the flagellar tubulin.The flagellar function is twofold - locomotion via oscilations along the attached flagellum and cell body, and attachment to the fly gut during the procyclic phase.
The VSG coat
The surface of the trypanosome is covered by a dense coat of ~1x107 molecules of Variable Surface
Glycoprotein (VSG) [Barry DJ & McCulloch R. Antigenic variation in trypanosomes: enhanced phenotypic variation in a eukaryotic parasite. Adv Parasitol (2001) vol. 49 pp. 1-70 [http://www.ncbi.nlm.nih.gov/pubmed/11461029?ordinalpos=2&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_RVDocSum] ] . This coat enables an infecting "T. brucei" population to persistently evade the host'simmune system , allowing chronic infection. The two properties of the VSG coat that allow immune evasion are:
*Shielding - the dense nature of the VSG coat prevents the immune system of the mammalian host from accessing the plasma membrane or any other invariant surfaceepitopes (such asion channels , transporters, receptors etc.) of the parasite. The coat is uniform, made up of millions of copies of the same molecule; therefore the only parts of the trypanosome the immune system can 'see' are the N-terminal loops of the VSG that make up the coat [Overath P, Chaudhri M, Steverding D, Ziegelbauer K. Invariant surface proteins in bloodstream forms of Trypanosoma brucei. Parasitol Today (Regul Ed) (1994) vol. 10 (2) pp. 53-8 [http://www.ncbi.nlm.nih.gov/pubmed/15275499?ordinalpos=8&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_RVDocSum] ] .
*Periodicantigenic variation - the VSG coat undergoes frequentstochastic genetic modification - 'switching' - allowing variants expressing a new VSG coat to escape the specific immune response raised against the previous coat.Antigenic variation
Sequencing of the "T. brucei"genome has revealed a huge "VSG" gene archive, made up of thousands of different "VSG"genes . All but one of these are 'silent' VSGs, as each trypanosome expresses only one "VSG" gene at a time. VSG is highlyimmunogenic , and an immune response raised against a specific VSG will rapidly kill trypanosomes expressing this "VSG". This can also be observedin vitro by a complement-mediatedlysis assay . However, with eachcell division there is a possibility that one or both of the progeny will switch expression to a silent "VSG" from the archive (see below). The frequency of such a switch has been measured to be approximately 1:100 [Turner CM. The rate of antigenic variation in fly-transmitted and syringe-passaged infections of Trypanosoma brucei. FEMS Microbiol Lett (1997) vol. 153 (1) pp. 227-31 [http://www.ncbi.nlm.nih.gov/pubmed/9252591?ordinalpos=1&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_RVDocSum] ] . This new VSG will likely not be recognised by the specific immune responses raised against previously expressed VSGs. It takes several days for an immune response against a specific VSG to develop, giving trypanosomes which have undergone VSG coat switching some time to reproduce (and undergo further VSG coat switching events) unhindered. Repetition of this process prevents extinction of the infecting trypanosome population, allowing chronic persistence of parasites in the host. The clinical effect of this cycle is successive 'waves' of parasitaemia (trypanosomes in the blood) [Barry DJ & McCulloch R. Antigenic variation in trypanosomes: enhanced phenotypic variation in a eukaryotic parasite. Adv Parasitol (2001) vol. 49 pp. 1-70 [http://www.ncbi.nlm.nih.gov/pubmed/11461029?ordinalpos=2&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_RVDocSum] ] .VSG structure
"VSG" genes are hugely variable at the sequence level. However, for them to fulfil their shielding function, different VSGs have strongly conserved structural features. VSGs are made up of a highly variable N terminal domain of around 300 to 350 amino acids, and a more conserved C terminal domain of around 100 amino acids. The C terminal domain forms a structural bundle of 4 alpha helices, while the N teminal domain forms a 'halo' around the helices. The
tertiary structure of this halo is well conserved between different VSGs (in spite of wide variation in amino acid sequence) allowing different VSGs to form the physical barrier required to shield the trypanosome's surface. VSG is anchored to the cell membrane via a glycophosphatidylinositol (GPI) anchor - acovalent linkage from the C terminus, to approximately 4 sugars, to aphosphatidylinositol phospholipid acid which lies in the cell membrane. VSGs formhomodimers .VSG archive structure
The "VSG" gene archive is the collection of silent VSGs in the "T. brucei"
genome . Some of these are full-length, intactgenes ; others arepseudogenes ) typically with omitted sections or premature stop codons, . Expression of an antigenically novel VSG can occur by simply switching to a different full-length VSG gene. However, only 5% of the archive is made up of such complete silent VSGs. To utilise the rest of the silent VSG archive, ‘mosaic’ VSGs can be formed by replacing part of the expressed VSG with a structurally homologous region from the archive [Marcello L, Barry JD. Analysis of the VSG gene silent archive in Trypanosoma brucei reveals that mosaic gene expression is prominent in antigenic variation and is favored by archive substructure. Genome Res (2007) vol. 17 (9) pp. 1344-52 [http://www.ncbi.nlm.nih.gov/pubmed/17652423] ] . The combinatorial nature of mosaic formation in conjunction with the huge silent VSG archive gives the parasite a theoretically limitless VSG library, and is the major barrier tovaccine development [Barbour et al. Antigenic variation in vector-borne pathogens. Emerging Infect Dis (2000) vol. 6 (5) pp. 449-57 [http://www.ncbi.nlm.nih.gov/pubmed/10998374?ordinalpos=2&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_RVDocSum] ] .VSG expression
One major focus in trypanosome research is how the majority of VSG genes are kept silent, and how these genes are switched. The expressed VSG is always located in an Expression Site - found at the
telomeres of the large and intermediate chromosomes. Each is a polycistronic unit, containing a number of Expression Site-Associated Genes (ESAGs) all expressed along with the active VSG. While there are at least 20 known expression sites, only a single one is ever active at one time. A number of mechanisms appear to be involved in this process, but the exact nature of the silencing is still unclear [E. Pays (2005). 'Regulation of antigen gene expression in Trypanosoma brucei'. Trends in Parasitology 21; 517-520 [http://www.ncbi.nlm.nih.gov/pubmed/16126458?ordinalpos=8&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_RVDocSum PMID: 16126458] ] .The VSG can be switched either by changing the active expression (from the active to a previously silent site) or by changing the VSG gene in the active site. The genome contains many copies of possible VSG genes, both on minichromosomes and in repeated sections in the interior of the chromosomes. These are generally silent, typically with omitted sections or premature stop codons, but are important in the evolution of new VSG genes. It is estimated up to 10% of the "T.brucei" genome may be made up of VSG genes or
pseudogenes . Any of these genes can be moved into the active site by recombination for expression. Again, the exact mechanisms that control this are still only partially known.Mitotic process
The mitotic division of "T.brucei" is unusual in terms of the cytoskeletal process. The basal body, unlike a
centrosome of most eukaryotic cells, plays an important role in the organisation of the spindle.Stages of mitosis:
#The basal body replicates, both remaining associated with the kinetoplast.
#The kinetoplast undergoes replication, and the daughter kinetoplasts are separated by the basal bodies.
#The second flagellum grows while the nucleus undergoes replication.
#The mitochondria divides, andcytokinesis progresses from the anterior to posterior end.
#The division resolves. The daughter cells may stay connected for a significant length of time after cytokinesis is complete.References
*K. Gull (1999). " [http://arjournals.annualreviews.org/doi/abs/10.1146%2Fannurev.micro.53.1.629 The Cytoskeleton of Trypanosomatid Parasites] ". Annual Review of Microbiology 53: 629-655. [http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=pubmed&cmd=Retrieve&dopt=AbstractPlus&list_uids=10547703&query_hl=7&itool=pubmed_DocSum PMID 10547703] .
Wikimedia Foundation. 2010.