- Disintegrin
-
Disintegrin 
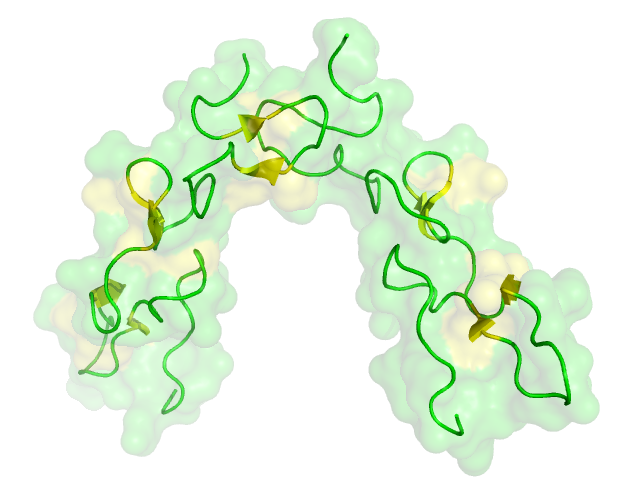
Structure of disintegrin heterodimer from Echis carinatus Identifiers Symbol Disintegrin Pfam PF00200 InterPro IPR001762 PROSITE PDOC00351 SCOP 1kst Available protein structures: Pfam structures PDB RCSB PDB; PDBe PDBsum structure summary Disintegrins are a family of small proteins (45–84 amino acids in length) from viper venoms that function as potent inhibitors of both platelet aggregation and integrin-dependent cell adhesion[1][2].
They work by countering the blood clotting steps, inhibiting the clumping of platelets. They interact with the beta-1 and -3 families of integrins receptors. Integrin are cell receptors involved in cell–cell and cell–extracellular matrix interactions, serving as the final common pathway leading to aggregation via formation of platelet–platelet bridges, which are essential in thrombosis and haemostasis. Disintegrins contain an RGD (Arg-Gly-Asp) or KGD (Lys-Gly-Asp) sequence motif that binds specifically to integrin IIb-IIIa receptors on the platelet surface, thereby blocking the binding of fibrinogen to the receptor–glycoprotein complex of activated platelets. Disintegrins act as receptor antagonists, inhibiting aggregation induced by ADP, thrombin, platelet-activating factor and collagen[3]. The role of disintegrin in preventing blood coagulation renders it of medical interest, particularly with regard to its use as an anti-coagulant[4].
Disintegrins from different snake species have been characterised: albolabrin, applagin, barbourin, batroxostatin, bitistatin, obtustatin[5], schistatin[6], echistatin[7], elegantin, eristicophin, flavoridin[8], halysin, kistrin, tergeminin, salmosin[9] and triflavin.
Disintegrin-like proteins are found in various species ranging from slime mold to humans. Some other proteins known to contain a disintegrin domain are:
- Some snake venom zinc metalloproteinases[10] consist of an N-terminal catalytic domain fused to a disintegrin domain. Such is the case for trimerelysin I (HR1B), atrolysin-e (Ht-e) and trigramin. It has been suggested that these proteinases are able to cleave themselves from the disintegrin domains and that the latter may arise from such a post-translational processing.
- The beta-subunit of guinea pig sperm surface protein PH30[11]. PH30 is a protein involved in sperm-egg fusion. The beta subunit contains a disintegrin at the N-terminal extremity.
- Mammalian epididymial apical protein 1 (EAP I)[12]. EAP I is associated with the sperm membrane and may play a role in sperm maturation. Structurally, EAP I consists of an N-terminal domain, followed by a zinc metalloproteinase domain, a disintegrin domain, and a large C-terminal domain that contains a transmembrane region.
- ADAM and ADAMTS protein families, which include important protease enzymes. As an example, the secreted protease ADAMTS13, found in serum, cleaves Von Willebrand factor and acts as a natural, endogenous, inhibitor of platelet adhesion and aggregation.
References
- ^ McLane MA, Sanchez EE, Wong A, Paquette-Straub C, Perez JC (2004). "Disintegrins". Curr Drug Targets Cardiovasc Haematol Disord 4 (4): -. PMID 15578957.
- ^ Lu X, Lu D, Scully MF, Kakkar VV (2005). "Snake venom metalloproteinase containing a disintegrin-like domain, its structure-activity relationships at interacting with integrins". Curr Med Chem Cardiovasc Hematol Agents 3 (3): -. PMID 15974889.
- ^ Rahman S, Xu CS (2001). "Identification by Site-directed Mutagenesis of Amino Acid Residues Flanking RGD Motifs of Snake Venom Disintegrins for Their Structure and Function". Acta Biochim. Biophys. Sin. 33 (2): -. PMID 12050803.
- ^ Lu X, Lu D, Scully MF, Kakkar VV (2006). "Integrins in drug targeting-RGD templates in toxins". Curr Pharm Des 12 (22): -. PMID 16918409.
- ^ Calvete JJ, Monleon D, Celda B, Paz Moreno-Murciano M, Marcinkiewicz C (2003). "NMR solution structure of the non-RGD disintegrin obtustatin". J. Mol. Biol. 329 (1): -. doi:10.1016/S0022-2836(03)00371-1. PMID 12742023.
- ^ Betzel C, Sharma S, Singh TP, Perbandt M, Yadav S, Kaur P, Bilgrami S (2005). "Crystal structure of the disintegrin heterodimer from saw-scaled viper (Echis carinatus) at 1.9 A resolution". Biochemistry 44 (33): -. doi:10.1021/bi050849y. PMID 16101289.
- ^ Calvete JJ, Kovacs H, Monleon D, Celda B, Esteve V (2005). "Conformation and concerted dynamics of the integrin-binding site and the C-terminal region of echistatin revealed by homonuclear NMR". Biochem J 387 (Pt 1): -. doi:10.1042/BJ20041343. PMC 1134932. PMID 15535803. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1134932.
- ^ Mizuno H, Morita T, Fujii Y, Fujimoto Z, Horii K, Okuda D (2003). "Crystal structure of trimestatin, a disintegrin containing a cell adhesion recognition motif RGD". J. Mol. Biol. 332 (5): -. doi:10.1016/S0022-2836(03)00991-4. PMID 14499613.
- ^ Lee W, Shin J, Kang I, Hong SY, Chung K, Jang Y, Kim DS (2003). "Solution structure of a novel disintegrin, salmosin, from Agkistrondon halys venom". Biochemistry 42 (49): -. doi:10.1021/bi0300276. PMID 14661951.
- ^ Teixeira Cde .F, Fernandes CM, Zuliani JP, Zamuner SF (2005). "Inflammatory effects of snake venom metalloproteinases". Mem. Inst. Oswaldo Cruz 100: -. PMID 15962120.
- ^ Turck CW, Myles DG, Primakoff P, Blobel CP, Wolfsberg TG, White JM (1992). "A potential fusion peptide and an integrin ligand domain in a protein active in sperm-egg fusion". Nature 356 (6366): 248–252. doi:10.1038/356248a0. PMID 1552944.
- ^ Hall L, Jones R, Barker PJ, Perry AC (1992). "A mammalian epididymal protein with remarkable sequence similarity to snake venom haemorrhagic peptides". Biochem. J. 286: 671–675. PMC 1132955. PMID 1417724. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1132955.
See also
External links

This protein-related article is a stub. You can help Wikipedia by expanding it.