- Oligopeptidase
-
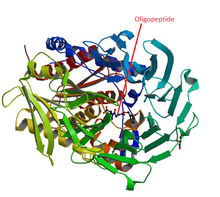
Oligopeptidase is an enzyme that cleaves peptides but not proteins, a property that is due to its structure: the active site of this enzyme is located at the end of a narrow cavity which can only be reached by peptides. These oligopeptides, peptides, predominantely smaller than 30 amino acids in length, play essential roles as hormones, in the surveillance against pathogens, and in neurological activities. Therefore these molecules constantly need to be specifically generated and inactivated, which is the role of the oligopeptidases. Oligopeptidase is a term coined in 1979 to designate a sub-group of the endopeptidases[1][2], which are not involved in the digestion nor in the processing of proteins like the pancreatic enzymes, proteasomes, cathepsins among many others. The prolyl-oligopeptidase or prolyl endopeptidase (POP) is a good example of how an oligopeptidase interacts with and metabolizes an oligopeptide. The peptide has first to penetrate into a 4 Å hole on the surface of the enzyme in order to reach an 8,500Å3 internal cavity, where the active site is located.[3][4]. Would any oligopeptide be susceptible to the action of the oligopeptidase? No. The activity of the oligopeptidase is rather selective, as experimental evidences demonstrated. They suggest that the oligopeptide should display specific structures in order to access the active site of the enzyme[5]. This peculiar mechanism of action sets the oligopeptidases at the end of the protein processing pathway, acting on specific oligopeptides, allowing them to play essential physiological and pathological roles, from microorganisms to man.
Contents
Historical Background
Autolysis:Proteins are essential macromolecules of living organisms. They are continuously been degraded into their constituent amino acids which can be reused in the synthesis of new proteins. Every cellular protein has its own half-life time. In humans, for instance, 50% of the liver and plasma proteins are replaced in 10 days, whereas in muscles it takes 180 days. In average, every 80 days about 50% of our proteins are totally replaced.[6] Although the regulation of the protein degradation is as important as their synthesis to keep each cell protein concentration at the optimum level, this research area remained neglected until the end of 1970s. Up to this time, lysosomes, discovered in the1950s by the Belgian cytologist Christian de Duve were held responsible for the complete digestion of intra- and extracellular proteins by the lysosomal hydrolytic enzymes. Limited proteolysis: Between the 1970s and 1980s, this view drastically changed. New experimental evidences showed that, under physiological conditions, non-lysosomal proteases were responsible for limited proteolysis of intra- and/or extracellular proteins, a concept originally conceived by Linderstᴓm-Lang in 1950.[7] Endogenous or exogenous proteins are processed by non-lysosomal proteases into intermediate-sized polypeptides, which display gene and metabolic regulation, neurologic, endocrine, and immunological roles, whose dysfunction might explain a number of pathologies. Consequently, protein degradation did not represent anymore the end of the biological function of proteins, but rather the beginning of a yet unexplored side of the biology of the cells. A number of intra- or extracellular proteases release protein fragments endowed with essential biological activities. These hydrolytic processes could be carried out by proteases such as Proteasomes, Proprotein Convertases,[8] Caspases, Rennin and Kallikreins. Among the products released by the non-lysosomal proteases are the bioactive oligopeptides such as hormones, neuropeptides and epitopes that, once released, could be modulated in their biological activities by specific peptidases, which promote the trimming, conversion and/or inactivation of the bioactive oligopeptides.
The history of oligopeptidases originates in the late 1960s, when the rabbit brain was searched for enzymes that cause inactivation of the nonapeptide bradykinin.[9] In the early and mid 1970s two thiol-activated endopeptidases, responsible for more than 90% of bradykinin inactivation, were isolated from cytosol of rabbit brain, and characterized.[10][11] They correspond to EOPA (endooligopeptidase A, EC 3.4.22.19), and Prolyl endopeptidase or Prolyl oligopeptidase (POP) (EC 3.4.21.26). Since their activities are restricted to oligopeptides (usually from 8-13 amino acid residues), and do not hydrolyze proteins or large peptides (>30 amino acid residues), they were designated oligopeptidases.[1] In the early and mid 1980s other oligopeptidases, mostly metallopeptidases, were described in the cytosol of mammalian tissues, such as the TOP (thimet oligopeptidase, EC 3.4.24.15),[12] and the neurolysin (EC 3.4.24.16).[13] Earlier on, the ACE (angiotensin-converting enzyme, EC 3.4.15.1), and the NEP (neprilysin, EC 3.4.24.11), had been described, at the end of the 1960s,[14] and in 1973,[15] respectively.
Function and clinical significance
Since the discovery of the neuropeptides and peptide hormones from the central nervous system (ACTH, β-MSH, endorphin, oxytocin, vasopressin, LHRH, enkephalins, substance P), and of peripheral vasoactive peptides (angiotensin, bradykinin) around the middle of last century, the number of biologically active peptides has exponentially increased. They are signaling molecules, participating in all essential aspects of life, from physiological homeostasis (as neuropeptides, peptide hormones, vasoactive peptides), to immunological defense (as MHC class I and II, cytokinins), and as regulatory peptides displaying more than a single action. These peptides result from partial proteolysis of intracellular or extracellular protein precursors performed by several processing enzymes or protease complexes (rennin, kallikreins, calpains, prohormone convertases, proteasomes, endosomes, lysosomes), which convert proteins into peptides, including those with biological activities. The resulting protein fragments of various sizes are either readily degraded into free amino acids,[16] or captured by oligopeptidases, whose peculiar binding and/or catalytic properties allow them to fulfill their physiological roles by trimming inactive peptide precursors leading to their active form,[14][17] converting bioactive peptides into novel ones.,[18] inactivating them, thus restraining the continuous activation of specific receptors,[10][11] or protecting the newly generated bioactive peptide from further degradation, suggesting a peptide chaperon-like activity.[19][20] TOP, an ubiquitous cytosolic oligopeptidase, is a remarkable example of how this enzyme could play an essential role in immune defense against cancer cells.[17] It has also been successfully used as a hook to fish novel bioactive peptides from cytosol of cells.[21]
The involvement of peptides in cell-cell interactions and in neuropsychiatric, autoimmune, and neurovegetative diseases are waiting for peptidomics[22] and gene silencing approaches, which will expedite the formation of new concepts in an emerging era for oligopeptidases.
The participation of oligopeptidases in a number of pathologies has long been reported. The ACE has benefited the most from a thorough knowledge on the enzyme structure and its mechanism of catalysis leading to the better understanding of its role in cardiovascular pathologies and therapeutics. Accordingly, for over 30 years, the treatment of human arterial hypertension has taken advantage of ACE inhibition by active site-directed inhibitors like captopril, enalapril, lisinopril, and others.[23] For the other oligopeptidases, especially those involved in human diseases, the existing studies are promising but not yet as developed as for the ACE. Some examples are: a) the POP of nervous tissues has been suggested to be involved in neuropsychiatric disorders, like in post-traumatic stress, depression, mania, nervous bulimia, anorexia, and schizophrenia, as reviewed in.[4] b) NEP has been involved in cancer;[24] c) the TOP has been involved in tuberculosis[20] and in cancer;[17] d) the EOPA or NUDEL/EOPA (NDEL1/EOPA gene product) has been involved in neuronal migration during the cortex formation in human embryo (lissencephaly) and neurite outgrowth in adults, as in schizophrenia.[25][26] Concidently, an activity related to the development of nervous tissue has been suggested for POP, nevertheless not involving its proteolytic activity.[27]
References
- ^ a b [Camargo ACM, Caldo H, Reis ML (1979) Susceptibility of a peptide derived from bradykinin to hydrolysis by brain endo-oligopeptidases and pacreatic proteinases. J Biol Chem 254, 5304-5307. PMID: 447650]
- ^ [Barrett AJ and Rawlings ND (1992). Oligopeptidases, and the emergence of the prolyl oligopeptidase family. Chem Hoppe-Seyler 373, 353-360. PMID: 1515061]
- ^ a b [ Fülöp V, Böcskei Z, Polgár L (1998). Prolyl oligopeptidase: an unusual beta propeller domain regulates proteolysis. Cell 94, 161-170. PMID: 9695945]
- ^ a b [ Li M, Chen C, Davies DR, Chiu TK (2010). Induced-fit mechanism for prolyl endopeptidase. J Biol Chem 285, 21487-21495. doi: 10.1074/jbc.M109.092692]
- ^ [Jacchieri SG, Gomes MD, Juliano L, Camargo ACM (1998). A comparative conformational analysis of thimet oligopeptidase (EC.3.4.2.4.15) substrates. J Pept Res 51, 452-459. PMID: 9650720]
- ^ Bachmair A, Finley D, Varshavsky A (Oct 1986), "In vivo half-life of a protein is a function of its amino-terminal residue", Science 243 (4773): 179-86, PMID 3018930
- ^ Linderstrøm-Lang K (1950), "In vivo half-life of a protein is a function of its amino-terminal residue", Cold Spring Harb Symp Quant Biol 14: 117-26, PMID 15442905
- ^ Seidah NG (Mar 2011), "What lies ahead for the proprotein convertases?", Ann N Y Acad. Sci 1220: 149-61, doi:10.1111/j.1749-6632.2010.05883.x, PMID 21388412
- ^ Camargo ACM, Graeff F (1969).Subcellular distribution and properties of the bradykinin inactivation system in rabbit brain homogenates. Biochem Pharmacol 18, 548-549. PMID: 5778163
- ^ a b [Camargo ACM, Shapanka R, Greene LJ (1973). Preparation, assay and partial characterization of a neutral endopeptidase from rabbit brain. Biochemistry 12, 1838-1844, 1973. PMID: 4699240]
- ^ a b [Oliveira EB, Martins AR, Camargo ACM. 1976. Isolation of brain endopeptidases: Influence of size and sequence of substrates structurally related to bradykinin. Biochemistry 15, 1967-1974. PMID: 5120]
- ^ [ Orlowski M, Michaud C, Chu TG (1983). A soluble metalloendopeptidase from rat brain. Purification of the enzyme and determination of specificity with synthetic and natural peptides. Eur J Biochem 135, 81–88. Doi: 10.1111/j.1432-1033.1983.tb07620.x]
- ^ [ Checler F, Vincent JP, Kitabgi P (1983). Degradation of neurotensin by synaptic membranes: involvement of a thermolysin-like metalloendopeptidase (enkephalinase), angiotensin-converting enzyme and other unidentified peptidases. J Neurochem 41, 375-384. PMID: 6308159]
- ^ a b [1]
- ^ [Kerr MA, Kenny AJ (1974). The purification and specificity of a neutral endopeptidase from rabbit kidney brush border. Biochem J 137, 477–488. PMID: 4423492]
- ^ [Goldberg AL (2003) Protein degradation and protection against misfolded or damaged proteins. Nature 426, 895–899. Doi:10.1038/nature02263]
- ^ a b c [ Kessier JH, Khan S, Seifert U et al. (2011) Antigen processing by nardilysin and thimet oligopeptidase generates cytotoxic T cell epitopes. Nature Immunology- http://www.nature.com/ni/journal/v12/n1/full/ni.1974.html]
- ^ [Camargo AC, Oliveira EB, Toffoletto O et al (1997). Brain endo-oligopeptidase A, a putative enkephalin converting enzyme. J Neurochem 48, 1258-1263. PMID: 2880931]
- ^ [ Portaro F, Gomes MD, Cabrera A et al (1999). Thimet oligopeptidase and the stability of MHC class I epitopes in macrophage cystosol. Biochem. Biophys. Res. Commum. 255, 596-601. Doi:10.1006/bbrc.1999.0251]
- ^ a b [Silva CL, Portaro FCV, Bonato VLD et al (1999). Thimet Oligopeptidase (EC.3.4.24.15) is a novel protein on the route of MHC class I antigen presentation. Biochem Biophys Res Commun 255, 591-595. Doi: 10.1006/bbrc.1999.0250]
- ^ [ Gelman JS, Sironi J, Castro LM, et al (2010). Hemopressins and other hemoglobin-derived peptides in mouse brain: comparison between brain, blood, and heart peptidome and regulation in Cpefat/fat mice. J. Neurochem 113, 871–880. Doi: 10.1111/j.1471-4159.2010.06653.x]
- ^ [Gelman JS, Sironi J, Castro LM et al (2011). Peptidomic analysis of human cell lines. J Proteome Res. Feb 17. Doi:10.1021/pr100952f]
- ^ [2]
- ^ [Maguer-Satta V, Besançon R, Bachelard-Cascale E (2011). Concise Review: Neutral Endopeptidase (NEP/CD10): a Multifaceted Environment Actor in Stem Cells, Physiological Mechanisms and Cancer. Stem Cells Jan 7; see www.StemCells.com for supporting information available online]
- ^ [ Hayashi MA, Portaro FC, Bastos MF et al. (2005). Inhibition of NUDEL (nuclear distribution element-like)-oligopeptidase activity by disrupted-in-schizophrenia 1. Proc. Natl. Acad. Sci 102 (10): 3828-3833. Doi:10.1073/pnas.0500330102]
- ^ [Hayashi MA, Guerreiro JR, Charych E et al. (2010). Assessing the role of endooligopeptidase activity of Ndel1 (nuclear-distribution gene E homolog like-1)in neurite outgrowth Mol Cell Neurosci. 44, 353-361. doi:10.1016/j.m.cn.2010.04.006]
- ^ [Hannula MJ, Männistö PT, Myöhänen TT (2010). Sequential Expression, Activity and Nuclear Localization of Prolyl Oligopeptidase Protein in the Developing Rat Brain Dev Neurosci DOI: 10.1159/000322082]
External links
Categories:- Hydrolases
- Neuroscience
- EC 3.4
- Peptidase
- Metabolism
- Immunology
Wikimedia Foundation. 2010.