- Photophosphorylation
-
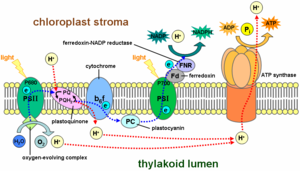 Photophosphorylation through light-dependent reactions of photosynthesis at the thylakoid membrane
Photophosphorylation through light-dependent reactions of photosynthesis at the thylakoid membrane
The production of ATP using the energy of sunlight is called photophosphorylation. Only two sources of energy are available to living organisms: sunlight and reduction-oxidation (redox) reactions. All organisms produce ATP, which is the universal energy currency of life.
In photophosphorylation, light energy is used to create a high-energy electron donor and a lower-energy electron acceptor. Electrons then move spontaneously from donor to acceptor through an electron transport chain.
Contents
Background
ATP is made by an enzyme called ATP synthase. Both the structure of this enzyme and its underlying gene are remarkably similar in all known forms of life.
ATP synthase is powered by a transmembrane electrochemical potential gradient, usually in the form of a proton gradient. The function of the electron transport chain is to produce this gradient. In all living organisms, a series of redox reactions is used to produce a transmembrane electrochemical potential gradient, or a so-called proton motive force (pmf).
Redox reactions are chemical reactions in which electrons are transferred from a donor molecule to an acceptor molecule. The underlying force driving these reactions is the Gibbs free energy of the reactants and products. The Gibbs free energy is the energy available (“free”) to do work. Any reaction that decreases the overall Gibbs free energy of a system will proceed spontaneously (given that the system is isobaric and also adiabatic)
The transfer of electrons from a high-energy molecule (the donor) to a lower-energy molecule (the acceptor) can be spatially separated into a series of intermediate redox reactions. This is an electron transport chain.
The fact that a reaction is thermodynamically possible does not mean that it will actually occur. A mixture of hydrogen gas and oxygen gas does not spontaneously ignite. It is necessary either to supply an activation energy or to lower the intrinsic activation energy of the system, in order to make most biochemical reactions proceed at a useful rate. Living systems use complex macromolecular structures to lower the activation energies of biochemical reactions.
It is possible to couple a thermodynamically favorable reaction (a transition from a high-energy state to a lower-energy state) to a thermodynamically unfavorable reaction (such as a separation of charges, or the creation of an osmotic gradient), in such a way that the overall free energy of the system decreases (making it thermodynamically possible), while useful work is done at the same time. Biological macromolecules that catalyze a thermodynamically favorable reaction if and only if a thermodynamically unfavorable reaction occurs simultaneously underlie all known forms of life.
Electron transport chains (most known as ETC) produce energy in the form of a transmembrane electrochemical potential gradient. This energy is used to do useful work. The gradient can be used to transport molecules across membranes. It can be used to do mechanical work, such as rotating bacterial flagella. It can be used to produce ATP and NADPH, high-energy molecules that are necessary for growth.
Cyclic photophosphorylation
In cyclic electron flow, the electron begins in a pigment complex called photosystem I, passes from the primary acceptor to ferredoxin, then to cytochrome b6f (a similar complex to that found in mitochondria), and then to plastocyanin before returning to chlorophyll. This transport chain produces a proton-motive force, pumping H+ ions across the membrane; this produces a concentration gradient that can be used to power ATP synthase during chemiosmosis. This pathway is known as cyclic photophosphorylation, and it produces neither O2 nor NADPH. Unlike non-cyclic photophosphorylation, NADP+ does not accept the electrons, but they are sent back to photosystem I. NADPH is NOT produced in cyclic photophosphorylation. In bacterial photosynthesis, a single photosystem is used, and therefore is involved in cyclic photophosphorylation. It is favoured in anaerobic conditions and conditions of high irradiance and CO2 compensation point.
Noncyclic photophosphorylation
The other pathway, noncyclic photophosphorylation, is a two-stage process involving two different chlorophyll photosystems. Being a light reaction, Noncyclic photophosphorylation occurs on thylakoid membranes inside chloroplasts. First, a water molecule is broken down into 2H+ + 1/2 O2 + 2e- by a process called photolysis (or light-splitting). The two electrons from the water molecule are kept in photosystem II, while the 2H+ and 1/2O2 are left out for further use. Then a photon is absorbed by chlorophyll pigments on surrounding the reaction core center of the photosystem. The light excites the electrons of each pigment, causing a chain reaction that eventually transfers energy to the core of photosystem II, exciting the two electrons that are transferred to the primary electron acceptor, pheophytin. The deficit of electrons is replenished by taking electrons from another molecule of water. The electrons transfer from pheophytin to plastoquinone, then to plastocyanin, providing the energy for hydrogen ions (H+) to be pumped into the thylakoid space. This creates a gradient, making H+ ions flow back into the stroma of the chloroplast, providing the energy for the regeneration of ATP.
The photosystem II complex replaced its lost electrons from an external source; however, the two other electrons are not returned to photosystem II as they would in the analogous cyclic pathway. Instead, the still-excited electrons are transferred to a photosystem I complex, which boosts their energy level to a higher level using a second solar photon. The highly excited electrons are transferred to the acceptor molecule, but this time are passed on to an enzyme called Ferredoxin- NADP reductase|NADP+ reductase, for short FNR, which uses them to catalyse the reaction (as shown):
- NADP+ + 2H+ + 2e- → NADPH + H+
This consumes the H+ ions produced by the splitting of water, leading to a net production of 1/2O2, ATP, and NADPH+H+ with the consumption of solar photons and water.
The concentration of NADPH in the chloroplast may help regulate which pathway electrons take through the light reactions. When the chloroplast runs low on ATP for the Calvin cycle, NADPH will accumulate and the plant may shift from noncyclic to cyclic electron flow.
References
- Fenchel T, King GM, Blackburn TH. Bacterial Biogeochemistry: The Ecophysiology of Mineral Cycling. 2nd ed. Elsevier; 1998.
- Lengeler JW, Drews G, Schlegel HG, editors. Biology of the Prokaryotes. Blackwell Science; 1999.
- Nelson DL, Cox MM. Lehninger Principles of Biochemistry. 4th ed. Freeman; 2005.
- Nicholls DG, Ferguson SJ. Bioenergetics 3. Academic Press; 2002.
- Stumm W, Morgan JJ. Aquatic Chemistry. 3rd ed. Wiley; 1996.
- Thauer RK, Jungermann K, Decker K. Energy Conservation in Chemotrophic Anaerobic Bacteria. Bacteriol. Rev. 41:100-180; 1977.
- White D. The Physiology and Biochemistry of Prokaryotes. 2nd ed. Oxford University Press; 2000.
- Voet D, Voet JG. Biochemistry. 3rd ed. Wiley; 2004.
- Cj C. Enverga
Categories:- Photosynthesis
- Light reactions
Wikimedia Foundation. 2010.