- Bacterial cell structure
Bacteria , despite their apparent simplicity, contain a well developed cell structure which is responsible for many of their unique biological properties. Many structural features are unique tobacteria and are not found amongarchaea oreukaryotes . Because of the simplicity ofbacteria relative to larger organisms and the ease with which they can be manipulated experimentally, the cell structure ofbacteria has been well studied, revealing many biochemical principles that have been subsequently applied to other organisms.Cell morphology
Perhaps the most elemental structural property of
bacteria is cell morphology (shape). Typical examples include:
*coccus (spherical)
* bacillus (rod-like)
*spirillum (spiral)
* filamentousCell shape is generally characteristic of a given bacterial species, but can vary depending on growth conditions. Some bacteria have complex life cycles involving the production of stalks and appendages (e.g. "Caulobacter ") and some produce elaborate structures bearing reproductive spores (e.g. "Myxococcus ", "Streptomyces ").Bacteria generally form distinctive cell morphologies when examined by light microscopy and distinct colony morphologies when grown on Petri plates. These are often the first characteristics observed by amicrobiologist to determine the identity of an unknownbacterial culture .The importance of cell size
Perhaps the most obvious structural characteristic of
bacteria is (with some exceptions) their small size. For example, "Escherichia coli ", an "average" sized bacterium with average cell length of ca. 1 µm has a cell volume of approximately 1 - 2 μm3. This corresponds to a wet mass of ca. 1 pg, assuming that the cell consists mostly of water. The dry mass of a single cell can be estimated as 20 % of the wet mass, amounting to 0.2 pg. About half of the dry mass of a bacterial cell consists of carbon, and also about half of it can be attributed to proteins. Therefore, a typical fully grown 1-liter culture of "Escherichia coli " (at an optical density of 1.0, corresponding to ca. 109 cells/ml) yields ca. 1 g wet cell mass.Small size is extremely important because it allows for a large
surface area-to-volume ratio which allows for rapid uptake and intracellular distribution of nutrients and excretion of wastes. At low surface area-to-volume ratios the diffusion of nutrients and waste products across the bacterial cell membrane limits the rate at which microbial metabolism can occur, making the cell less evolutionarily fit. The reason for the existence of large cells is unknown, although it is speculated that the increased cell volume is used primarily for storage of excess nutrients.The bacterial cell wall
As in other organisms, the bacterial
cell wall provides structural integrity to the cell. Inprokaryotes , the primary function of the cell wall is to protect the cell from internalturgor pressure caused by the much higher concentrations of proteins and other molecules inside the cell compared to its external environment. The bacterial cell wall differs from that of all other organisms by the presence ofpeptidoglycan (poly-"N"-acetylglucosamine and "N"-acetylmuramic acid), which is located immediately outside of thecytoplasmic membrane .Peptidoglycan is responsible for the rigidity of the bacterial cell wall and for the determination of cell shape. It is relatively porous and is not considered to be a permeability barrier for small substrates. While all bacterial cell walls (with a few exceptions e.g. intracellular parasites such as "Mycoplasma ") contain peptidoglycan, not all cell walls have the same overall structures. There are two main types of bacterial cell walls, Gram positive and Gram negative, which are differentiated by theirGram staining characteristics. For both Gram-positive and Gram-negative bacteria, particles of approximately 2 nm can pass through the peptidoglycan. [cite journal | author=Demchick PH and Koch AL | title=The permeability of the wall fabric of Escherichia coli and Bacillus subtilis | journal=Journal of Bacteriology | year=1996 | pages=768–73| volume=178 | issue=3 [http://jb.asm.org/cgi/reprint/178/3/768] ]The Gram positive cell wall
The
Gram positive cell wall is characterized by the presence of a very thickpeptidoglycan layer, which is responsible for the retention of the crystal violet dyes during theGram staining procedure. It is found exclusively in organisms belonging to theActinobacteria (or high %G+C Gram positive organisms) and theFirmicutes (or low %G+CGram positive organisms). Bacteria within theDeinococcus-Thermus group may also exhibitGram positive staining behaviour but contain some cell wall structures typical ofGram negative organisms. Embedded in the Gram positive cell wall are polyalcohols calledteichoic acid s, some of which are lipid-linked to form lipoteichoic acids. Because lipoteichoic acids are covalently linked to lipids within thecytoplasmic membrane they are responsible for linking thepeptidoglycan to thecytoplasmic membrane .Teichoic acid s give theGram positive cell wall an overall negative charge due to the presence ofphosphodiester bonds betweenteichoic acid monomers.The Gram negative cell wall
Unlike the Gram positive cell wall, the Gram negative cell wall contains a thin
peptidoglycan layer adjacent to thecytoplasmic membrane , which is responsible for the cell wall's inability to retain the crystal violet stain upon decolourisation with ethanol duringGram staining . In addition to thepeptidoglycan layer, the Gram negative cell wall also contains an additional outer membrane composed byphospholipid s andlipopolysaccharide s which face into the external environment. As thelipopolysaccharide s are highly-charged, the Gram negative cell wall has an overall negative charge. The chemical structure of the outer membranelipopolysaccharide s is often unique to specific bacterial strains (i.e. sub-species) and is responsible for many of theantigen ic properties of these strains.The bacterial cytoplasmic membrane
The bacterial cytoplasmic membrane is composed of a
phospholipid bilayer and thus has all of the general functions of acell membrane such as acting as a permeability barrier for most molecules and serving as the location for the transport of molecules into the cell. In addition to these functions,prokaryotic membranes also function in energy conservation as the location about which aproton motive force is generated. Unlikeeukaryotes , bacterial membranes (with some exceptions e.g. "Mycoplasma " and methanotrophs) generally do not containsterols . However, many microbes do contain structurally related compounds calledhopanoid s which likely fulfill the same function. Unlikeeukaryotes ,bacteria can have a wide variety offatty acid s within their membranes. Along with typical saturated and unsaturatedfatty acid s, bacteria can contain fatty acids with additionalmethyl ,hydroxy or even cyclic groups. The relative proportions of these fatty acids can be modulated by the bacterium to maintain the optimum fluidity of the membrane (e.g. following temperature change).As a
phospholipid bilayer , the lipid portion of the outer membrane is impermeable to charged molecules. However, channels called porins are present in the outer membrane that allow forpassive transport of manyion s,sugar s andamino acid s across the outer membrane. These molecules are therefore present in theperiplasm , the region between the cytoplasmic and outer membranes. Theperiplasm contains the peptidoglycan layer and many proteins responsible for substrate binding orhydrolysis and reception of extracellular signals. The periplasm it is thought to exist as a gel-like state rather than a liquid due to the high concentration of proteins andpeptidoglycan found within it. Because of its location between the cytoplasmic and outer membranes, signals received and substrates bound are available to be transported across thecytoplasmic membrane using transport and signalling proteins imbedded there.Other bacterial surface structures
Fimbrae and Pili
"Main article:"
Pilus Fimbrae are protein tubes that extend out from the outer membrane in many members of the
Proteobacteria . They are generally short in length and present in high numbers about the entire bacterial cell surface. Fimbrae usually function to facilitate the attachment of abacterium to a surface (e.g. to form abiofilm ) or to other cells (e.g. animal cells duringpathogenesis )). A few organisms (e.g. "Myxococcus ") use fimbrae for motility to facilitate the assembly of multicellular structures such as fruiting bodies. Pili are similar in structure to fimbrae but are much longer and present on the bacterial cell in low numbers. Pili are involved in the process ofbacterial conjugation . Non-sex pili also aid bacteria in gripping surfaces.-layers
"Main article:"
S-layer An
S-layer is a cell surface protein layer found in many differentbacteria and in somearchaea where it serves as the cell wall. AllS-layer s are made up of a two-dimensional array of proteins and have a crystalline appearance, the symmetry of which differs between species. The exact function ofS-layer s is unknown, but it has been suggested that they act as a partial permeability barrier for large substrates. For example, anS-layer could conceivably keep extracellular proteins near the cell membrane by preventing their diffusion away from the cell. In some pathogenic species, anS-layer may help to facilitate survival within the host by conferring protection against host defence mechanisms.Capsules and Slime Layers
"Main article:"
Slime layer Many
bacteria secrete extracellular polymers outside of their cell walls. These polymers are usually composed ofpolysaccharide s and sometimesprotein . Capsules are relatively impermeable structures that cannot be stained with dyes such asIndia ink . They are structures that help protectbacteria fromphagocytosis anddesiccation .Slime layer s are somewhat looser, fibrous structures generally involved in attachment ofbacteria to other cells or inanimate surfaces to formbiofilm s. Slime layers can also be used as a food reserve for the cell.
*An example of how a bacterial cell uses their slime layer to attach to a surface is in the Streptococcus mutans. Streptococcus mutans attaches to the teeth with a slime layer and forms a sticky film that traps food particles and other bacteria on the teeth (dental plaque). The bacteria then metabolizes the trapped food particles and release acids (thus possibly causing tooth decay).[
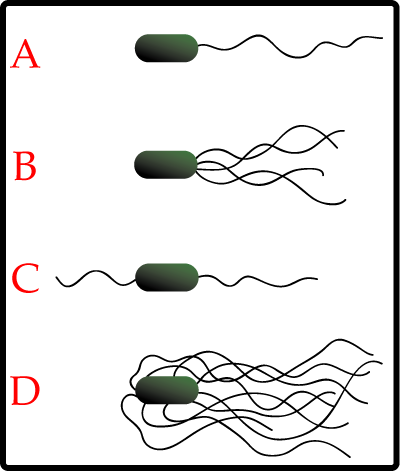 thumb|200px|right
thumb|200px|right
A-Monotrichous; B-Lophotrichous; C-Amphitrichous; D-Peritrichous; ]Flagella
"Main article:"
Flagellum Perhaps the most recognizable extracellular bacterial cell structures are
flagella .Flagella are whip-like structures protruding from the bacterial cell wall and are responsible for bacterialmotility (i.e. movement). The arrangement of flagella about the bacterial cell is unique to the species observed. Common forms include:
*Peritrichous - Multiple flagella found at several locations about the cell
*Polar - Single flagella found at one or both cell poles
*Lophotrichous - A tuft of flagella found at one cell poleFlagella are complex structures that are composed of many different proteins. These includeflagellin , which makes up the whip-like tube and aprotein complex that spans the cell wall and cell membrane to form a motor that causes theflagellum to rotate. This rotation is normally driven byproton motive force and are found in the body of the cell.Intracellular bacterial cell structures
In comparison to
eukaryotes , the intracellular features of the bacterial cell are extremely simplistic. Bacteria do not containorganelle s in the same sense aseukaryote s. Instead, thechromosome and perhapsribosomes are the only easily observable intracellular structures found in allbacteria . There do exist, however, specialized groups of bacteria that contain more complex intracellular structures, some of which are discussed below.The bacterial chromosome and plasmids
"Main article:"
Plasmid Unlike
eukaryotes , the bacterialchromosome is not enclosed inside of a membrane-bound nucleus but instead resides inside the bacterialcytoplasm . This means that the transfer of cellular information through the processes oftranslation , transcription andDNA replication all occur within the same compartment and can interact with other cytoplasmic structures, most notablyribosome s. The bacterial chromosome is not packaged usinghistones to formchromatin as ineukaryote s but instead exists as a highly compactsupercoil ed structure, the precise nature of which remains unclear. Most bacterial chromosomes are circular although some examples of linear chromosomes exist (e.g. "Borrelia burgdorferi "). Along with chromosomal DNA, most bacteria also contain small independent pieces of DNA calledplasmid s that often encode for traits that are advantageous but not essential to their bacterial host.Plasmid s can be easily gained or lost by a bacterium and can be transferred between bacteria as a form ofhorizontal gene transfer .Ribosomes and other multiprotein complexes
"Main article:"
Ribosome In most
bacteria the most numerous intracellular structure is theribosome , the site of protein synthesis in all living organisms. Allprokaryotes have 70S (where S=Svedberg units) ribosomes whileeukaryotes contain larger 80Sribosome s in theircytosol . The 70Sribosome is made up of a 50S and 30S subunits. The 50S subunit contains the 23S and 5SrRNA while the 30S subunit contains the 16SrRNA . TheserRNA molecules differ in size ineukaryotes and are complexed with a large number of ribosomal proteins, the number and type of which can vary slightly between organisms. While theribosome is the most commonly observed intracellular multiprotein complex inbacteria other large complexes do occur and can sometimes be seen usingmicroscopy .Intracellular membranes
While not typical of all
bacteria some microbes contain intracellular membranes in addition to (or as extensions of) their cytoplasmic membranes. An early idea was that bacteria might contain membrane folds termedmesosome s, but these were later shown to be artifacts produced by the chemicals used to prepare the cells forelectron microscopy . [cite journal |author=Ryter A |title=Contribution of new cryomethods to a better knowledge of bacterial anatomy |journal=Ann. Inst. Pasteur Microbiol. |volume=139 |issue=1 |pages=33–44 |year=1988 |pmid=3289587] Examples ofbacteria containing intracellular membranes arephototroph s,nitrifying bacteria andmethane -oxidising bacteria. Intracellular membranes are also found inbacteria belonging to the poorly studiedPlanctomycetes group, although these membranes more closely resemble organellar membranes ineukaryotes and are currently of unknown function. [cite journal |author=Fuerst J |title=Intracellular compartmentation in planctomycetes |journal=Annu Rev Microbiol |volume=59 |pages=299–328 |year=2005 |pmid=15910279 |doi=10.1146/annurev.micro.59.030804.121258]Cytoskeleton
The prokaryotic cytoskeleton is the collective name for all structural filaments in
prokaryotes . It was once thought that prokaryotic cells did not possess cytoskeletons, but recent advances in visualization technology and structure determination have shown that filaments indeed exist in these cells.cite journal |author=Gitai Z |title=The new bacterial cell biology: moving parts and subcellular architecture |journal=Cell |volume=120 |issue=5 |pages=577–86 |year=2005 |pmid=15766522 |doi=10.1016/j.cell.2005.02.026] In fact,homologues for all major cytoskeletal proteins ineukaryotes have been found in prokaryotes. Cytoskeletal elements play essential roles incell division , protection, shape determination, and polarity determination in various prokaryotes. [cite journal |author=Shih YL, Rothfield L |title=The bacterial cytoskeleton |journal=Microbiol. Mol. Biol. Rev. |volume=70 |issue=3 |pages=729–54 |year=2006 |pmid=16959967 |url=http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pubmed&pubmedid=16959967 |doi=10.1128/MMBR.00017-06]Nutrient storage structures
Most
bacteria l habitats do not live in environments that contain large amounts of essential nutrients at all times. To accommodate these transient levels of nutrientsbacteria contain several different methods of nutrient storage in times of plenty for use in times of want. For example, manybacteria store excess carbon in the form of polyhydroxyalkanoates orglycogen . Some microbes store soluble nutrients such asnitrate invacuole s. Sulfur is most often stored as elemental (S0) granules which can be deposited either intra- or extracellularly. Sulfur granules are especially common inbacteria that usehydrogen sulfide as an electron source. Most of the above mentioned examples can be viewed using amicroscope and are surrounded by a thin nonunit membrane to separate them from thecytoplasm .Gas vesicles
Gas vesicles are spindle-shaped structures found in some
plankton ic bacteria that providesbuoyancy to these cells by decreasing their overall cell density. They are made up of a protein coat that is very impermeable to solvents such as water but permeable to mostgases . By adjusting the amount of gas present in their gas vesiclesbacteria can increase or decrease their overall cell density and thereby move up or down within the water column to maintain their position in an environment optimal for growth.Carboxysomes
"Main article:"
Carboxysome Carboxysome s are intracellular structures found in manyautotrophic bacteria such as Cyanobacteria, Knallgasbacteria, Nitroso- and Nitrobacteria. They are proteinaceous structures resembling phage heads in their morphology and contain the enzymes of carbon dioxide fixation in these organisms (especially ribulose bisphosphate carboxylase/oxygenase, RuBisCO, and carbonic anhydrase). It is thought that the high local concentration of the enzymes along with the fast conversion of bicarbonate to carbon dioxide by carbonic anhydrase allows faster and more efficient carbon dioxide fixation than possible inside the cytoplasm.Similar structures are known to harbor the coenzyme B12-containing glycerol dehydratase, the key enzyme of glycerol fermentation to 1,3-propanediol, in some Enterobacteriaceae (e. g. Salmonella).
Magnetosomes
"Main article:"
Magnetosome Magnetosome s are intracellular structures found inmagnetotactic bacteria that allow them to sense and align themselves along a magnetic field (magnetotaxis). The ecological role of magnetotaxis is unknown but it is hypothesized to be involved in the determination of optimal oxygen concentrations.Magnetosome s are composed of the mineralmagnetite and are surrounded by a nonunit membrane. The morphology ofmagnetosome s is species-specific.Endospores
"Main article:"
Endospores Perhaps the most well known bacterial adaptation to stress is the formation of
endospore s.Endospore s are bacterial survival structures that are highly resistant to many different types of chemical and environmental stresses and therefore enable the survival ofbacteria in environments that would be lethal for these cells in their normal vegetative form. It has been proposed thatendospore formation has allowed for the survival of somebacteria for hundreds of millions of years (e.g. in salt crystals) [ [http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=pubmed&cmd=Retrieve&dopt=AbstractPlus&list_uids=11057666&query_hl=11&itool=pubmed_docsum Vreeland RH, Rosenzweig WD, Powers DW. "Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal." Nature. 2000 Oct 19;407(6806):897-900.] ] [ [http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?itool=abstractplus&db=pubmed&cmd=Retrieve&dopt=abstractplus&list_uids=7538699 Cano RJ, Borucki MK. "Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber." Science. 1995 May 19;268(5213):1060-4.] ] although these publications have been questioned. [ [http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=pubmed&cmd=Retrieve&dopt=AbstractPlus&list_uids=7754393&itool=pubmed_abstractplus Fischman J. "Have 25-million-year-old bacteria returned to life?" Science. 1995 May 19;268(5213):977.] ] [ [http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=pubmed&cmd=Retrieve&dopt=AbstractPlus&list_uids=11057647&itool=pubmed_AbstractPlus Parkes RJ. "A case of bacterial immortality?" Nature. 2000 Oct 19;407(6806):844-5.] ]Endospore formation is limited to several genera of Gram-positivebacteria such as "Bacillus " and "Clostridium ". It differs from reproductive spores in that only one spore is formed per cell resulting in no net gain in cell number uponendospore germination. The location of anendospore within a cell is species-specific and can be used to determine the identity of abacterium .References
Further reading
* [http://www.microbiologytext.com/index.php?module=Book&func=displayarticlesinchapter&chap_id=35 Cell Structure and Organization]
* Madigan, M. T., Martinko, J. M. "Brock Biology of Microorganisms, 11th Ed." (2005) Pearson Prentice Hall, Upper Saddle River, NJ.External links
[http://www.blackwellpublishing.com/trun/artwork/Animations/Overview/overview.html Animated guide to bacterial cell structure.]
Wikimedia Foundation. 2010.