- Cognitive neuroscience of visual object recognition
-
Object recognition is the ability to perceive an object’s physical properties (such as shape, colour and texture) and apply semantic attributes to the object, which includes the understanding of its use, previous experience with the object and how it relates to others.[1]
Contents
Basic Stages of Object Recognition
One model of object recognition, based on neuropsychological evidence, provides information that allows us to divide the process into four different stages.[2][3][4]
- Stage 1 Processing of basic object components, such as colour, depth, and form.
- Stage 2 These basic components are then grouped on the basis of similarity, providing information on distinct edges to the visual form. Subsequently, figure-ground segregation is able to take place.
- Stage 3 The visual representation is matched with structural descriptions in memory.
- Stage 4 Semantic attributes are applied to the visual representation, providing meaning, and thereby recognition.
It should be noted that, within these stages, there are more specific processes that take place to complete the different processing components. In addition, other existing models propose integrative hierarchies (top-down and bottom-up), as well as parallel processing, as opposed to this general bottom-up hierarchy.
Hierarchical Recognition Processing
Visual recognition processing has been typically viewed as a bottom-up hierarchy in which information is processed sequentially with increasing complexities, where lower-level cortical processors, such as the primary visual cortex, are at the bottom of the processing hierarchy and higher-level cortical processors, such as the inferotemporal cortex (IT), are at the top, where recognition is facilitated.[5] A most recognized bottom-up hierarchical theory is David Marr’s theory of vision.[6] In contrast, an increasingly popular recognition processing theory, is that of top-down processing. One model, proposed by Moshe Bar (2003), describes a “shortcut” method in which early visual inputs are sent, partially analyzed, from the early visual cortex to the prefrontal cortex (PFC). Possible interpretations of the crude visual input is generated in the PFC and then sent to the inferotemporal cortex (IT) subsequently activating relevant object representations which are then incorporated into the slower, bottom-up process. This “shortcut” is meant to minimize the amount of object representations required for matching thereby facilitating object recognition.[5] Lesion studies have supported this proposal with findings of slower response times for individuals with PFC lesions, suggesting use of only the bottom-up processing.[7]
Object Constancy and Theories of Object Recognition
A significant aspect of object recognition is that of object constancy: the ability to recognize an object across varying viewing conditions. These varying conditions include object orientation, lighting, and object variability (size, colour, and other within-category differences). For the visual system to achieve object constancy, it must be able to extract a commonality in the object description across different viewpoints and the retinal descriptions.[8] Several theories have been generated to provide insight on how object constancy may be achieved for the purpose of object recognition including, viewpoint-invariant, viewpoint-dependent and multiple views theories.
Viewpoint-Invariant Theories
Viewpoint-invariant theories suggest that object recognition is based on structural information, such as individual parts, allowing for recognition to take place regardless of the object’s viewpoint. Accordingly, recognition is possible from any viewpoint as individual parts of an object can be rotated to fit any particular view.[9] This form of analytical recognition requires little memory as only structural parts need to be encoded, which can produce multiple object representations through the interrelations of these parts and mental rotation.[9] Therefore, storage of multiple object viewpoints is not required in memory.
3-D Model Representation
This model, proposed by Marr and Nishihara (1978), states that object recognition is achieved by matching 3-D model representations obtained from the visual object with 3-D model representations stored in memory. The 3-D model representations obtained from the object are formed by first identifying the concavities of the object, which separate the stimulus into individual parts. Then the axis of each individual part of the object are found. Identifying the principal axis of the object assists in the normalization process via mental rotation that is required because only the canonical description of the object is stored in memory. Recognition is acquired when the observed object viewpoint is mentally rotated to match the stored canonical description.[10]
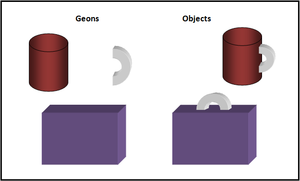 Figure 1. This image, created based on Biederman's (1987) Recognition by Components theory, is an example of how objects can be broken down into Geons.
Figure 1. This image, created based on Biederman's (1987) Recognition by Components theory, is an example of how objects can be broken down into Geons.
Recognition by Components
An extension of Marr and Nishihara's model, the Recognition by Components Theory, proposed by Biederman (1987), proposes that the visual information gained from an object is divided into simple geometric components, such as blocks and cylinders, also known as “geons” (geometric ions), and are then matched with the most similar object representation that is stored in memory to provide the object’s identification (see Figure 1).[11]
Viewpoint-Dependent Theories
Viewpoint-dependent theories suggest that object recognition is affected by the viewpoint at which it is seen, implying that objects seen in novel viewpoints reduce the accuracy and speed of object identification.[12] This theory of recognition is based on a more holistic system rather than by parts, suggesting that objects are stored in memory with multiple viewpoints and angles. This form of recognition requires a lot of memory as each viewpoint must be stored. Accuracy of recognition also depends on how familiar the observed viewpoint of the object is.[9]
Multiple Views Theory
This theory proposes that object recognition lies on a viewpoint continuum where each viewpoint is recruited for different types of recognition. At one extreme of this continuum, viewpoint-dependent mechanisms are used for within-category discriminations, while at the other extreme, viewpoint-invariant mechanisms are used for the categorization of objects.[12]
Neural Substrates
 The Dorsal Stream is shown in green and the Ventral Stream in purple.
The Dorsal Stream is shown in green and the Ventral Stream in purple.The Dorsal and Ventral Stream
The visual processing of objects in the brain can be divided into two processing pathways: the dorsal stream (how/where), which extends from the visual cortex to the parietal lobes and ventral stream (what), which extends from the visual cortex to the inferotemporal cortex (IT). The existence of these two separate visual processing pathways was first proposed by Ungerleider and Mishkin (1982) who, based on their lesion studies, suggested that the dorsal stream is involved in the processing of visual spatial information, such as object localization (where), and the ventral stream is involved in the processing of visual object identification information (what).[13] Since this initial proposal, it has been alternatively suggested that the dorsal pathway should be known as the ‘How’ pathway as the visual spatial information processed here provides us with information about how to interact with objects [14] For the purpose of object recognition, the neural focus is on the ventral stream.
Functional Specialization in the Ventral Stream
Within the ventral stream, various regions of proposed functional specialization have been observed in functional imaging studies. The brain regions most consistently found to display functional specialization are the Fusiform Face Area (FFA), which shows increased activation for faces when compared with objects, the Parahippocampal Place Area (PPA) for scenes vs. objects, the Extrastriate Body Area (EBA) for body parts vs. objects, MT+/V5 for moving stimuli vs. static stimuli, and the Lateral Occipital Complex (LOC) for discernable shapes vs. scrambled stimuli.[15] (See Also: Neural processing for individual categories of objects)
Structural Processing: The Lateral Occipital Complex
The Lateral Occipital Complex (LOC) has been found to be particularly important for object recognition at the perceptual structural level. In an event-related fMRI study that looked at the adaptation of neurons activated in visual processing of objects, it was discovered that the similarity of an object’s shape is necessary for subsequent adaptation in the LOC, but specific object features such as edges and contours are not. This suggests that activation in the LOC represents higher-level object shape information and not simple object features.[16] In a related fMRI study, the activation of the LOC, which occurred regardless of the presented object’s visual cues such as motion, texture, or luminance contrasts, suggests that the different low-level visual cues used to define an object converge in “object-related areas” to assist in the perception and recognition process.[17] None of the mentioned higher-level object shape information seems to provide any semantic information about the object as the LOC shows a neuronal response to varying forms including non-familiar, abstract objects.[18]
Further experiments have proposed that the LOC consists of a hierarchical system for shape selectivity indicating greater selective activation in the posterior regions for fragments of objects whereas the anterior regions show greater activation for full or partial objects.[19] This is consistent with previous research that suggests a hierarchical representation in the ventral temporal cortex where primary feature processing occurs in the posterior regions and the integration of these features into a whole and meaningful object occurs in the anterior regions.[20]
Semantic Processing
Through information provided from neuropsychological patients, dissociations of recognition processing have been identified between structural and semantic processing as structural, colour, and associative information can be selectively impaired. In one PET study, areas found to be involved in associative semantic processing include the left anterior superior/middle temporal gyrus and the left temporal pole comparative to structural and colour information, as well as the right temporal pole comparative to colour decision tasks only.[21] These results indicate that stored perceptual knowledge and semantic knowledge involve separate cortical regions in object recognition as well as indicating that there are hemispheric differences in the temporal regions.
Research has also provided evidence which indicates that visual semantic information converges in the fusiform gyri of the inferotemporal lobes. In a study that compared the semantic knowledge of category versus attributes, it was found that they play separate roles in how they contribute to recognition. For categorical comparisons, the lateral regions of the fusiform gyrus were activated by living objects, in comparison to nonliving objects which activated the medial regions. For attribute comparisons, it was found that the right fusiform gyrus was activated by global form, in comparison to local details which activated the left fusiform gyrus. These results suggest that the type of object category determines which region of the fusiform gyrus is activated for processing semantic recognition, whereas the attributes of an object determines the activation in either the left or right fusiform gyrus depending on whether global form or local detail is processed.[22]
In addition, it has been proposed that activation in anterior regions of the fusiform gyri indicate successful recognition.[23] However, levels of activation have been found to depend on the semantic relevance of the object. The term semantic relevance here refers to “a measure of the contribution of semantic features to the ‘‘core’’ meaning of a concept.”[24] Results showed that objects with high semantic relevance, such as artefacts, created an increase in activation compared to objects with low semantic relevance, such as natural objects.[24] This is due to the proposed increased difficulty to distinguish between natural objects as they have very similar structural properties which makes them harder to identify in comparison to artefacts.[23] Therefore, the easier the object is to identify, the more likely it will be successfully recognized.
Another condition that affects successful object recognition performance is that of contextual facilitation. It is thought that during tasks of object recognition, an object is accompanied by a “context frame”, which offers semantic information about the object’s typical context.[25] It has been found that when an object is out of context, object recognition performance is hindered with slower response times and greater inaccuracies in comparison to recognition tasks when an object was in an appropriate context.[25] Based on results from a study using fMRI, it has been proposed that there is a “context network” in the brain for contextually associated objects with activity largely found in the Parahippocampal cortex (PHC) and the Retrosplenial Complex (RSC).[26] Within the PHC, activity in the Parahippocampal Place Area (PPA), has been found to be preferential to scenes rather than objects; however, it has been suggested that activity in the PHC for solitary objects in tasks of contextual facilitation may be due to subsequent thought of the spatial scene in which the object is contextually represented. Further experimenting found that activation was found for both non-spatial and spatial contexts in the PHC, although activation from non-spatial contexts was limited to the anterior PHC and the posterior PHC for spatial contexts.[26]
Recognition Memory
When you see an object, you know what the object is because you've seen it on a past occasion; this is recognition memory. Not only do abnormalities to the ventral (what) stream of the visual pathway effect our ability to recognize an object but also the way in which an object is presented to us.
Familiarity
A mechanism that is context free in the sense that what we recognize just feels familiar rather than spending time trying to find in what context we know this object.[27] The ventro-lateral region of the frontal lobe is involved in memory encoding during incidental learning and then later maintaining and retrieving semantic memories.[27] Familiarity can induce perceptual processes different to those of unfamiliar objects which means that our perception of a finite amount of familiar objects is unique.[28] Deviations from typical viewpoints and contexts can affect the efficiency for which an object is recognized most effectively.[29] It was found that not only are familiar objects recognized more effciently when viewed from a familiar viewpoint opposed to an unfamiliar one, but also this principle applies to novel objects. This deduces to the thought that representations of objects in our brain are organized in more of a familiar fashion of the objects observed in the environment.[29] Recognition is not only largely driven by object shape and/or views but also by dynamic information.[30] Familiarity can benefit the perception of dynamic point-light displays, moving objects, the sex of faces, and face recognition.[29]
Recollection
Recollection shares many similarities with familiarity; however it is context dependent, requiring specific information from the inquired incident.[27]
Impairments
Affects of lesions in the ventral stream
Object recognition is a complex task and involves several different areas of the brain - not just one. If one area is damaged then object recognition can be impaired. The main area for object recognition takes place in the temporal lobe. For example it was found that lesions to the perirhinal cortex in rats causes impairments in object recognition especially with an increase in feature ambiguity.[31] Neonatal aspiration lesions of the amygdaloid complex in monkeys appear to have resulted in a greater object memory loss than early hippocampal lesions. However, in adult monkeys, the object memory impairment is better accounted for by damage to the perirhinal and entorhinal cortex than by damage to the amygdaloid nuclei [32] Combined amygdalohippocampal (A + H) lesions in rats impaired performance on an object recognition task when the retention intervals were increased beyond 0s and when test stimuli were repeated within a session. Damage to the amygdala or hippocampus does not affect object recognition, whereas A + H damage produces clear deficits.[33] In an object recognition task, the level of discrimination was significantly lower in the electrolytic lesions of globus pallidus (part of the basal ganglia) in rats compared to the Substantia- Innominata/Ventral Pallidum which was in turn worse compared to Control and Medial Septum/Vertical Diagonal Band of Broca groups; however, only globus pallidus did not discriminate between new and familiar objects.[34] These lesions damage the ventral (what) pathway of the visual processing of objects in the brain.
Visual Agnosias
Agnosia is a rare occurrence and can be the result of a stroke, dementia, head injury, brain infection, or hereditary.[35] Apperceptive agnosia is a deficit in object perception creating an inability to understand the significance of objects.[27] Similarly Associative agnosia is the inability to understand the significance of objects; however, this time the deficit is in semantic memory.[27] Both of these agnosias can effect the pathway to object recognition, like Marr's Theory of Vision. More specifically unlike apperceptive agnosia, associative agnosic patients are more successful at drawing, copying, and matching taks; however, these patients demonstrate that they can perceive but not recognize.[35] Integrative agnosia(a subtype of associative agnosia)is the inability to integrate separate parts to form a whole image.[27] With these types of agnosias there is damage to the ventral (what) stream of the visual processing pathway. Object orientation agnosia is the inability to extract the orientation of an object despite adequate object recognition.[27] With this type of agnosia there is damage to the dorsal (where) stream of the visual processing pathway. This can effect object recognition in terms of familiarity and even more so in unfamiliar objects and viewpoints. A difficulty in recognizing faces can be explained by prosopagnosia. Someone with prosopagnosia cannot identify the face but is still able to perceive age, gender, and emotional expression.[35] The brain region that specifies in facial recognition is the fusiform face area. Prosopagnosia can also be divided into apperceptive and associative subtypes. Recognition of individual chairs, cars, animals can also be impaired; therefore, these object share similar perceptual features with the face that are recognized in the fusiform face area.[35]
Alzheimer's Disease
The distinction between category and attribute in semantic representation may inform our ability to assess semantic function in aging and disease states affecting semantic memory, such as Alzheimer’s disease (AD).[36] Because of semantic memory deficits, persons suffering from Alzheimer's disease have difficulties recognizing objects as the semantic memory is known to be used to retrieve information for naming and categorizing objects.[37] In fact, it is highly debated whether the semantic memory deficit in AD reflects the loss of semantic knowledge for particular categories and concepts or the loss of knowledge of perceptual features and attributes.[36]
See also
- Visual system
- Perception
- Perceptual constancy
- Visual perception
- Neural processing for individual categories of objects
- Face perception
- Haptic perception
References
- ^ Enns, J. T. (2004). The Thinking Eye, The Seeing Brain: Explorations in Visual Cognition. New York: W. W. Norton & Company.
- ^ Humphreys, G., Price, C., & Riddoch, J. (1999). From objects to names: A cognitive neuroscience approach. Psychological Research , 62, 118-130.
- ^ Riddoch, M., & Humphreys, G. (2001). Object Recognition. In B. Rapp (Ed.), Handbook of Cognitive Neuropsychology. Hove: Psychology Press.
- ^ Ward, J. (2006). The Student's Guide to Cognitive Neuroscience. New York: Psychology Press.
- ^ a b Bar, M. (2003). A cortical mechanism for triggering top-down facilitation in visual object recognition. Journal of Cognitive Neuroscience , 15 (4), 600-609.
- ^ Marr, D. (1976). Early processing of vision information. Philosophical Transactions of the Royal Society of London B , 275, 483-524.
- ^ Richer, F., & Boulet, C. (1999). Frontal lesions and fluctuations in response preparation. Brain and Cognition , 40, 234-238.
- ^ Humphreys, G., & Quinlan, P. (1987). Normal and Pathological Processes in Visual Object Constancy . In G. Humphreys, & M. Riddoch (Eds.), Visual Object Processing: A Cognitive Neuropsychological Approach (pp. 43-105). London: Lawrence Erlbaum Associates .
- ^ a b c Peterson, M. A., & Rhodes, G. (Eds.). (2003). Perception of Faces, Objects and Scenes: Analytic and Holistic Processes. New York: Oxford University Press.
- ^ Marr, D., & Nishihara, H. (1978). Representation and recognition of the spatial organization of three-dimensional shapes. Proceedings of the Royal Society of London , 200, 269-294.
- ^ Biederman, I. (1987). Recognition by components: A theory of human image understanding. Psychological Review , 94, 115-147.
- ^ a b Tarr, M., & Bulthoff, H. (1995). Is human object recognition better described by geon structural descriptions or by multiple views? Comment on Biederman and Gerhardstein (1993). Journal of Experimental Psychology: Human Perception & Performance , 21, 1494-1505.
- ^ Ungerleider, L.G., Mishkin, M., 1982. Two cortical visual systems.In: Ingle, D.J., Goodale, M.A., Mansfield, R.J.W. (Eds.), Analysis of Visual Behavior. InMIT Press, Cambridge, pp. 549–586.
- ^ Goodale, M., & Milner, A. (1992). Separate visual pathways for perception and action. Trends in Neurosciences , 15 (1), 20-25.
- ^ Spiridon, M., Fischl, B., & Kanwisher, N. (2006). Location and spatial profile of category-specific regions in human extrastriate cortex. Human Brain Mapping , 27, 77-89.
- ^ Kourtzi, Z., & Kanwisher, N. (2001). Representation of perceived object shape by the human lateral occipital complex. Science , 293, 1506-1509.
- ^ Grill-Spector, K., Kushnir, T., Edelman, S., Itzchak, Y., & Malach, R. (1998). Cue-invariant activation in object-related areas of the human occipital lobe. Neuron , 21, 191-202.
- ^ Malach, R., Reppas, J., Benson, R., Kwong, K., Jiang, H., Kennedy, W., et al. (1995). Object-related activity revealed by functional magnetic resonance imaging in human occipital cortex. Proceedings of the Natural Academy of Science USA , 92, 8135-8139.
- ^ Grill-Spector, K., Kourtzi, Z., & Kanwisher, N. (2001). The lateral occipital complex and its role in object recognition. Vision Research , 42, 1409-1422.
- ^ Ungerleider, L.G., Mishkin, M., 1982. Two cortical visual systems. In: Ingle, D.J., Goodale, M.A., Mansfield, R.J.W. (Eds.), Analysis of Visual Behavior. InMIT Press, Cambridge, pp. 549–586.
- ^ Kellenbach, M., Hovius, M., & Patterson, K. (2005). A PET study of visual and semantic knowledge about objects. Cortex , 41, 121-132.
- ^ Wierenga, C., Perlstein, W., Benjamin, M., Leonard, C., Rothi, L., Conway, T., et al. (2009). Neural substrates of object identification: Functional magnetic resonance imaging evidence that category and visual attribute contribute to semantic knowledge. Journal of the International Neuropsychological Society , 15, 169-181.
- ^ a b Gerlach, C. (2009). Category-specificity in visual object recognition. Cognition , 111, 281-301.
- ^ a b Mechelli, A., Sartori, G., Orlandi, P., & Price, C. (2006). Semantic relevance explains category effects in medial fusiform gyri. NeuroImage , 30, 992-1002.
- ^ a b Bar, M., & Ullman, S. (1996). Spatial context in recognition. Perception , 25, 343-352.
- ^ a b Bar, M., & Aminoff, E. (2003). Cortical analysis of visual context. Neuron , 38, 347-358.
- ^ a b c d e f g Ward, J. (2006). The Student's Guide to Cognitive Neuroscience. New York: Psychology Press
- ^ Bulthoff, I., & Newell, F. (2006). The role of familiarity in the recognition of static and dynamic objects. Progress in Brain Research , 154, 315-325
- ^ a b c Bulthoff, I., & Newell, F. (2006).The role of familiarity in the recognition of static and dynamic objects. Progress in Brain Research , 154, 315-325
- ^ Vuong, Q., & Tarr, M. (2004). Rotation direction affects object recognition
- ^ Norman, G., & Eacott, M. (2004). Impaired object recognition with increasing levels of feature ambiguity in rats with perirhinal cortex lesions. Behavioural Brain Research , 148, 79-91
- ^ Bachevalier, J., Beauregard, M., & Alvarado, M. C. (1999). Long-term effects of neonatal damage to the hippocampal formation and amygdaloid complex on object discrimination and object recognition in rhesus monkeys. Behavioral Neuroscience, 113.
- ^ Aggleton, J. P., Blindt, H. S., & Rawlins, J. N. P. (1989). Effects of amygdaloid and Amygdaloid–Hippocampal lesions on object recognition and spatial working memory in rats. Behavioral Neuroscience, 103(5), 962-974
- ^ Ennaceur, A. B. D. E. L. K. A. D. E. R. (1998). Effects of lesions of the substantia Innominata/ventral pallidum, globus pallidus and medial septum on rat's performance in object-recognition and radial-maze tasks: Physostigmine and amphetamine treatments. Pharmacological Research, 38(4), 251-263
- ^ a b c d Bauer, R. M. (2006). The agnosias. DC, US: American Psychological Association: Washington
- ^ a b Hajilou, B. B., & Done, D. J. (2007). Evidence for a dissociation of structural and semantic knowledge in dementia of the alzheimer type (DAT). Neuropsychologia, 45(4), 810-816.
- ^ Laatu, S., A, R., Jaykka, H., Portin, R., & Rinne, J. (2003). Visual object recognition in early Alzheimer's disease: deficits in semantic processing. Acta Neurologica Scandinavica , 108, 82-89
Categories:

Wikimedia Foundation. 2010.