- PDE2
The PDE2 (phosphodiesterase 2)
enzyme is one of 21 differentphosphodiesterase s (PDE) found inmammal s. These different PDEs can be subdivided to 11 families (PDE1 – PDE11). The different PDEs of the same family are functionally related despite the fact that theiramino acid sequences show considerable divergence cite news|author=Iffland, A et al|title=Structural determinants for inhibitor specificity and selectivity in PDE2A using the wheat germ in vitro translation system|date=2005|publisher=Biochemistry. 44(23): p. 8312-25.] . The PDEs have different substrate specificities. Some arecAMP ("figure 1") selectivehydrolase s (PDE 4, -7 and -8), others arecGMP ("figure 1") selective hydrolases (PDE 5, -6 and -9) and the rest can hydrolyse bothcAMP andcGMP (PDE1, -2, -3, -10 and -11) cite news|author=Martinez, SE, Wu, AY, Glavas, NA, et al|title=The two GAF domains in phosphodiesterase 2A have distinct roles in dimerization and in cGMP binding|date=2002|publisher=Proc Natl Acad Sci U S A. 99(20): p. 13260-5] .
There is only onegene family coding for the PDE2, which is the PDE2A. Threesplice variant s have been found, the PDE2A1, PDE2A2 and PDE2A3 (PDE2A2 has only been found in rats). PDE2A1 iscytosol ic whereas -A2 and -A3 are membrane bound. It has been suggested that different localization of PDE2A2 and -A3 is due to a unique N-terminal sequence, which is absent in PDE2A1. Despite the PDE2Asplice variant s being different, there is no known differences in their kinetic behavior. (See review article cite news|author=Bender, AT and Beavo, JA|title=Cyclic nucleotide phosphodiesterases: molecular regulation to clinical use|date=2006|publisher=Pharmacol Rev. 58(3): p. 488-520] ).Crystal structure
The
crystal structure of theactive site of the PDE2 enzyme has been reported. (Picture online: http://www.pdb.org/pdb/explore/explore.do?structureId=1Z1L cite news|author=Brookhaven National Laboratory|title=Protein Data Bank|date=2006|publisher=http://www.pdb.org/pdb/explore/explore.do?structureId=1Z1L] ).
Even thoughamino acid sequences, for members of the PDE family show considerable difference (25-35% identity), the overall folding, functional and structural elements of the active sites are very similar. Theactive site is formed by residues that are highly conserved among all PDEs. Thebinding pocket contains metal ion (zinc and magnesium) binding sites. The twohistidine and twoaspartic acid residues, which bind zinc are conserved among all studied PDEs (See review article ).The structure of several other PDE iso-enzymes has been elucidated and among them few co-crystal structures, with inhibitors residing in the active site cite news|author=Card, GL, England, BP, Suzuki, Y, et al|title=Structural basis for the activity of drugs that inhibit phosphodiesterases|date=2004|publisher=Structure. 12(12): p. 2233-47] . The co-crystal structures for PDE4B, PDE4D andPDE5 A have revealed two common features of inhibitor binding to PDEs. One is aplanar ring structure of the inhibitors, which align in theactive site of the enzymes and the other is a conservedglutamine residue (the “glutamine switch” mentioned below), which is essential fornucleotide recognition and selectivity cite news|author=Zhang, KY, Card, GL, Suzuki, Y, et al|title=A glutamine switch mechanism for nucleotide selectivity by phosphodiesterases|date=2004|publisher=Mol Cell. 15(2): p. 279-86] .Substrate selectivity
[

cAMP (to the right) andcGMP to the left. Natural substrates for PDE2."] As mentioned above PDE2 is able to hydrolyze both cAMP and cGMP ("figure 1"), whereas some other members of the PDE family are selective for either of the two cyclic nucleotides. The variability in selectivity towards either cAMP or cGMP is thought to be determined by a so called “glutamine switch”. The “glutamine switch” is an invariant glutamine found in all PDEs, for which thecrystal structure has been solved. In PDE2 this residue is the Gln859. It has potential to formhydrogen bonds with the exocyclicamino group of cAMP and the exocycliccarbonyl oxygen of cGMP. In PDEs, which canhydrolyze bothcAMP andcGMP thisglutamine is able to rotate freely. In PDEs that are selective for either cAMP or cGMP this glutamine is constrained by neighboring residues to a position favoring selectivity for either cyclicnucleotide (See review article ).Regulation
When cGMP binds to the
allosteric GAF-B domain of the PDE, it causesconformational change in theprotein structure leading to higher enzyme activity. Increased hydrolyzis ofcAMP due to binding ofcGMP to the GAF-B domain is well documented, however there are no known examples for the reverse (See review article ). It has been shown that the GAF-B domain has 30-100 fold loweraffinity forcAMP than forcGMP cite news|author=Wu, AY, Tang, XB, Martinez, SE, et al|title=Molecular determinants for cyclic nucleotide binding to the regulatory domains of phosphodiesterase 2A|date=2004|publisher=J Biol Chem. 279(36): p. 37928-38] . This information combined with what is currently known aboutintracellular cAMP concentrations, renders it unlikely that activation ofcGMP hydrolysis bycAMP can take placein vivo . (See review article .)Clinical value of PDE2
PDE2 is expressed in various tissues, for example:
adrenal medulla , brain, heart,platelet ,macrophage s andendothelial cells . The enzyme is thought to be involved in regulating many differentintracellular processes, such as:
*aldosterone secretion from theadrenal gland
* intracellular concentrations of cAMP and cGMP inplatelets
* cGMP inneurons and effect on long-term memory
* barrier function of endothelial cells under inflammatory conditions(See review article )
Several enzyme functions have been reported for the PDE2. It has been shown that PDE2 lowers cAMP through increased cGMP caused byatrial natriuretic peptide (ANP) resulting in decreasedaldosterone secretion (See review article ).
It has also been suggested that PDE2 might play an important role in the regulation of elevatedintracellular concentrations of cAMP and cGMP in platelets. PDE3 is an important player in platelet aggregation. It has been reported that higher concentration of cGMP causes inhibition ofPDE3 , whereas it stimulates PDE2. Interplay between those two functions seems to mediate an opposing regulation of cAMP in platelets (See review article ).
PDE2 regulates cardiac L-type Ca2+ current incardiac myocyte s, where activation of PDE2 by cGMP lowers cAMP and thereby affecting cardiac function. (See review article ).
PDE2 is expressed in several regions of the brain and rat experiments have indicated that inhibition of PDE2 enhances functions such as memory(See review article ).
PDE2 is up regulated whenmonocyte s differentiate intomacrophages , but the role of PDE2 in matured macrophages is yet to be characterized. Furthermore, PDE2 has been indicated to play a role in inflammatory responses as it has been detected in microvessels, but not in larger vessels. It has been speculated thattumor necrosis factor-alpha (TNFα) might regulate the function of PDE2 inendothelial cells and thereby affecting flow of fluid and cells through the endothelial barrier asin vitro experiments on endothelial cells show up regulation of both PDE2mRNA and activity. (See review article )
Until now PDE2 inhibitors have mostly been used as research tools, but are presently being investigated for improving memory and decreasing endothelial permeability under inflammatory conditions (See review article ).PDE2A inhibitors
[

EHNA . An PDE2 inhibitor."]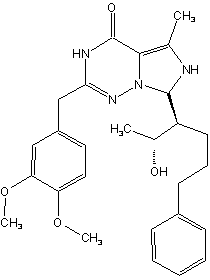 thumb|200px|inhibitor.
thumb|200px|inhibitor.EHNA
The first specific inhibitor developed for PDE2 was
EHNA (erythro-9-(2-hydroxy-3-nonyl)adenine, "figure 2"). It has been demonstrated to specifically act on PDE2 by inhibiting cGMP-activation of PDE2 with an IC50 value of ~1µM and an at least 50-fold selectivity over other PDEs cite news|author=Podzuweit, T, Nennstiel, P, and Muller, A,|title=Isozyme selective inhibition of cGMP-stimulated cyclic nucleotide phosphodiesterases by erythro-9-(2-hydroxy-3-nonyl) adenine|date=1995|publisher=Cell Signal. 7(7): p. 733-8] . The core structure of EHNA resembles cAMP but differentiates in the fact that EHNA has a bulkyhydrophobic carbon side chain replacing the phospho-ribosemoiety in cAMP .Inhibitory effects of EHNA
In
primary culture s of rat cortical neurons, the inhibition of PDE2A by EHNA potentiatesNMDA (N-metyl-D-aspartate) receptor activated increase in cGMP, but has no effect on cAMP concentrations. cite news|author=Suvarna, NU and O'Donnell, JM|title=Hydrolysis of N-methyl-D-aspartate receptor-stimulated cAMP and cGMP by PDE4 and PDE2 phosphodiesterases in primary neuronal cultures of rat cerebral cortex and hippocampus|date=2002|publisher=J Pharmacol Exp Ther, 2002. 302(1): p. 249-56] EHNA is also a verypotent adenosine deaminase inhibitor with an IC50 ~2 nM cite news|author=Boess, FG, Hendrix, M, van der Staay, FJ, et al|title=Inhibition of phosphodiesterase 2 increases neuronal cGMP, synaptic plasticity and memory performance|date=2004|publisher=Neuropharmacology. 47(7): p. 1081-92] . This dual inhibition would lead to the accumulation of the two inhibitorymetabolite s, adenosine cite news|author=van Calker, D, Muller, M, and Hamprecht, B|title=Adenosine inhibits the accumulation of cyclic AMP in cultured brain cells|date=1978|publisher=Nature. 276(5690): p. 839-41] and cGMP cite news|author=Goldberg, ND, Haddox, MK, Nicol, SE, et al|title=Biologic regulation through opposing influences of cyclic GMP and cyclic AMP: the Yin Yang hypothesis|date=1975|publisher=Adv Cyclic Nucleotide Res. 5: p. 307-30] , which may act in synergy to mediate diverse pharmacological responses including anti-viral, anti-tumour and anti-arrhythmic effects. Although EHNA potently inhibitsadenosine deaminase , it has been successfully used with the proper controls as a tool to probe PDE2 functions. EHNA has been used to study implication of PDE2 in calcium control in cardiac myocytes cite news|author=Rivet-Bastide, M, Vandecasteele, G, Hatem, S, et al|title=cGMP-stimulated cyclic nucleotide phosphodiesterase regulates the basal calcium current in human atrial myocytes|date=1997|publisher=J Clin Invest. 99(11): p. 2710-8] and has shown to be effective to reverse hypoxic pulmonary vasoconstricion in perfused lung models cite news|author=Haynes, J, Jr., Killilea, DW, Peterson, PD, et al|title=1996|date=1996|publisher=J Pharmacol Exp Ther. 276(2): p. 752-7] .EHNA has been therefore been used for two purposes:
# to serve as alead structure for the rational design of more selective and potent PDE2 inhibitors, and
# to define some of PDE’s biological targets.
However, the use of EHNA as a chemical tool in determing the pharmacological role of PDE2 is limited due to low PDE2 potency and high potency in inhibiton of adenosine deaminase cite news|author=Chambers, RJ, Abrams, K, Garceau, NY, et al|title=A new chemical tool for exploring the physiological function of the PDE2 isozyme|date=2006|publisher=Bioorg Med Chem Lett. 16(2): p. 307-10] .BAY 60-7550, Oxindole and PDP
BAY 60-7550 ("figure 3") is an analog of
EHNA , which is more than 100-fold more potent and is highly selective for PDE2A . Other newly discovered selecitve PDE2 inhibitors are PDP (9-(6-Phenyl-2-oxohex-3-yl)-2-(3,4-dimethoxybenzyl)-purin-6-one, "figure 4") cite news|author=Seybold, J, Thomas, D, Witzenrath, M, et al|title=Tumor necrosis factor-alpha-dependent expression of phosphodiesterase 2: role in endothelial hyperpermeability|date=2005|publisher=Blood. 105(9): p. 3569-76] and Oxindole ("figure 5") ."Table 1" shows the
potency of PDE2 inhibitors including EHNA. There is a large increase in potency between EHNA, Bay 60-7550 and PDP. The large dimethoxybenzyl group in position 2 of the purine moiety of Bay-60 7550 and PDP might be contributing to the added potency.Structure and bonding of the inhibitors
Comparison of these inhibitors with the natural substrates of the enzyme, cAMP and cGMP ("see figure 1 through 5") reveal some common characteristics of the molecules. The main characteristic of all the molecules is the flat moiety comprising at least two fused ring structures, a six atom ring and a five atom ring. This ring system in cGMP and cAMP is a purine ring system, and the same is true for EHNA and PDP. Bay 60-7550 and oxindole lack the purine core but do possess a related ring system. Hydrogen bond acceptors, mostly nitrogen but also oxygen, reside in the ring system of the inhibitors. These atoms might interact with hydrogen bond donators, which are part of amino acids in the active site of the enzyme and thereby contribute to the inhibition of the enzyme from hydrolyzing cAMP and cGMP similar to how the natural substrates bind to the active site .
Structural similarity of the inhibitors
The structures of Bay 60-7550 and PDP are very similar ("see figure 3 and 4"). The difference between these molecules is the exocyclic
methyl group on Bay 60-7550, which replaces the nitrogen atom in PDP decreasing the possibility to formhydrogen bond s with the enzyme in an important site for substrate and inhibitor binding. The oxindole structure differs from the other inhibitors since it is more divergent from thepurine ring system and has less hydrogen binding possibilities. The molecule also lacks the large side group, analogous to the dimethoxybenzyl group of Bay 60-7550 and PDP. It is difficult to predict possible interactions to the enzyme without a co crystal structure of the phenomenon.Possible structure-activity relationship for PDE2 inhibitors
There is a lack of co crystal structures of inhibitors bound in the
active site of PDE2. However, a computerizeddocking model of the inhibitor EHNA and the substrates cAMP and cGMP bound in the catalytic site have been made . The docking model of EHNA showed that themutation s of the amino acids Asp811 to Ala (Asp811Ala) and Ile826 to Val (Ile826Val) at theactive site , where the only amino acid substitutions that significantly affected the inhibition by EHNA. The Asp811 mutation to alanine increased IC50 value for EHNA 6-fold and the Ile826 mutation to valine leads to a 7-fold increased IC50 value for EHNA compared to wild type PDE2A.Upper binding pocket: Gln859 and Asp811
EHNA is in close proximity to Gln859 at the active site, which could donate two hydrogen bonds to N1 and N6 of the nitrogen atoms in the adenine ring of EHNA. On the other site of the binding pocket the Asp811 could donate another hydrogen bond to N7 in the adenine ring in order to stabilize the bond inhibitor. This hypothesis is supported by the fact that the Asp811Ala mutant has decreased activity toward cAMP, whereas activity toward cGMP is unchanged.
The lower binding pocket: Ile 826
The residues in the lower binding pocket may lie too far away for interaction with the inhibitor and therefore might be irrelevant for EHNA selectivity.However the residues may play an indirect role of EHNA selectivity. Ile826 is positioned below the purine ring of EHNA and thereby limits the space for EHNA. Substitution with smaller valine (Ile826Val mutation) could increase the space for EHNA and cause the loss of hydrogen binding with residues in the upper binding pocket, while improving hydrogen binding within the lower binding pocket. This shift of interactions could destabilize binding of the adenine ring of EHNA, which could be the reason for higher IC50 value.
No models are available for the other inhibitors than EHNA, which align in the active site. Therefore it is more difficult to interpret the molecular binding. When looking at the inhibitors and their overall similarity, it is likely that they bind with a similar mechanism to the active site and that the different side groups determine potency of the inhibitor.The determinants of inhibitor specificity within the PDE2 active site are not very well known and with better understanding of these determinants it would facilitate the development of inhibitors with increased potency .
References


Wikimedia Foundation. 2010.